A Novel Pectic Polysaccharide of Jujube Pomace: Structural Analysis and Intracellular Antioxidant Activities



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Extraction and Purification of SAZMP4
2.3. Structural analysis of SAZMP4
2.3.1. Physical and Chemical Analysis
2.3.2. Methylation Analysis
2.3.3. NMR Analysis
2.4. Molecular Morphological Analysis
2.5. Antioxidant Activity of SAZMP4
2.5.1. Cell Culture
2.5.2. Cell Viability Analysis
2.5.3. Treatment Procedure
2.5.4. Measurement of SOD and GSH-Px
2.5.5. Intracellular ROS and MDA Levels
2.6. Data Analysis
3. Results
3.1. Separation and Purification of SAZMP4
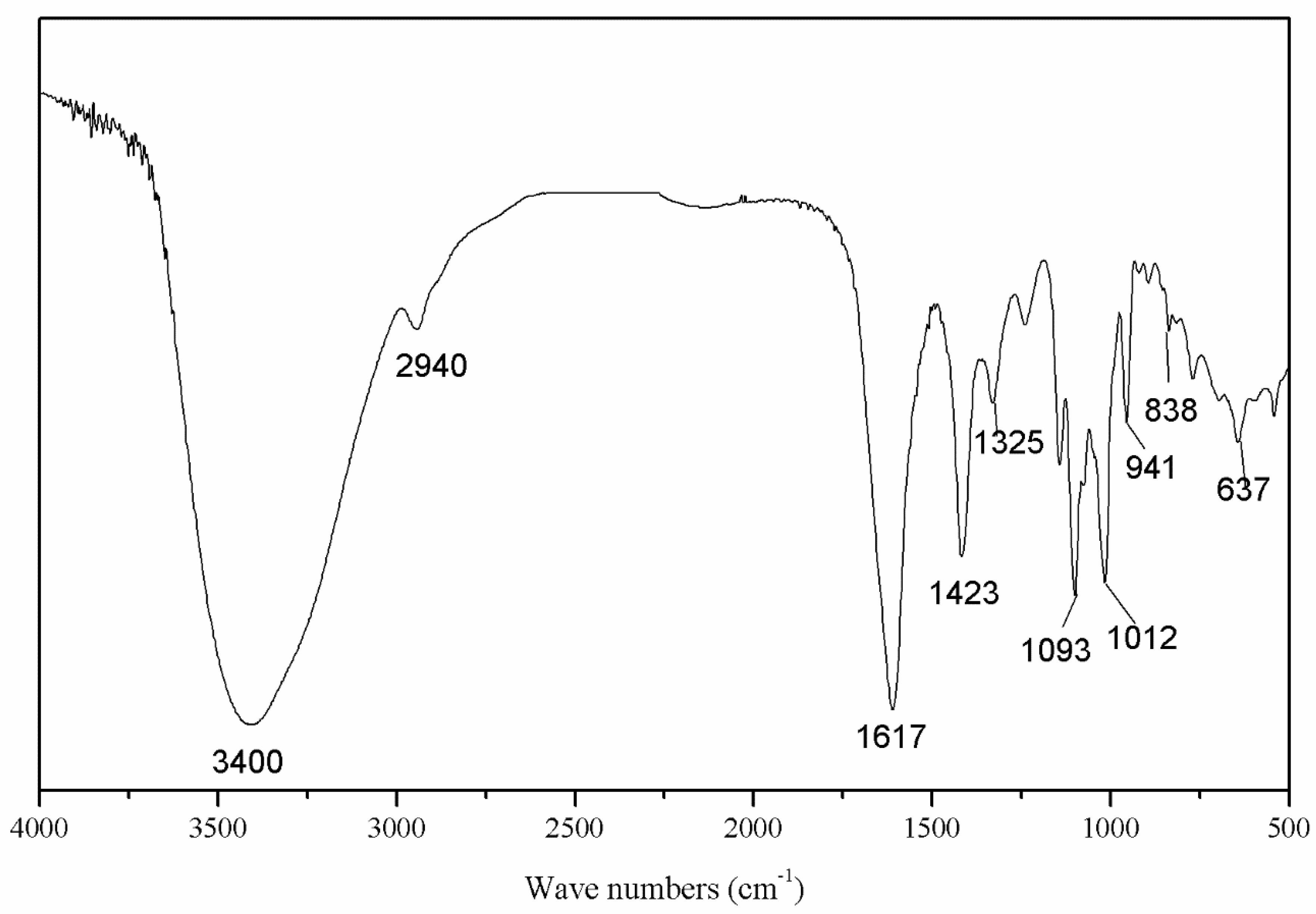
3.2. Preliminary Characterizations of SAZMP4
3.3. Methylation Analysis
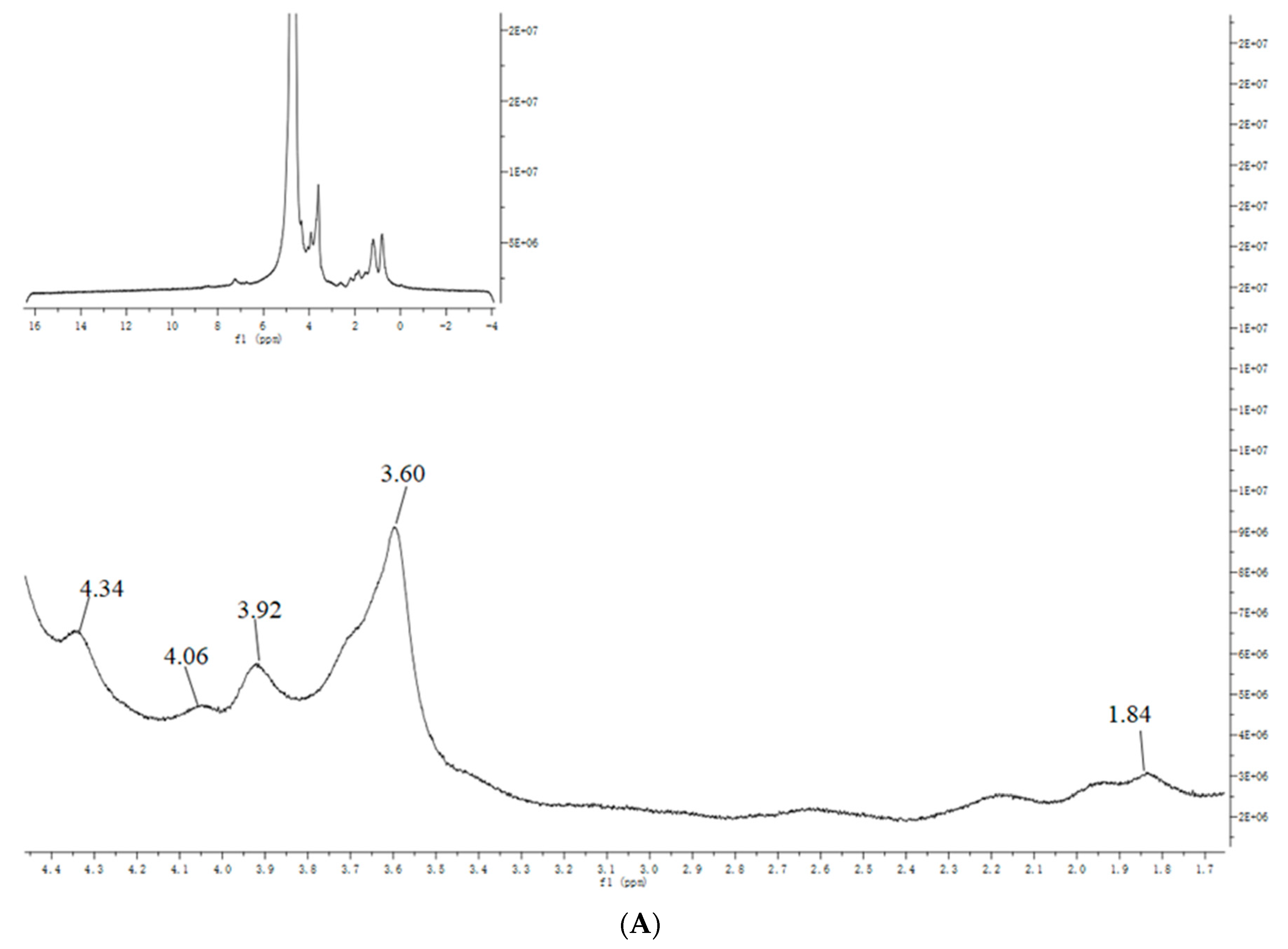
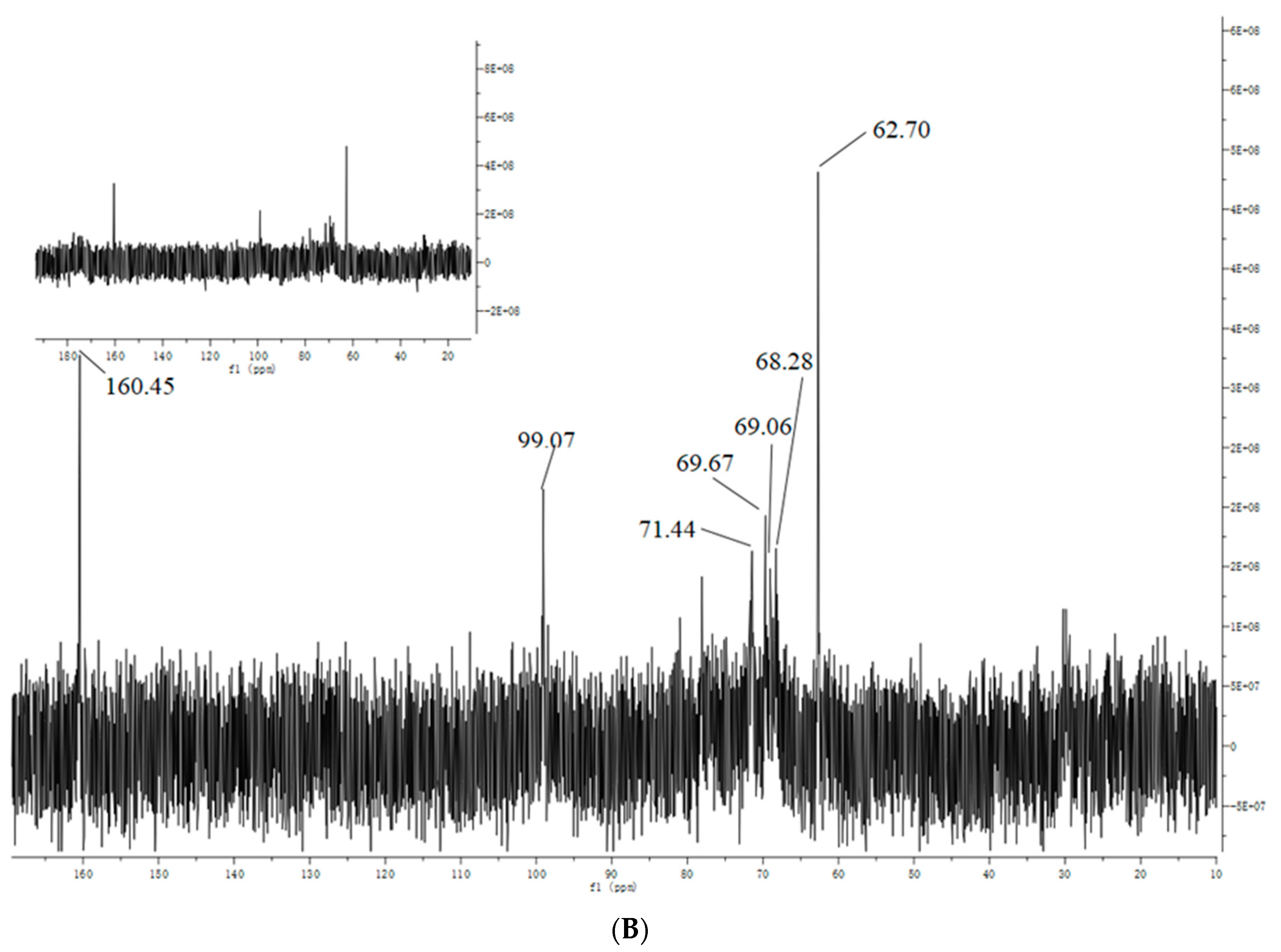
3.4. NMR Analysis
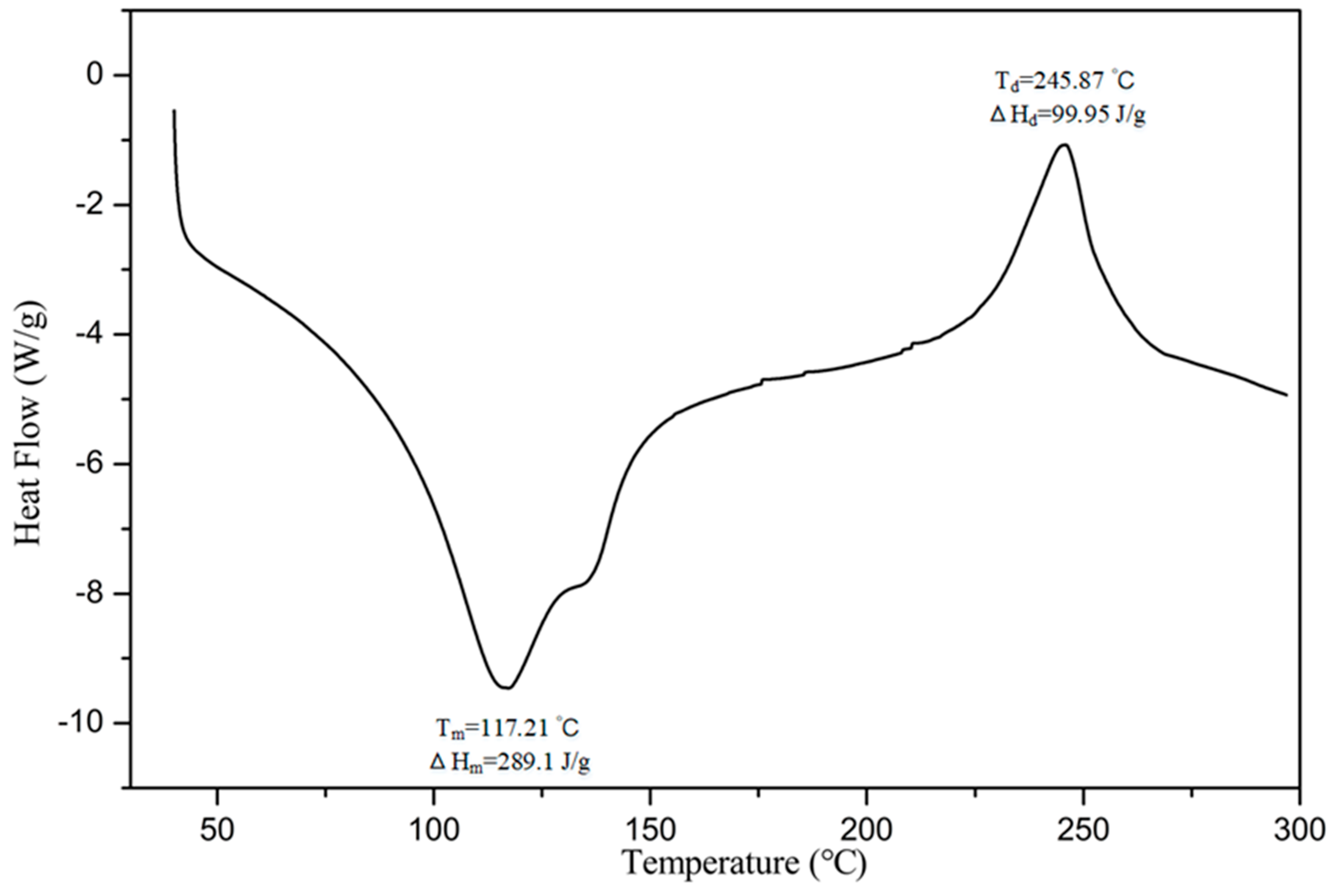
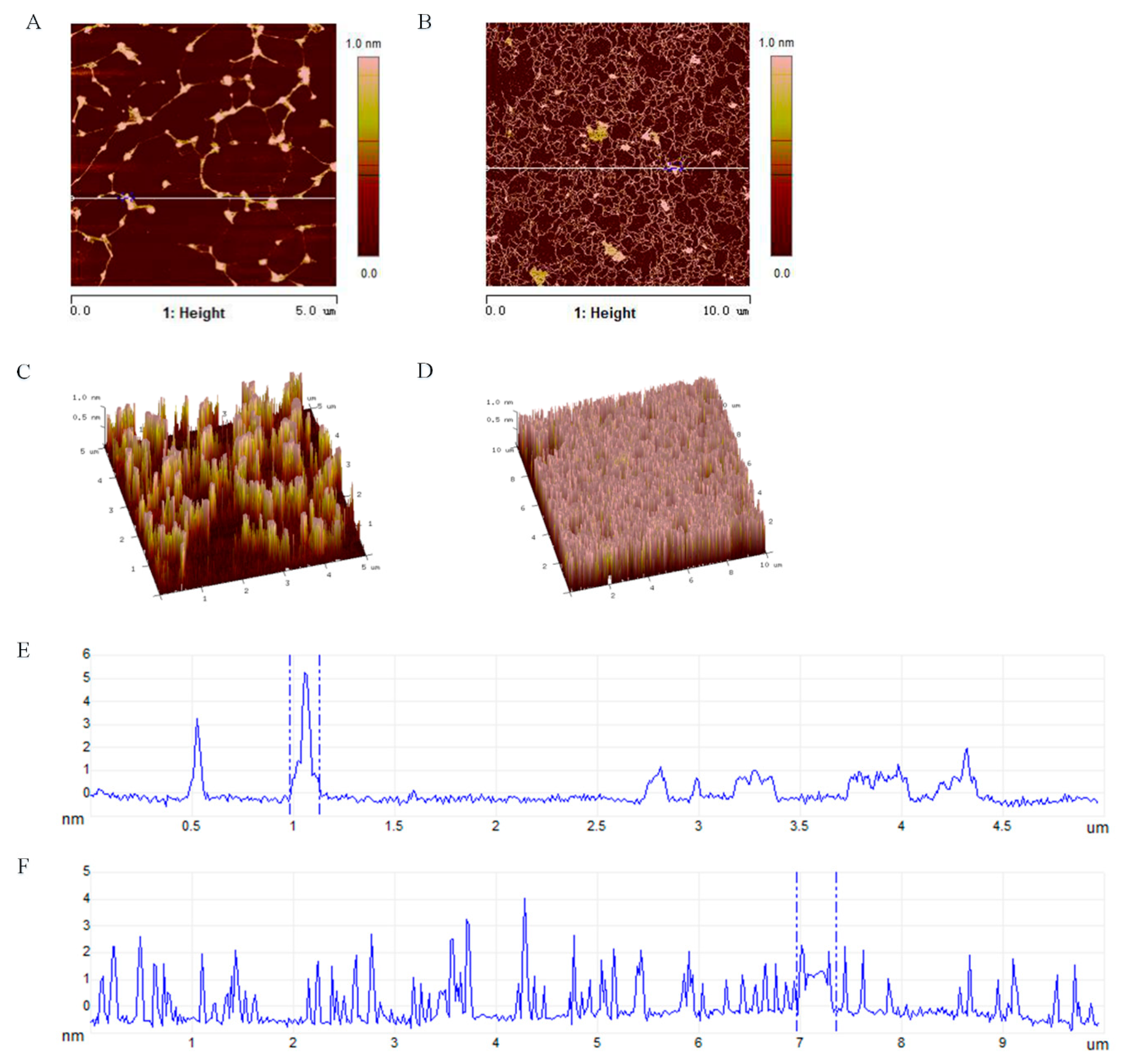
3.5. Molecular Morphological Properties
3.6. Antioxidant Activity of SAZMP4
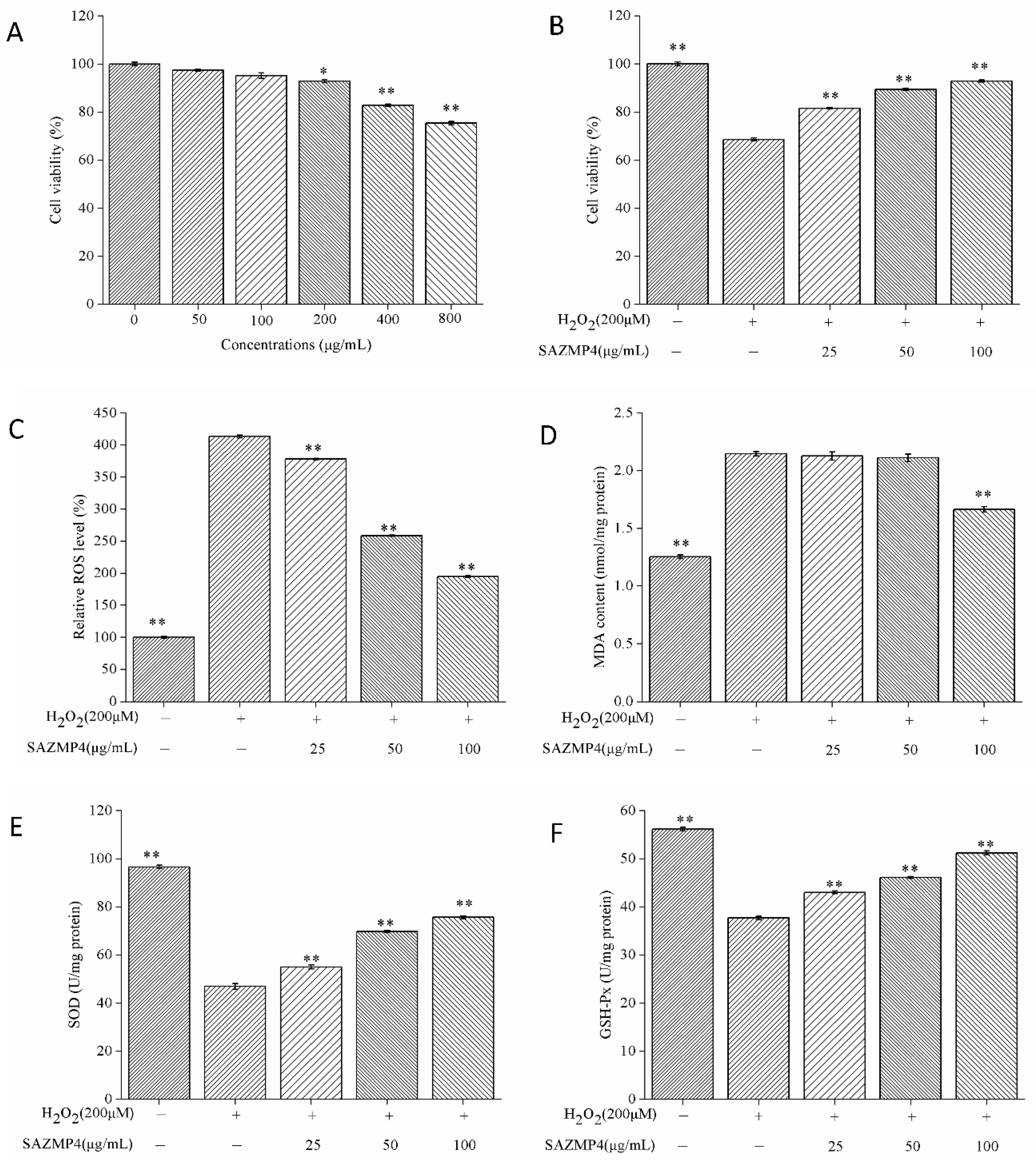
3.6.1. The Cytotoxicity of SAZMP4 to Caco-2 Cells
3.6.2. Protective Effect against H2O2-induced Toxicity
3.6.3. ROS and MDA Levels in Caco-2 Cells
3.6.4. The Activity of SOD and GSH-Px in Caco-2 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Noreen, A.; Nazli, Z.I.H.; Akram, J.; Rasul, I.; Mansha, A.; Yaqoob, N.; Iqbal, R.; Tabasum, S.; Zuber, M.; Zia, K.M. Pectins functionalized biomaterials; a new viable approach for biomedical applications: A review. Int. J. Biol. Macromol. 2017, 101, 254–272. [Google Scholar] [CrossRef] [PubMed]
- Qiu, N.; Tian, Y.; Qiao, S.; Deng, H. Apple Pectin Behavior Separated by Ultrafiltration. Agri. Sci. China 2009, 8, 1193–1202. [Google Scholar] [CrossRef]
- John, I.; Muthukumar, K.; Arunagiri, A. A review on the potential of citrus waste for d-Limonene, pectin, and bioethanol production. Int. J. Green Energy. 2017, 14, 599–612. [Google Scholar] [CrossRef]
- Sun, Y.; Hou, S.; Song, S.; Zhang, B.; Ai, C.; Chen, X.; Liu, N. Impact of acidic, water and alkaline extraction on structural features, antioxidant activities of Laminaria japonica polysaccharides. Int. J. Biol. Macromol. 2018, 112, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Y.; Fan, L.; Ai, L.; Shan, L. Antioxidant activities of polysaccharides from the fruiting bodies of Zizyphus Jujuba cv. Jinsixiaozao. Carbohydr. Polym. 2011, 84, 390–394. [Google Scholar] [CrossRef]
- Chen, J.; Du, C.Y.; Lam, K.Y.; Zhang, W.L.; Lam, C.T.; Yan, A.L.; Yao, P.; Lau, D.T.; Dong, T.T.; Tsim, K.W. The standardized extract of ziziphus jujuba fruit (Jujube) regulates pro-inflammatory cytokine expression in cultured murine macrophages: suppression of lipopolysaccharide-stimulated NF-kB activity. Phytother. Res. 2014, 28, 1527–1532. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Wu, S.; Zhang, H.; Zhang, X.; Niu, Y.; Cao, X.; Huang, F.; Ding, H. Characterization and hepatoprotective effect of polysaccharides from Ziziphus jujuba Mill. var. spinosa (Bunge) Hu ex HF Chou sarcocarp. Food Chem. Toxicol. 2014, 74, 76–84. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, X.; Ren, D.; Wang, D.; Xuan, Y. Preventive effects of jujube polysaccharides on fructose-induced insulin resistance and dyslipidemia in mice. Food Funct. 2014, 5, 1771–1778. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Zhang, J.; Liu, G.; Liu, Y.; Wang, K.; Yang, K.; Cheng, H.; Zhao, Z. Structural characterization and in vitro antitumor activity of polysaccharides from Zizyphus jujuba cv. Muzao. RSC Adv. 2015, 5, 7860–7867. [Google Scholar] [CrossRef]
- Huang, H.; Chen, F.; Long, R.; Huang, G. The antioxidant activities in vivo of bitter gourd polysaccharide. Int. J. of Biol. Macromol. 2020, 145, 141–144. [Google Scholar] [CrossRef]
- Chen, S.; Huang, H.; Huang, G. Extraction, derivatization and antioxidant activity of cucumber polysaccharide. Int. J. of Biol. Macromol. 2019, 140, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Li, L. Structural characterization and antioxidant activity of polysaccharide from four auriculariales. Carbohydr. Polym. 2020, 229, 115407. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhu, P.; Zhao, S.; Nie, C.; Wang, N.; Du, X.; Zhou, Y. Characterization, antioxidant activity and immunomodulatory activity of polysaccharides from the swollen culms of Zizania latifolia. Int. J. of Biol. Macromol. 2017, 95, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. Lawrence Berkeley Natl. Lab. 1955, 11, 298–300. [Google Scholar] [CrossRef] [Green Version]
- Sugamura, K.; Keaney, J.F. Reactive oxygen species in cardiovascular disease. Free. Radic. Biol. Med. 2011, 51, 978–992. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Ji, X.; Wang, M.; Yin, S.; Peng, Q. An alkali-extracted polysaccharide from Zizyphus jujuba cv. Muzao: Structural characterizations and antioxidant activities. Int. J. of Biol. Macromol. 2019, 136, 607–615. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Harmilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. of Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Seedevi, P.; Moovendhan, M.; Sudharsan, S.; Sivasankar, P.; Sivakumar, L.; Vairamani, S.; Shanmugam, A. Isolation and chemical characteristics of rhamnose enriched polysaccharide from Grateloupia lithophila. Carbohydr. Polym. 2018, 195, 486–494. [Google Scholar] [CrossRef]
- Ren, J.; Hou, C.; Shi, C.; Lin, Z.; Liao, W.; Yuan, E. A polysaccharide isolated and purified from Platycladus orientalis (L.) Franco leaves, characterization, bioactivity and its regulation on macrophage polarization. Carbohydr. Polym. 2019, 213, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, X. Characterization of pectic polysaccharides extracted from apple pomace by hot-compressed water. Carbohydr. Polym. 2014, 102, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, L.; Wu, J.; Xu, W.; Wang, X.; Lu, X. Improvement of simultaneous determination of neutral monosaccharides and uronic acids by gas chromatography. Food Chem. 2017, 220, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, J.; Li, H.; Xu, R.; Yuan, Y.; Cao, J. Physicochemical properties and functional bioactivities of different bonding state polysaccharides extracted from tomato fruit. Carbohydr. Polym. 2019, 219, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.L.; Conrad, H.E. Stoichiometric depolymerization of polyuronides and glycosaminoglycuronans to monosaccharides following reduction of their carbodiimide-activated carboxyl groups. Biochem. 1972, 11, 1383–1388. [Google Scholar] [CrossRef]
- Needs, P.W.; Selvendran, R.R. Avoiding oxidative-degradation during dodium-hydroxide methyl iodlde-mediated carbohydrate methylation in dimethyl-sulfoxide. Carbohydr. Polym. 1993, 245, 1–10. [Google Scholar] [CrossRef]
- Chaves, P.F.P.; Iacomini, M.; Cordeiro, L.M.C. Chemical characterization of fructooligosaccharides, inulin and structurally diverse polysaccharides from chamomile tea. Carbohydr. Polym. 2019, 214, 269–275. [Google Scholar] [CrossRef]
- Yan, J.K.; Ding, Z.C.; Gao, X.; Wang, Y.Y.; Yang, Y.; Wu, D.; Zhang, H.N. Comparative study of physicochemical properties and bioactivity of Hericium erinaceus polysaccharides at different solvent extractions. Carbohydr. Polym. 2018, 193, 373–382. [Google Scholar] [CrossRef]
- Wang, L.; Liu, H.M.; Qin, G.Y. Structure characterization and antioxidant activity of polysaccharides from Chinese quince seed meal. Food Chem. 2017, 234, 314–322. [Google Scholar] [CrossRef]
- Zou, M.; Chen, Y.; Sun-Waterhouse, D.; Zhang, Y.; Li, F. Immunomodulatory acidic polysaccharides from Zizyphus jujuba cv. Huizao: Insights into their chemical characteristics and modes of action. Food Chem. 2018, 258, 35–42. [Google Scholar] [CrossRef]
- Huang, F.; Liu, H.; Zhang, R.; Dong, L.; Liu, L.; Ma, Y.; Jia, X.; Wang, G.; Zhang, M. Physicochemical properties and prebiotic activities of polysaccharides from longan pulp based on different extraction techniques. Carbohydr. Polym. 2019, 206, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, Y.; Duan, X.; Tang, T.; Shen, Y.; Hu, B.; Liu, A.; Chen, H.; Li, C.; Liu, Y. Characterization and antioxidant activities of polysaccharides from thirteen boletus mushrooms. Int. J. of Biol. Macromol. 2018, 113, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, X.; Wang, Y.; Liu, G.; Zhang, Z.; Zhao, Z.; Cheng, H. In vitro antioxidative and immunological activities of polysaccharides from Zizyphus Jujuba cv. Muzao. Int. J. of Biol. Macromol. 2017, 95, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Yan, Y.; Hou, C.; Shi, M.; Liu, Y. Structural characterization of a galacturonic acid-rich polysaccharide from Ziziphus Jujuba cv. Muzao. Int. J. of Biol. Macromol. 2019. [Google Scholar] [CrossRef]
- Iijima, M.; Nakamura, K.; Hatakeyama, T.; Hatakeyama, H. Phase transition of pectin with sorbed water. Carbohydr. Polym. 2000, 41, 101–106. [Google Scholar] [CrossRef]
- Godeck, R.; Kunzek, H.; Kabbert, R. Thermal analysis of plant cell wall materials depending on the chemical structure and pre-treatment prior to drying. Eur. Food Res. Technol. 2001, 213, 395–404. [Google Scholar] [CrossRef]
- Bjorndal, H.; Hellerqv, C.; Lindberg, B.; Svensson, S. Gas-liquid chromatography and mass spectrometry in methylation analysis of polysaccharides. Angew. Chem. -Int. Ed. 1970, 9, 610–618. [Google Scholar] [CrossRef]
- Ridley, B.L.; O’Neill, M.A.; Mohnen, D. Pectins: structure, biosynthesis, and oligogalacturonide-related signaling. Phytochem. 2001, 57, 927–967. [Google Scholar] [CrossRef]
- Agrawal, P.K. NMR spectroscopy in the structural elucidation of oligosaccharides and glycoside. Phytochem. 1992, 31, 3307–3330. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, H.; Zhang, N.; Chen, S.; Tian, J.; Zhang, Y.; Wang, Z. Extrusion treatment for improved physicochemical and antioxidant properties of high-molecular weight polysaccharides isolated from coarse tea. Food Res. Int. 2013, 53, 726–731. [Google Scholar] [CrossRef]
- Myrick, J.M.; Vendra, V.K.; Krishnan, S. Self-assembled polysaccharide nanostructures for controlled-release applications. Nanotechnol. Rev. 2014, 3, 319–346. [Google Scholar] [CrossRef]
- Ochiai, A.; Itoh, T.; Kawamata, A.; Hashimoto, W.; Murata, K. Plant cell wall degradation by saprophytic Bacillus subtilis strains: gene clusters responsible for rhamnogalacturonan depolymerization. Appl. Environ. Microbiol. 2007, 73, 3803–3813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, P.; Zhang, A.; Zhang, F.; Linhardt, R.J.; Sun, P. Structure and bioactivity of a polysaccharide containing uronic acid from Polyporus umbellatus sclerotia. Carbohydr. Polym. 2016, 152, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, J.; Wu, X.; Dai, H.; Gao, X.; Liu, M.; Tu, P. Structures and immunological activities of two pectic polysaccharides from the fruits of Ziziphus jujuba Mill. cv. jinsixiaozao Hort. Food Res. Int. 2006, 39, 917–923. [Google Scholar] [CrossRef]
- Misaki, M.K.A.; Sasaki, T.; Tanaka, M.; Miyaji, H. Studies on interrelation of structure and antitumor effects of polysaccharides: Antitumor action of periodate-modified, branched (1 → 3)- β-d-glucan of auricularia auricula-judae, and other polysaccharides containing (1 → 3)-glycosidic linkages. Carbohydr. Res. 1981, 92, 115–129. [Google Scholar] [CrossRef]
- Ahn, S.; Halake, K.; Lee, J. Antioxidant and ion-induced gelation functions of pectins enabled by polyphenol conjugation. Int. J. of Biol. Macromol. 2017, 101, 776–782. [Google Scholar] [CrossRef]
- Klosterhoff, R.R.; Bark, J.M.; Glanzel, N.M.; Iacomini, M.; Martinez, G.R.; Winnischofer, S.M.B.; Cordeiro, L.M.C. Structure and intracellular antioxidant activity of pectic polysaccharide from acerola (Malpighia emarginata). Int. J. of Biol. Macromol. 2018, 106, 473–480. [Google Scholar] [CrossRef]
- Yu, X.; Kensler, T. Nrf2 as a target for cancer chemoprevention. Mutat. Res. 2005, 591, 93–102. [Google Scholar] [CrossRef]
- Ramyaa, P.; Krishnnaswamy, R.; Padma, V.V. Quercetin modulates OTA-induced oxidative stress and redox signaling in HepG2 cells-up regulation of Nrf2 expression and down regulation of NF- NF-κB and COX-2. Biochim. et Biophys. Acta-Gen. Subj. 2014, 1840, 681–692. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Retention Time (min) | Monosaccharide | Molar Radio |
|---|---|---|---|
| 1 | 11.645 | Rhamnose | 1 |
| 2 | 12.289 | Arabinose | 0.9 |
| 3 | 12.921 | Xylose | 0.05 |
| 4 | 22.800 | Mannose | 0.07 |
| 5 | 38.020 | Galacturonic acid | 28.9 |
| Peak No. | Retention Time (min) | Methylated Sugars | Linkage Patterns | Molar Radio |
|---|---|---|---|---|
| 1 | 20.355 | 2,3,4-Me3-Rhap | 1-linked Rhap | 0.5 |
| 2 | 26.563 | 2,3,5-Me3-Araf | 1-linked Araf | 0.47 |
| 3 | 31.915 | 2-Me-Araf | 1,3,5-linked Araf | 0.46 |
| 4 | 34.707 | 3-Me-Rhap | 1,2,4-linked Rhap | 0.52 |
| 5 | 35.092 | 2,3,6-Me3-Galp | 1,4-linked Galp | 28.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, X.; Liu, K.; Yin, S.; Qin, Y.; Shen, P.; Peng, Q. A Novel Pectic Polysaccharide of Jujube Pomace: Structural Analysis and Intracellular Antioxidant Activities. Antioxidants 2020, 9, 127. https://doi.org/10.3390/antiox9020127
Lin X, Liu K, Yin S, Qin Y, Shen P, Peng Q. A Novel Pectic Polysaccharide of Jujube Pomace: Structural Analysis and Intracellular Antioxidant Activities. Antioxidants. 2020; 9(2):127. https://doi.org/10.3390/antiox9020127
Chicago/Turabian StyleLin, Ximeng, Keshan Liu, Sheng Yin, Yimin Qin, Peili Shen, and Qiang Peng. 2020. "A Novel Pectic Polysaccharide of Jujube Pomace: Structural Analysis and Intracellular Antioxidant Activities" Antioxidants 9, no. 2: 127. https://doi.org/10.3390/antiox9020127
APA StyleLin, X., Liu, K., Yin, S., Qin, Y., Shen, P., & Peng, Q. (2020). A Novel Pectic Polysaccharide of Jujube Pomace: Structural Analysis and Intracellular Antioxidant Activities. Antioxidants, 9(2), 127. https://doi.org/10.3390/antiox9020127