Phenolic and Anthocyanin Compounds and Antioxidant Activity of Tamarillo (Solanum betaceum Cav.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Chemicals, Reagents and Standards
2.3. Extraction and Quantification of Phenolics
2.4. Extraction and Quantification of Anthocyanins
2.5. Total Phenolic Content (TPC)
2.6. Antioxidant Activity by Ferric Reducing Ability of Plasma (FRAP) and Cupric ion-Reducing Antioxidant Capacity (CUPRAC) Assays
2.7. Statistical Analysis
3. Results
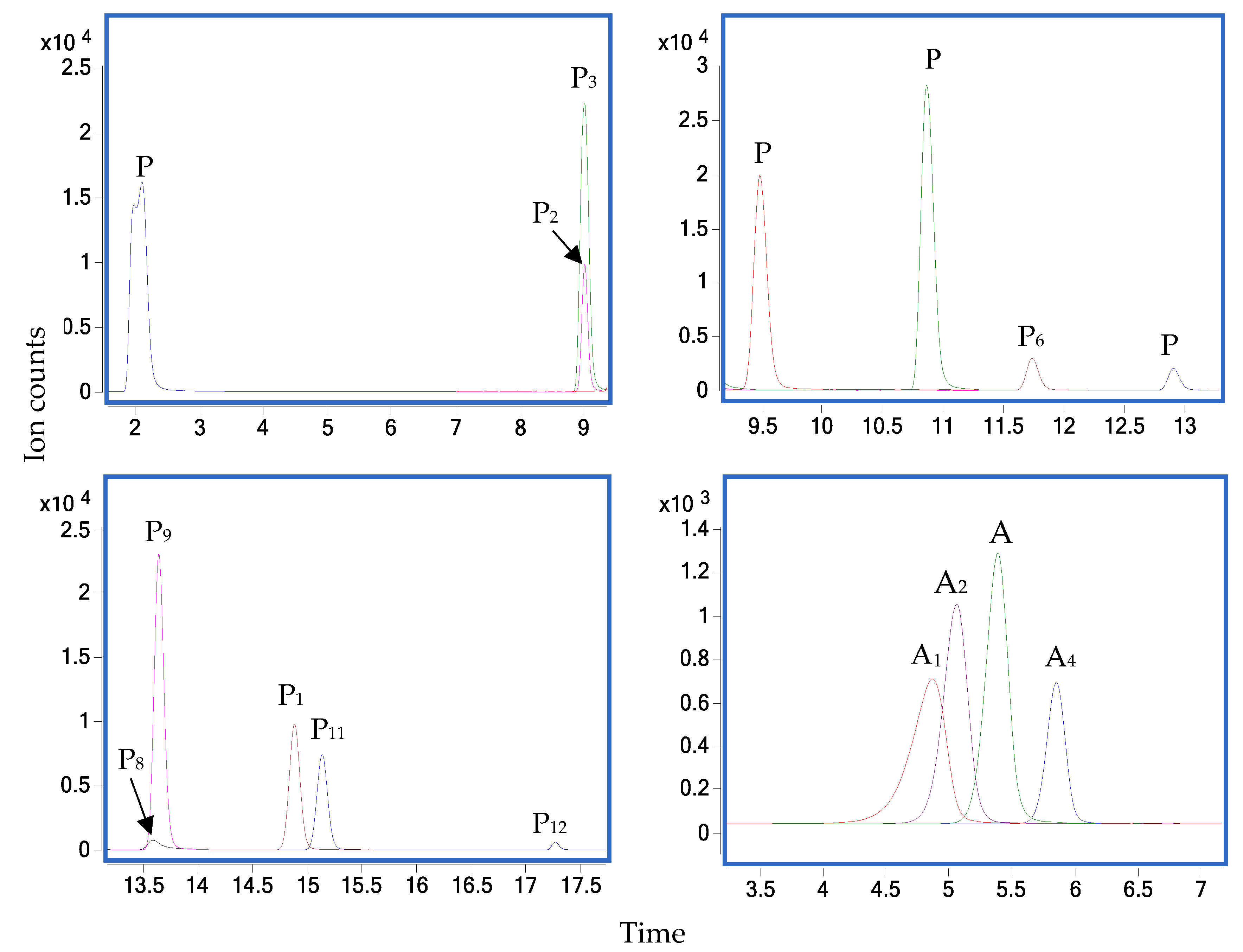
3.1. Phenolic Compound Profiles
3.2. Anthocyanin Compound Profiles
3.3. Total Phenolic Content (TPC)
3.4. Antioxidant Activity
3.5. Correlation between TPC, CUPRAC and FRAP
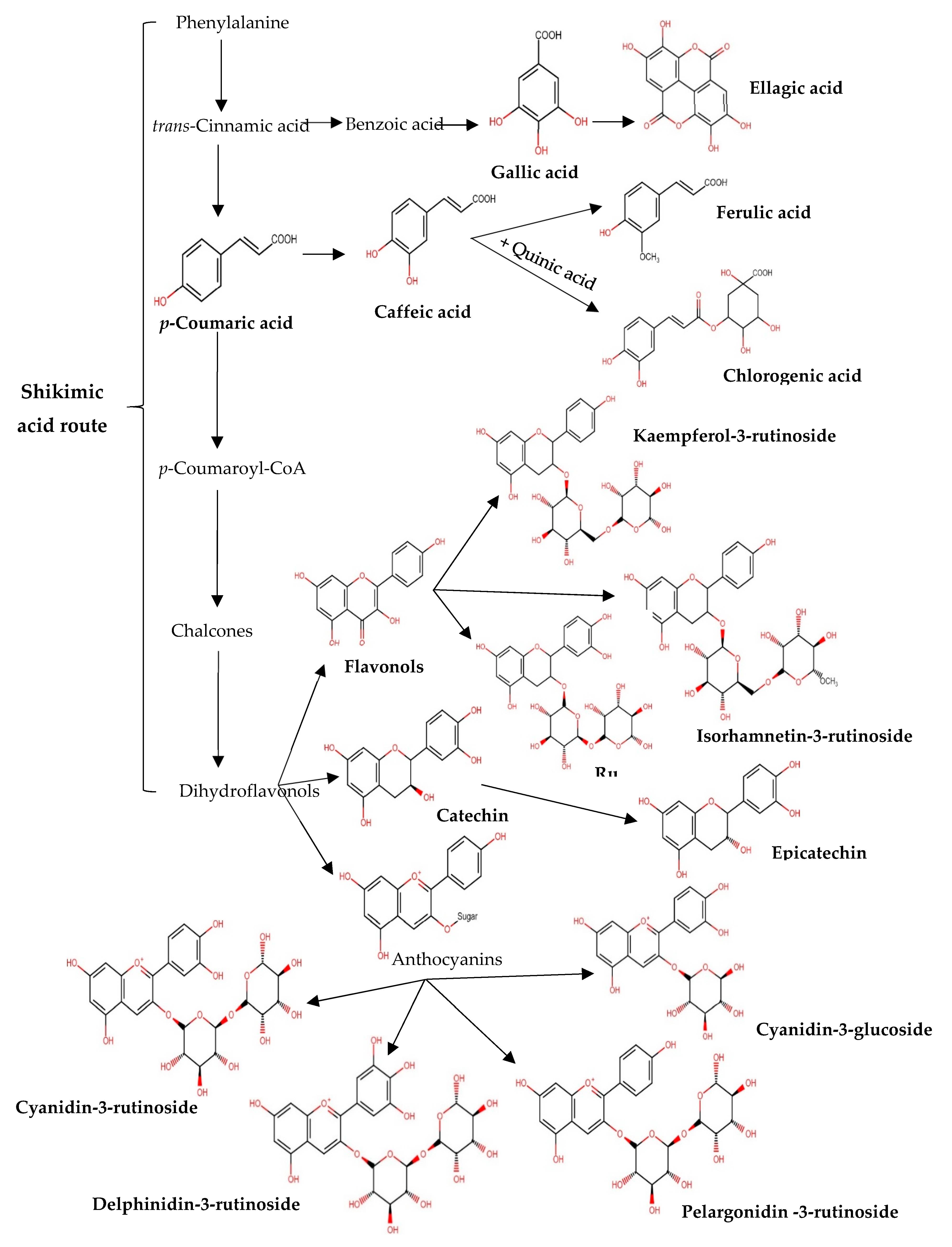
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lister, C.; Morrison, S.; Kerkhofs, N.; Wright, K. The nutritional composition and health benefits of New Zealand tamarillos. Crop Food Res. Confid. Rep. 2005, 29. [Google Scholar]
- Aitken, A.G.; Hewett, E.W. FreshFacts: New Zealand Horticulture; New Zealand Institute for Plant & Food Research: Auckland, New Zealand, 2016; p. 14. [Google Scholar]
- Acosta-Quezada, P.G.; Martínez-Laborde, J.B.; Prohens, J. Variation among tree tomato (Solanum betaceum Cav.) accessions from different cultivar groups: Implications for conservation of genetic resources and breeding. Genet. Resour. Crop Evol. 2011, 58, 943–960. [Google Scholar] [CrossRef] [Green Version]
- Espin, S.; Gonzalez-Manzano, S.; Taco, V.; Poveda, C.; Ayuda-Durán, B.; Gonzalez-Paramas, A.M.; Santos-Buelga, C. Phenolic composition and antioxidant capacity of yellow and purple-red Ecuadorian cultivars of tree tomato (Solanum betaceum Cav.). Food Chem. 2016, 194, 1073–1080. [Google Scholar] [CrossRef]
- Mutalib, M.A.; Ali, F.; Othman, F.; Ramasamy, R.; Rahmat, A. Phenolics profile and anti-proliferative activity of Cyphomandra Betacea fruit in breast and liver cancer cells. SpringerPlus 2016, 5, 2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rosso, V.V.; Mercadante, A.Z. HPLC–PDA–MS/MS of anthocyanins and carotenoids from dovyalis and tamarillo fruits. J. Agric. Food Chem. 2007, 55, 9135–9141. [Google Scholar] [CrossRef] [PubMed]
- Osorio, C.; Hurtado, N.; Dawid, C.; Hofmann, T.; Heredia-Mira, F.J.; Morales, A.L. Chemical characterisation of anthocyanins in tamarillo (Solanum betaceum Cav.) and Andes berry (Rubus glaucus Benth.) fruits. Food Chem. 2012, 132, 1915–1921. [Google Scholar] [CrossRef]
- Mertz, C.; Gancel, A.-L.; Gunata, Z.; Alter, P.; Dhuique-Mayer, C.; Vaillant, F.; Perez, A.M.; Ruales, J.; Brat, P. Phenolic compounds, carotenoids and antioxidant activity of three tropical fruits. J. Food Compos. Anal. 2009, 22, 381–387. [Google Scholar] [CrossRef]
- Kou, M.-C.; Yen, J.-H.; Hong, J.-T.; Wang, C.-L.; Lin, C.-W.; Wu, M.-J. Cyphomandra betacea Sendt. phenolics protect LDL from oxidation and PC12 cells from oxidative stress. LWT-Food Sci. Technol. 2009, 42, 458–463. [Google Scholar] [CrossRef]
- Abdul Kadir, N.A.; Rahmat, A.; Jaafar, H.Z. Protective Effects of Tamarillo (Cyphomandra betacea) Extract against High Fat Diet Induced Obesity in Sprague-Dawley Rats. J. Obes. 2015, 2015, 846041. [Google Scholar] [CrossRef]
- Mutalib, M.A.; Rahmat, A.; Ali, F.; Othman, F.; Ramasamy, R. Nutritional Compositions and Antiproliferative Activities of Different Solvent Fractions from Ethanol Extract of Cyphomandra betacea (Tamarillo) Fruit. Malays. J. Med. Sci. 2017, 24, 19–32. [Google Scholar]
- Ordóñez, R.M.; Ordóñez, A.A.; Sayago, J.E.; Moreno, M.I.N.; Isla, M.I. Antimicrobial activity of glycosidase inhibitory protein isolated from Cyphomandra betacea Sendt. fruit. Peptides 2006, 27, 1187–1191. [Google Scholar] [CrossRef] [PubMed]
- Castro-Vargas, H.I.; Benelli, P.; Ferreira, S.R.S.; Parada-Alfonso, F. Supercritical fluid extracts from tamarillo (Solanum betaceum Sendtn) epicarp and its application as protectors against lipid oxidation of cooked beef meat. J. Supercrit. Fluids 2013, 76, 17–23. [Google Scholar] [CrossRef]
- Wrolstad, R.E.; Heatherbell, D.A. Identification of anthocyanins and distribution of flavonoids in tamarillo fruit (Cyphomandra betaceae (Cav.) Sendt.). J. Sci. Food Agric. 1974, 25, 1221–1228. [Google Scholar] [CrossRef]
- Guideline ICH Harmonised Tripartite. Validation of analytical procedures: Text and methodology Q2 (R1). In Proceedings of the International Conference on Harmonization, Geneva, Switzerland, November 2005; pp. 11–12. [Google Scholar]
- Dorman, H.D.; Koşar, M.; Kahlos, K.; Holm, Y.; Hiltunen, R. Antioxidant properties and composition of aqueous extracts from Mentha species, hybrids, varieties, and cultivars. J. Agric. Food Chem. 2003, 51, 4563–4569. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Özyürek, M.; Güçlü, K.; Tütem, E.; Başkan, K.S.; Erçağ, E.; Celik, S.E.; Baki, S.; Yıldız, L.; Karaman, Ş.; Apak, R. A comprehensive review of CUPRAC methodology. Anal. Methods 2011, 3, 2439–2453. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Lucci, P.; Núñez, O.; Tundis, R.; Balzano, M.; Frega, N.G.; Conte, L.; Moret, S.; Filatova, D.; Moyano, E. Native Colombian fruits and their by-products: Phenolic profile, antioxidant activity and hypoglycaemic potential. Foods 2019, 8, 89. [Google Scholar] [CrossRef] [Green Version]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent Research on the Health Benefits of Blueberries and Their Anthocyanins. Adv. Nutr. 2019. [Google Scholar] [CrossRef]
- Corona, G.; Tang, F.; Vauzour, D.; Rodriguez-Mateos, A.; Spencer, J.P. Assessment of the anthocyanidin content of common fruits and development of a test diet rich in a range of anthocyanins. J. Berry Res. 2011, 1, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Vasco, C.; Avila, J.; Ruales, J.; Svanberg, U.; Kamal-Eldin, A. Physical and chemical characteristics of golden-yellow and purple-red varieties of tamarillo fruit (Solanum betaceum Cav.). Int. J. Food Sci. Nutr. 2009, 60, 278–288. [Google Scholar] [CrossRef]
- Acosta-Quezada, P.G.; Raigon, M.D.; Riofrio-Cuenca, T.; Garcia-Martinez, M.D.; Plazas, M.; Burneo, J.I.; Figueroa, J.G.; Vilanova, S.; Prohens, J. Diversity for chemical composition in a collection of different varietal types of tree tomato (Solanum betaceum Cav.), an Andean exotic fruit. Food Chem. 2015, 169, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Vasco, C.; Ruales, J.; Kamal-Eldin, A. Total phenolic compounds and antioxidant capacities of major fruits from Ecuador. Food Chem. 2008, 111, 816–823. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltveit, M.E. Synthesis and metabolism of phenolic compounds. In Fruit and Vegetable Phytochemicals: Chemistry, Nutritional Value and Stability; Wiley-Blackwell: Ames, IA, USA, 2010; pp. 89–100. [Google Scholar]
- Niggeweg, R.; Michael, A.J.; Martin, C. Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotechnol. 2004, 22, 746. [Google Scholar] [CrossRef]
- Gómez-Romero, M.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Metabolite profiling and quantification of phenolic compounds in methanol extracts of tomato fruit. Phytochemistry 2010, 71, 1848–1864. [Google Scholar] [CrossRef]
- Clé, C.; Hill, L.M.; Niggeweg, R.; Martin, C.R.; Guisez, Y.; Prinsen, E.; Jansen, M.A. Modulation of chlorogenic acid biosynthesis in Solanum lycopersicum; consequences for phenolic accumulation and UV-tolerance. Phytochemistry 2008, 69, 2149–2156. [Google Scholar] [CrossRef]
- Lallemand, L.A.; Zubieta, C.; Lee, S.G.; Wang, Y.; Acajjaoui, S.; Timmins, J.; McSweeney, S.; Jez, J.M.; McCarthy, J.G.; McCarthy, A.A. A structural basis for the biosynthesis of the major chlorogenic acids found in coffee. Plant Physiol. 2012, 160, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Onakpoya, I.; Spencer, E.; Thompson, M.; Heneghan, C. The effect of chlorogenic acid on blood pressure: A systematic review and meta-analysis of randomized clinical trials. J. Hum. Hypertens. 2015, 29, 77. [Google Scholar] [CrossRef]
- Tajik, N.; Tajik, M.; Mack, I.; Enck, P. The potential effects of chlorogenic acid, the main phenolic components in coffee, on health: A comprehensive review of the literature. Eur. J. Nutr. 2017, 56, 2215–2244. [Google Scholar] [CrossRef]
- Sun, T.; Ho, C.-T. Antioxidant activities of buckwheat extracts. Food Chem. 2005, 90, 743–749. [Google Scholar] [CrossRef]
- Calabro, M.; Tommasini, S.; Donato, P.; Stancanelli, R.; Raneri, D.; Catania, S.; Costa, C.; Villari, V.; Ficarra, P.; Ficarra, R. The rutin/β-cyclodextrin interactions in fully aqueous solution: Spectroscopic studies and biological assays. J. Pharm. Biomed. Anal. 2005, 36, 1019–1027. [Google Scholar] [CrossRef]
- Yang, J.; Guo, J.; Yuan, J. In vitro antioxidant properties of rutin. LWT-Food Sci. Technol. 2008, 41, 1060–1066. [Google Scholar] [CrossRef]
- Poiroux-Gonord, F.; Bidel, L.P.; Fanciullino, A.-L.; Gautier, H.; Lauri-Lopez, F.; Urban, L. Health benefits of vitamins and secondary metabolites of fruits and vegetables and prospects to increase their concentrations by agronomic approaches. J. Agric. Food Chem. 2010, 58, 12065–12082. [Google Scholar] [CrossRef]
- Ramírez, F.; Kallarackal, J. Tree tomato (Solanum betaceum Cav.) reproductive physiology: A review. Sci. Hortic. 2019, 248, 206–215. [Google Scholar] [CrossRef]
- Jang, Y.; Wang, Z.; Lee, J.-M.; Lee, J.-Y.; Lim, S. Screening of Korean natural products for anti-adipogenesis properties and isolation of kaempferol-3-O-rutinoside as a potent anti-adipogenetic compound from Solidago virgaurea. Molecules 2016, 21, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.; Visser, R.G.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables: A review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Butelli, E.; De Stefano, R.; Schoonbeek, H.-j.; Magusin, A.; Pagliarani, C.; Wellner, N.; Hill, L.; Orzaez, D.; Granell, A. Anthocyanins double the shelf life of tomatoes by delaying overripening and reducing susceptibility to gray mold. Curr. Biol. 2013, 23, 1094–1100. [Google Scholar] [CrossRef] [Green Version]
- Watson, R.R.; Schönlau, F. Nutraceutical and antioxidant effects of a delphinidin-rich maqui berry extract Delphinol®: A review. Minerva Cardioangiol. 2015, 63, 1–12. [Google Scholar]
- Czank, C.; Cassidy, A.; Zhang, Q.; Morrison, D.J.; Preston, T.; Kroon, P.A.; Botting, N.P.; Kay, C.D. Human metabolism and elimination of the anthocyanin, cyanidin-3-glucoside: A 13C-tracer study. Am. Clin. Nutr. 2013, 97, 995–1003. [Google Scholar] [CrossRef] [Green Version]
- Fang, J. Classification of fruits based on anthocyanin types and relevance to their health effects. Nutrition 2015, 31, 1301–1306. [Google Scholar] [CrossRef]
- Hurtado, N.H.; Morales, A.L.; González-Miret, M.L.; Escudero-Gilete, M.L.; Heredia, F.J. Colour, pH stability and antioxidant activity of anthocyanin rutinosides isolated from tamarillo fruit (Solanum betaceum Cav.). Food Chem. 2009, 117, 88–93. [Google Scholar] [CrossRef]
- Maier, A.; Hoecker, U. COP1/SPA ubiquitin ligase complexes repress anthocyanin accumulation under low light and high light conditions. Plant Signal. Behav. 2015, 10, e970440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Han, W.; Wang, M. Ultraviolet and environmental stresses involved in the induction and regulation of anthocyanin biosynthesis: A review. Afr. J. Biotechnol. 2008, 7. [Google Scholar]
- Jiang, M.; Ren, L.; Lian, H.; Liu, Y.; Chen, H. Novel insight into the mechanism underlying light-controlled anthocyanin accumulation in eggplant (Solanum melongena L.). Plant Sci. 2016, 249, 46–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Løvdal, T.; Olsen, K.M.; Slimestad, R.; Verheul, M.; Lillo, C. Synergetic effects of nitrogen depletion, temperature, and light on the content of phenolic compounds and gene expression in leaves of tomato. Phytochemistry 2010, 71, 605–613. [Google Scholar] [CrossRef]
- Eiro, M.J.; Heinonen, M. Anthocyanin color behavior and stability during storage: Effect of intermolecular copigmentation. J. Agric. Food Chem. 2002, 50, 7461–7466. [Google Scholar] [CrossRef]
- Marković, D.; Petranović, N.A.; Baranac, J.M. A spectrophotometric study of the copigmentation of malvin with caffeic and ferulic acids. J. Agric. Food Chem. 2000, 48, 5530–5536. [Google Scholar] [CrossRef]
- Noor Atiqah, A.; Maisarah, A.; Asmah, R. Comparison of antioxidant properties of tamarillo (Cyphomandra betacea), cherry tomato (Solanumly copersicum var. cerasiform) and tomato (Lyopersicon esulentum). Int. Food Res. J. 2014, 21. [Google Scholar]
- Ordóñez, R.M.; Cardozo, M.L.; Zampini, I.C.; Isla, M.I. Evaluation of antioxidant activity and genotoxicity of alcoholic and aqueous beverages and pomace derived from ripe fruits of Cyphomandra betacea Sendt. J. Agric. Food Chem. 2009, 58, 331–337. [Google Scholar] [CrossRef]
- Baeza, G.; Amigo-Benavent, M.; Sarriá, B.; Goya, L.; Mateos, R.; Bravo, L. Green coffee hydroxycinnamic acids but not caffeine protect human HepG2 cells against oxidative stress. Food Res. Int. 2014, 62, 1038–1046. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.W.; Piao, M.J.; Kim, K.C.; Yao, C.W.; Zheng, J.; Kim, S.M.; Hyun, C.L.; Ahn, Y.S.; Hyun, J.W. The polyphenol chlorogenic acid attenuates UVB-mediated oxidative stress in human HaCaT keratinocytes. Biomol. Ther. 2014, 22, 136. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.J.; Zhu, B.T. Inhibition of DNA methylation by caffeic acid and chlorogenic acid, two common catechol-containing coffee polyphenols. Carcinogenesis 2006, 27, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cinkilic, N.; Cetintas, S.K.; Zorlu, T.; Vatan, O.; Yilmaz, D.; Cavas, T.; Tunc, S.; Ozkan, L.; Bilaloglu, R. Radioprotection by two phenolic compounds: Chlorogenic and quinic acid, on X-ray induced DNA damage in human blood lymphocytes in vitro. Food Chem. Toxicol. 2013, 53, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Karthikesan, K.; Pari, L.; Menon, V.P. Protective effect of tetrahydrocurcumin and chlorogenic acid against streptozotocin–nicotinamide generated oxidative stress induced diabetes. J. Funct. Foods 2010, 2, 134–142. [Google Scholar] [CrossRef]
- Pavlica, S.; Gebhardt, R. Protective effects of ellagic and chlorogenic acids against oxidative stress in PC12 cells. Free Radic. Res. 2005, 39, 1377–1390. [Google Scholar] [CrossRef] [PubMed]
- Neiva, T.; Morais, L.; Polack, M.; Simoes, C.; D’amico, E. Effects of catechins on human blood platelet aggregation and lipid peroxidation. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 1999, 13, 597–600. [Google Scholar] [CrossRef]
- Jiménez-Aliaga, K.; Bermejo-Bescós, P.; Benedí, J.; Martín-Aragón, S. Quercetin and rutin exhibit antiamyloidogenic and fibril-disaggregating effects in vitro and potent antioxidant activity in APPswe cells. Life Sci. 2011, 89, 939–945. [Google Scholar] [CrossRef]
- Wang, J.; Fang, X.; Ge, L.; Cao, F.; Zhao, L.; Wang, Z.; Xiao, W. Antitumor, antioxidant and anti-inflammatory activities of kaempferol and its corresponding glycosides and the enzymatic preparation of kaempferol. PLoS ONE 2018, 13, e0197563. [Google Scholar] [CrossRef]
- Bornsek, S.M.; Ziberna, L.; Polak, T.; Vanzo, A.; Ulrih, N.P.; Abram, V.; Tramer, F.; Passamonti, S. Bilberry and blueberry anthocyanins act as powerful intracellular antioxidants in mammalian cells. Food Chem. 2012, 134, 1878–1884. [Google Scholar] [CrossRef]
- Ramirez-Tortosa, C.; Andersen, Ø.M.; Gardner, P.T.; Morrice, P.C.; Wood, S.G.; Duthie, S.J.; Collins, A.R.; Duthie, G.G. Anthocyanin-rich extract decreases indices of lipid peroxidation and DNA damage in vitamin E-depleted rats. Free Radic. Biol. Med. 2001, 31, 1033–1037. [Google Scholar] [CrossRef]
- Mazza, G.; Kay, C.D.; Cottrell, T.; Holub, B.J. Absorption of anthocyanins from blueberries and serum antioxidant status in human subjects. J. Agric. Food Chem. 2002, 50, 7731–7737. [Google Scholar] [CrossRef] [PubMed]
- Mazzucato, A.; Willems, D.; Bernini, R.; Picarella, M.E.; Santangelo, E.; Ruiu, F.; Tilesi, F.; Soressi, G.P. Novel phenotypes related to the breeding of purple-fruited tomatoes and effect of peel extracts on human cancer cell proliferation. Plant Physiol. Biochem. 2013, 72, 125–133. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Bioactive Compounds | Peel | Pulp | p-Value * | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Amber | Laird’s Large | Mulligan | Amber | Laird’s Large | Mulligan | C | T | C × T | |
| Chlorogenic acid | 225.85 ± 12.43ax | 231.18 ± 9.76bx | 278.03 ± 11.89cx | 54.67 ± 3.81ay | 66.35 ± 1.1by | 73.95 ± 1.98cy | <0.05 | <0.05 | <0.05 |
| Caffeic acid | 2.61 ± 0.18ax | 3.56 ± 0.52bx | 3.52 ± 0.85bx | 1.01 ± 0.03ay | 1.2 ± 0.06by | 1.32 ± 0.04by | <0.05 | <0.05 | 0.0328 |
| p-coumaric acid | 0.05 ± 0.01ax | 0.02 ± 0.01bx | 0.03 ± 0.01cx | 0.12 ± 0.01ay | 0.07 ± 0.01by | 0.08 ± 0.01cy | <0.05 | <0.05 | <0.05 |
| Ferulic acid | 0.03 ± 0.03ax | 0.01 ± 0.01bx | 0.04 ± 0.03cx | <0.005ay | <0.005by | 0.02 ± 0.02cy | <0.05 | <0.05 | 0.1158 |
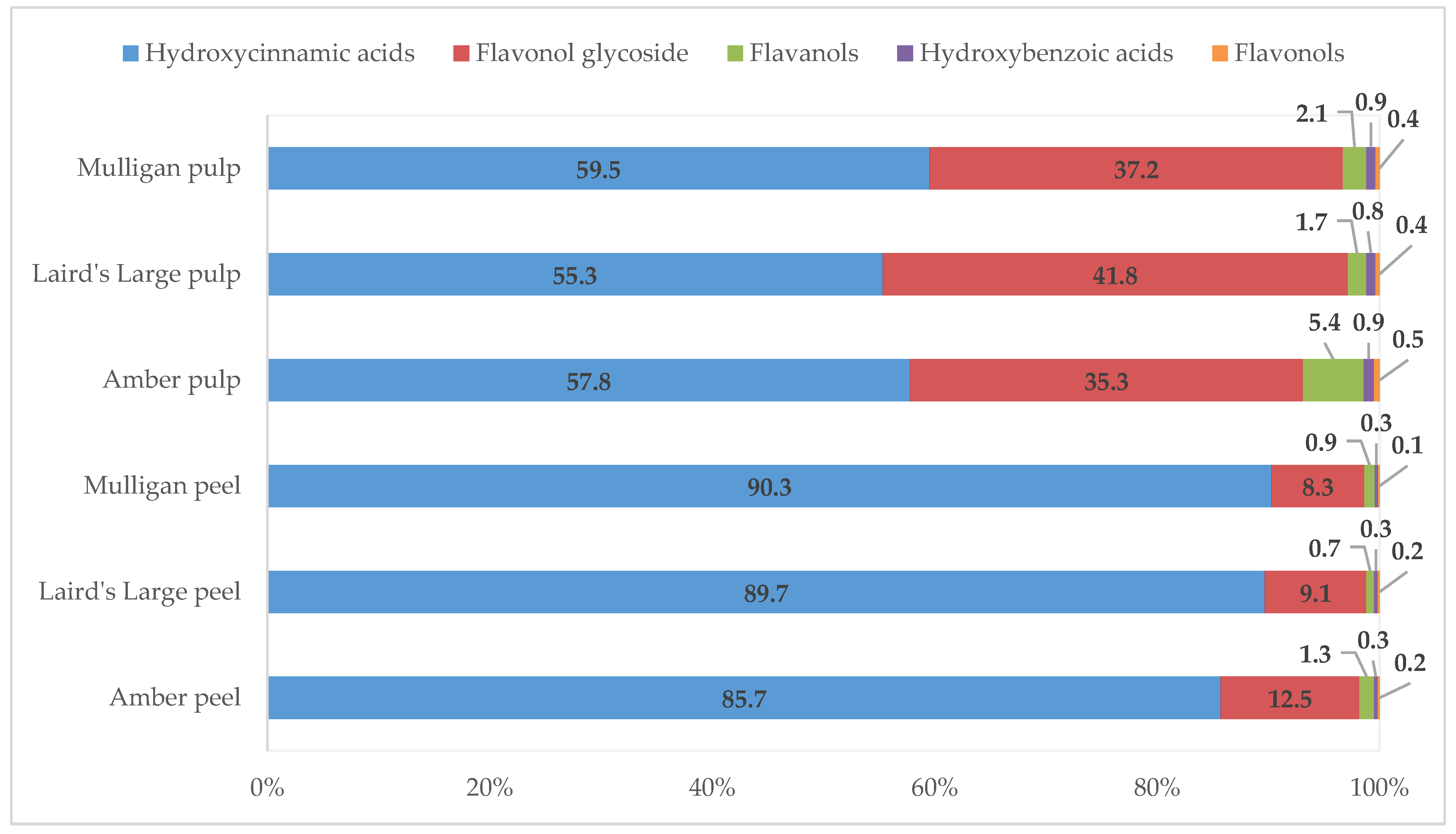
| Total hydroxycinnamic acids | 228.53 ± 12.65ax | 234.77 ± 10.3bx | 281.63 ± 12.78cx | 55.80 ± 3.86ay | 67.62 ± 1.18by | 75.37 ± 2.04cy | <0.05 | <0.05 | <0.05 |
| Gallic acid | 0.8 ± 0ax | 0.79 ± 0bx | 0.8 ± 0.01bx | 0.79 ± 0ay | 0.93 ± 0.06by | 1 ± 0.2by | <0.05 | <0.05 | <0.05 |
| Ellagic acid | 0.12 ± 0.04x | 0.11 ± 0.03x | 0.1 ± 0.04x | 0.11 ± 0.02y | 0.09 ± 0.01y | 0.09 ± 0.02y | 0.2304 | 0.0483 | 0.6023 |
| Total hydroxybenzoic acids | 0.91 ± 0.05ax | 0.9 ± 0.03abx | 0.91 ± 0.05bx | 0.90 ± 0.03ay | 1.01 ± 0.07aby | 1.09 ± 0.22by | 0.0227 | <0.05 | 0.0147 |
| Kaempferol | 0.43 ± 0.01ax | 0.43 ± 0.01bx | 0.43 ± 0.01bx | 0.5 ± 0.04ay | 0.45 ± 0.01by | 0.45 ± 0.01by | <0.05 | <0.05 | <0.05 |
| Total flavonols | 0.43 ± 0.01ax | 0.43 ± 0.01bx | 0.43 ± 0.01bx | 0.5 ± 0.04ay | 0.45 ± 0.01by | 0.45 ± 0.01by | <0.05 | <0.05 | <0.05 |
| Catechin | 2.13 ± 0.83ax | 0.28 ± 0bx | 0.33 ± 0.02bx | 3.91 ± 0.68ay | 0.3 ± 0by | 0.32 ± 0by | <0.05 | <0.05 | <0.05 |
| Epicatechin | 1.36 ± 0.03a | 1.49 ± 0.12b | 2.6 ± 0.72c | 1.34 ± 0a | 1.73 ± 0.01b | 2.31 ± 0.02c | <0.05 | 0.763 | 0.0545 |
| Total flavanols | 3.49 ± 0.86ax | 1.78 ± 0.13bx | 2.94 ± 0.74cx | 5.25 ± 0.68ay | 2.03 ± 0.01by | 2.63 ± 0.02cy | <0.05 | <0.05 | <0.05 |
| Rutin | 24.33 ± 1.85ax | 3.71 ± 0.45bx | 12.68 ± 1.1cx | 3.23 ± 0.08ay | 0.97 ± 0.03by | 1.32 ± 0.03cy | <0.05 | <0.05 | <0.05 |
| Kaempferol-3-rutinoside | 8.32 ± 0.57ax | 19.22 ± 1.2bx | 12.45 ± 1.09cx | 30.72 ± 0.97ay | 50.04 ± 1.12by | 45.6 ± 1.41cy | <0.05 | <0.05 | <0.05 |
| Isorhamnetin-3-rutinoside | 0.59 ± 0.05ax | 0.96 ± 0.05bx | 0.9 ± 0.02cx | 0.16 ± 0.01ay | 0.13 ± 0by | 0.14 ± 0cy | <0.05 | <0.05 | <0.05 |
| Total flavonol glycosides | 33.25 ± 2.47ax | 23.89 ± 1.7bx | 26.03 ± 2.21bx | 34.11 ± 1.06ay | 51.14 ± 1.14by | 47.06 ± 1.44by | <0.05 | <0.05 | <0.05 |
| Total phenolics | 266.62 ± 16.04ax | 261.77 ± 12.16bx | 311.93 ± 15.80cx | 96.56 ± 5.67ay | 122.26 ± 2.42by | 126.60 ± 3.74cy | <0.05 | <0.05 | <0.05 |
| Delphinidin-3-rutinoside | 0.43 ± 0.02ax | 32.41 ± 2.98bx | 49.11 ± 2.23cx | 29.17 ± 2.47ay | 254.76 ± 6.33by | 273.36 ± 12.7cy | <0.05 | <0.05 | <0.05 |
| Cyanidin-3-glucoside | n.d | 0.33 ± 0.04a | 1.97 ± 0.14b | n.d | n.d | n.d | <0.05 | – | – |
| Cyanidin-3-rutinoside | 0.29 ± 0ax | 68.72 ± 4.33bx | 114.47 ± 5.97cx | 0.18 ± 0.02ay | 25.94 ± 1.99by | 30.67 ± 2.76cy | <0.05 | <0.05 | <0.05 |
| Pelargonidin-3-rutinoside | 0.52 ± 0.06ax | 54.36 ± 3.24bx | 93.63 ± 2.48cx | 0.35 ± 0.03ay | 200.66 ± 8.51by | 182.81 ± 11.17cy | <0.05 | <0.05 | <0.05 |
| Total anthocyanins | 1.24 ± 0.08ax | 155.82 ± 10.58bx | 259.18 ± 10.81cx | 29.70 ± 2.52ay | 481.37 ± 16.83by | 486.84 ± 26.63cy | <0.05 | <0.05 | <0.05 |
| Tissues | Cultivars | Total Phenolic Content (mg GAE/100 g DW) | CUPRAC Value (μmol TEAC/g DW) | FRAP Value (μmol TEAC/g DW) |
|---|---|---|---|---|
| Peel | Amber | 1583.8 ± 40.09ax | 117.59 ± 9.35ax | 84.7 ± 11.13ax |
| Laird’s Large | 1673.28 ± 63.97bx | 136.68 ± 6.72bx | 102.55 ± 12.19bx | |
| Mulligan | 2225.06 ± 50.87cx | 265.29 ± 18.35cx | 161.74 ± 14.53cx | |
| Pulp | Amber | 678.98± 19.09ay | 42.92 ± 8.73ay | 52.23 ± 6.7ay |
| Laird’s Large | 707.04 ± 30.65by | 52.42 ± 8.39by | 60.19 ± 5.21by | |
| Mulligan | 874.9 ± 30.48cy | 71.57 ± 7.81cy | 72.14 ± 9.41cy | |
| p-value * | C | <0.05 | <0.05 | <0.05 |
| T | <0.05 | <0.05 | <0.05 | |
| C × T | <0.05 | <0.05 | <0.05 |
| Content | TPC | CUPRAC | FRAP | |
|---|---|---|---|---|
| TPC | Pearson’s Correlation | 1 | 0.941 ** | 0.906 ** |
| Sig. (2-tailed) | 0.000 | 0.000 | ||
| Number | 24 | 24 | 24 | |
| CUPRAC | Pearson’s Correlation | 0.941 ** | 1 | 0.959 ** |
| Sig. (2-tailed) | 0.000 | .000 | ||
| Number | 24 | 24 | 24 | |
| FRAP | Pearson’s Correlation | 0.906 ** | 0.959 ** | 1 |
| Sig. (2-tailed) | 0.000 | 0.000 | ||
| Number | 24 | 24 | 24 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diep, T.; Pook, C.; Yoo, M. Phenolic and Anthocyanin Compounds and Antioxidant Activity of Tamarillo (Solanum betaceum Cav.). Antioxidants 2020, 9, 169. https://doi.org/10.3390/antiox9020169
Diep T, Pook C, Yoo M. Phenolic and Anthocyanin Compounds and Antioxidant Activity of Tamarillo (Solanum betaceum Cav.). Antioxidants. 2020; 9(2):169. https://doi.org/10.3390/antiox9020169
Chicago/Turabian StyleDiep, Tung, Chris Pook, and Michelle Yoo. 2020. "Phenolic and Anthocyanin Compounds and Antioxidant Activity of Tamarillo (Solanum betaceum Cav.)" Antioxidants 9, no. 2: 169. https://doi.org/10.3390/antiox9020169
APA StyleDiep, T., Pook, C., & Yoo, M. (2020). Phenolic and Anthocyanin Compounds and Antioxidant Activity of Tamarillo (Solanum betaceum Cav.). Antioxidants, 9(2), 169. https://doi.org/10.3390/antiox9020169