Evaluation of Apple Pomace Flour Obtained Industrially by Dehydration as a Source of Biomolecules with Antioxidant, Antidiabetic and Antiobesity Effects

 ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
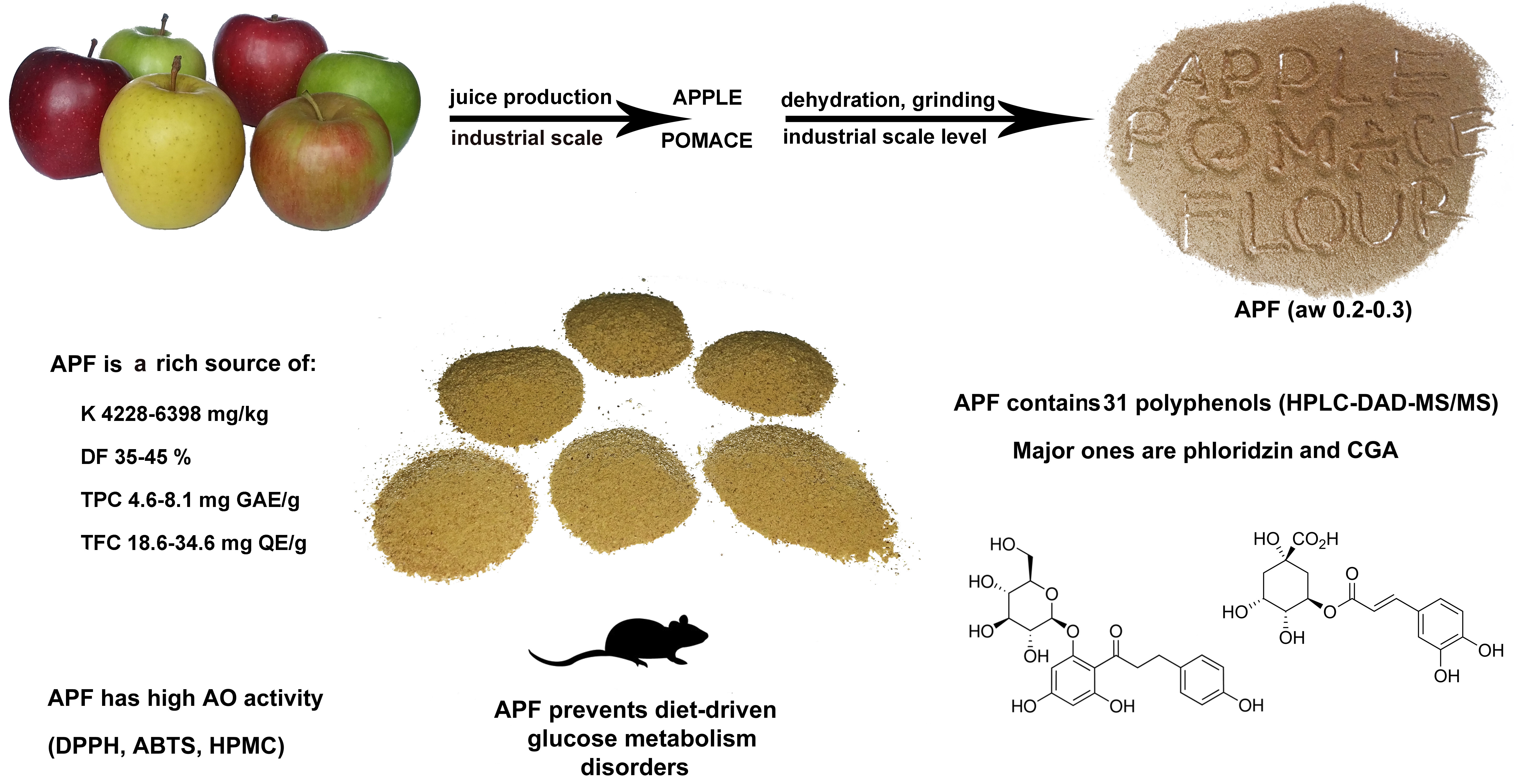
2.1.1. Apple Pomace Flour Production
2.1.2. Chemicals
2.2. Elemental Analysis
2.3. Determination of Dietary Fibre Content
2.4. Determination of Total Phenol (TPC) and Flavonoid (TFC) Content
2.5. Identification and Quantification of Phenolics by HPLC–DAD–MS/MS
2.5.1. Preparation of Samples
2.5.2. Preparation of Standards
2.5.3. Qualitative and Quantitative Analysis
2.6. Determination of AO Activity and Relative Antioxidant Capacity Index (RACI)
2.6.1. Radicals Scavenging
2.6.2. Reducing Activity
2.6.3. Relative Antioxidant Capacity Index (RACI)
2.7. Determination of Water and Oil Holding Capacity and Solubility of APF
2.7.1. Animals
2.7.2. Diets
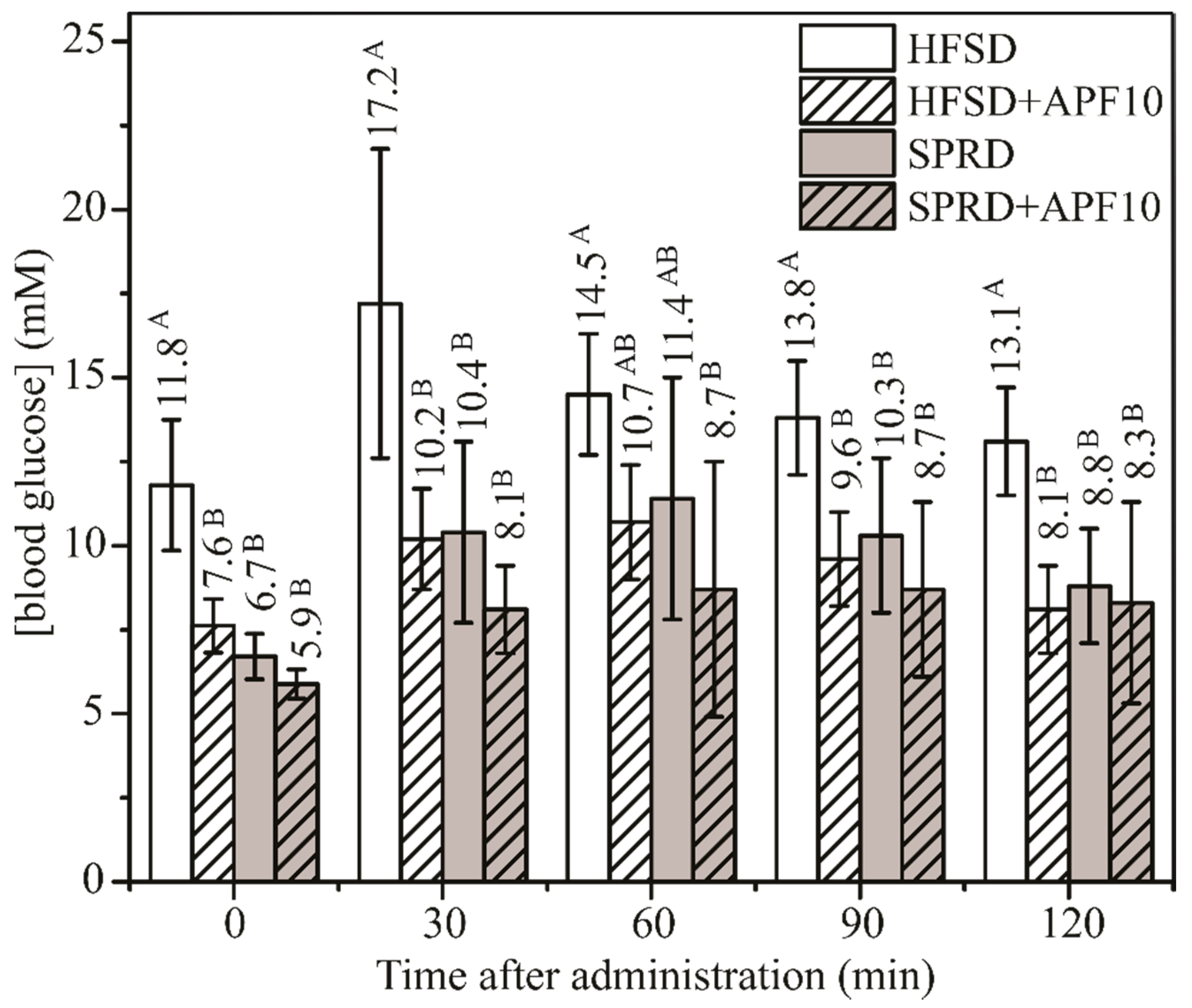
2.7.3. The Glycaemic Status
2.7.4. Oral Glucose Tolerance Test (OGTT)
2.8. Statistical Analysis
3. Results and Discussion
3.1. Production of APF
3.2. Composition of APF
3.2.1. Mineral Content
3.2.2. Content of Dietary Fibres
3.2.3. Total Phenol and Flavonoid Content
3.3. Identification and Quantification of Individual Phenolic Compounds Present in APF
3.4. Antioxidant Activity
3.5. The Correlation Analysis: Individual Phenol Contribution to Total AO Activity
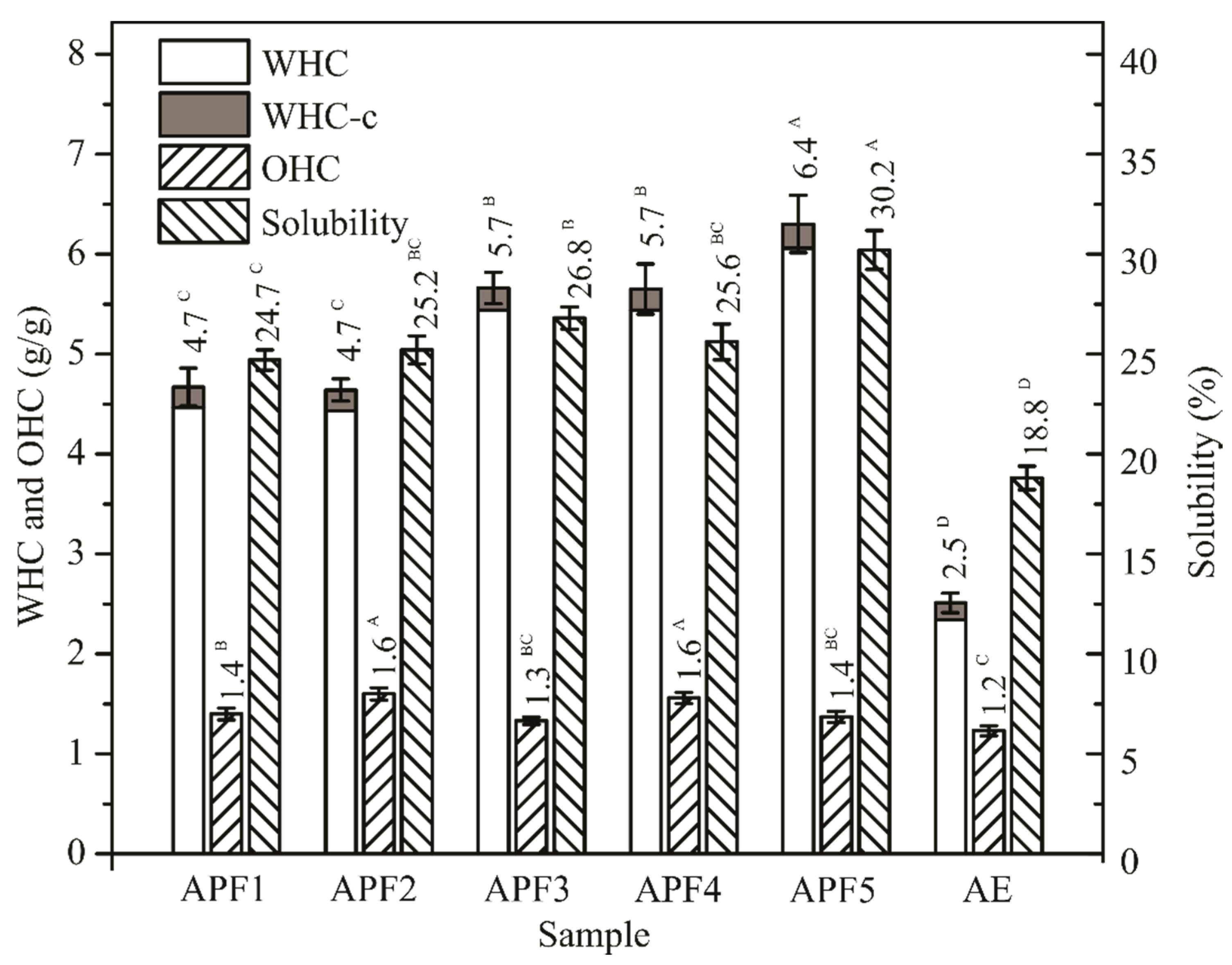
3.6. Functional Properties of APF
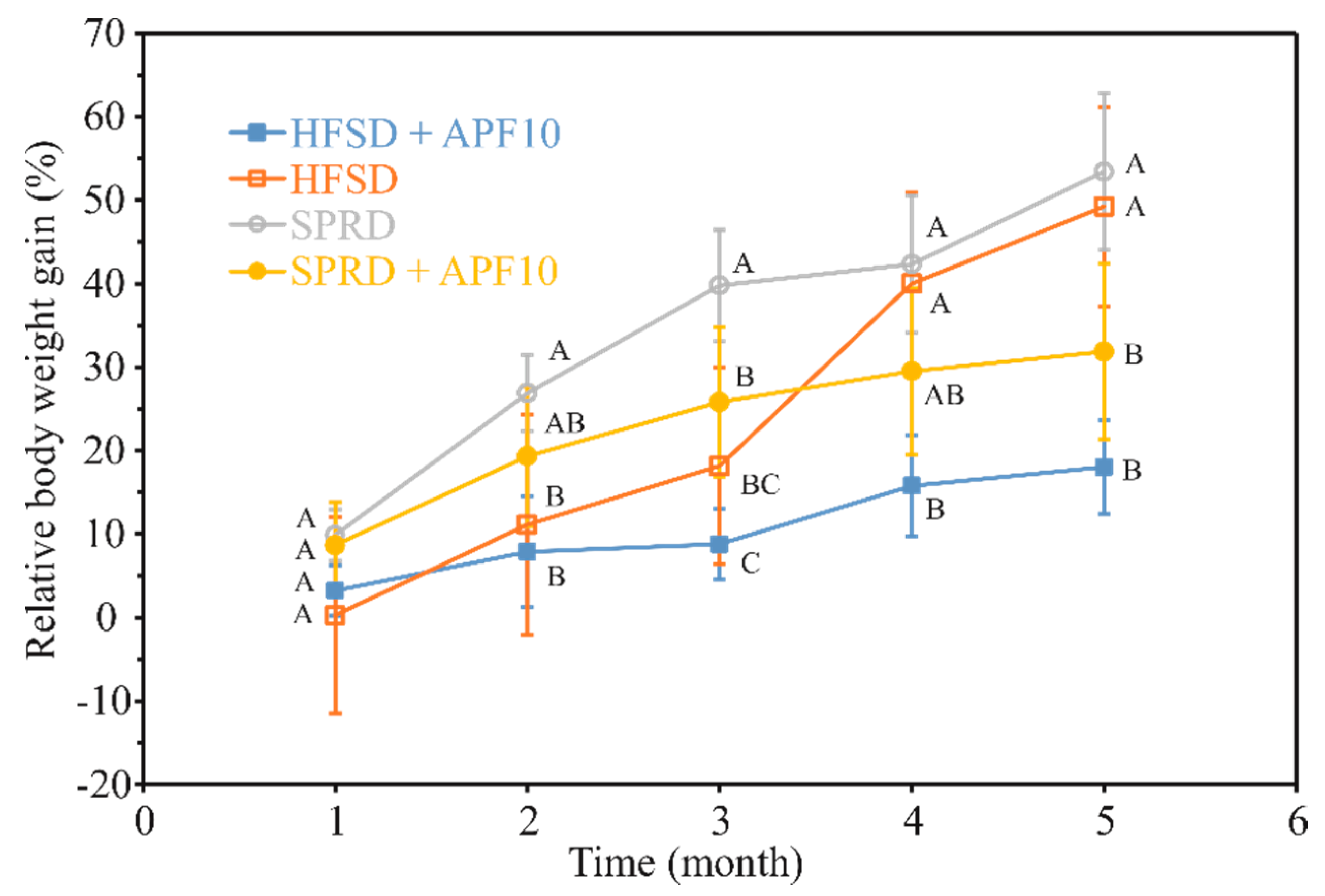
3.7. Effect of APF Presence in High-Fat and Sucrose Diet on Glucose Metabolism and Body Weight Management
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- United States Department of Agriculture Fresh Apples. Grapes, and Pears: World Markets and Trade. Foreign Agric. Serv. 2019, 1–10. [Google Scholar]
- Home | Statistical Office of the Republic of Serbia. Available online: https://www.stat.gov.rs/en-US/ (accessed on 9 September 2019).
- Skinner, R.C.; Gigliotti, J.C.; Ku, K.-M.M.; Tou, J.C. A comprehensive analysis of the composition, health benefits, and safety of apple pomace. Nutr. Rev. 2018, 76, 893–909. [Google Scholar] [CrossRef] [PubMed]
- Waldbauer, K.; McKinnon, R.; Kopp, B. Apple Pomace as Potential Source of Natural Active Compounds. Planta Med. 2017, 83, 994–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavelli, V.; Corti, S. Phloridzin and other phytochemicals in apple pomace: Stability evaluation upon dehydration and storage of dried product. Food Chem. 2011, 129, 1578–1583. [Google Scholar] [CrossRef]
- Heras-Ramírez, M.E.; Alberto, A.; Barnard, J.; Quintero-Ramos, A.; Camacho-Dávila, A.A.; Barnard, J.; Talamás-Abbud, R.; Torres-Muñoz, J.V.; Salas-Muñoz, E. Effect of Blanching and Drying Temperature on Polyphenolic Compound Stability and Antioxidant Capacity of Apple Pomace. Food Bioprocess Technol. 2012, 5, 2201–2210. [Google Scholar] [CrossRef]
- Rana, S.; Gupta, S.; Rana, A.; Bhushan, S. Functional properties, phenolic constituents and antioxidant potential of industrial apple pomace for utilization as active food ingredient. Food Sci. Hum. Wellness 2015, 4, 180–187. [Google Scholar] [CrossRef] [Green Version]
- Ratti, C. Hot air and freeze-drying of high-value foods: A review. J. Food Eng. 2001, 49, 311–319. [Google Scholar] [CrossRef]
- Zlatanović, S.; Gorjanović, S.; Ostojić, S.; Micić, D.; Pastor, F.; Kalušević, A.; Laličić-Petronijević, J. Method for producing gluten-free flour made of apple pomace. WO2020/027683, 6 February 2020. [Google Scholar]
- Sudha, M.L.; Baskaran, V.; Leelavathi, K. Apple pomace as a source of dietary fiber and polyphenols and its effect on the rheological characteristics and cake making. Food Chem. 2007, 104, 686–692. [Google Scholar] [CrossRef]
- Veselinović, N.; Veselinović, M. Solar and electric energy dryers. US2010223807 (A1), 9 September 2010. [Google Scholar]
- AOAC. AOAC Total dietary fiber in foods: Enzymatic-gravimetric method. In Official Methods of Analysis, Method 985.29; AOAC International: Arlington, VA, USA, 1990. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Kaneda, H.; Kobayashi, N.; Furusho, S.; Sahara, H.; Koshino, S. Reducing activity and flavour stability of beer. Master Brew. Assoc. Am. Tech. Q. 1995, 32, 90–94. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Petrovic, M.; Suznjevic, D.; Pastor, F.; Veljovic, M.; Pezo, L.; Antic, M.; Gorjanovic, S. Antioxidant Capacity Determination of Complex Samples and Individual Phenolics—Multilateral Approach. Comb. Chem. High Throughput Screen. 2016, 19, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Zlatanović, S.; Kalušević, A.; Micić, D.; Laličić-Petronijević, J.; Tomić, N.; Ostojić, S.; Gorjanović, S. Functionality and Storability of Cookies Fortified at the Industrial Scale with up to 75% of Apple Pomace Flour Produced by Dehydration. Foods 2019, 8, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, A.; Zhao, Y. Effect of Different Drying Methods and Storage Time on the Retention of Bioactive Compounds and Antibacterial Activity of Wine Grape Pomace (Pinot Noir and Merlot). J. Food Sci. 2012, 77, H192–H201. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Cavender, G.; Zhao, Y. Impingement drying for preparing dried apple pomace flour and its fortification in bakery and meat products. J. Food Sci. Technol. 2015, 52, 5568–5578. [Google Scholar] [CrossRef]
- Zlatanović, S.; Ostojić, S.; Micić, D.; Rankov, S.; Dodevska, M.; Vukosavljević, P.; Gorjanović, S. Thermal behaviour and degradation kinetics of apple pomace flours. Thermochim. Acta 2019, 673, 17–25. [Google Scholar] [CrossRef]
- Corey, M.E.; Kerr, W.L.; Mulligan, J.H.; Lavelli, V. Phytochemical stability in dried apple and green tea functional products as related to moisture properties. LWT Food Sci. Technol. 2011, 44, 67–74. [Google Scholar] [CrossRef]
- Hager, A.S.; Wolter, A.; Jacob, F.; Zannini, E.; Arendt, E.K. Nutritional properties and ultra-structure of commercial gluten free flours from different botanical sources compared to wheat flours. J. Cereal Sci. 2012, 56, 239–247. [Google Scholar] [CrossRef]
- Chatterjee, R.; Yeh, H.-C.; Edelman, D.; Brancati, F. Potassium and risk of Type 2 diabetes. Expert Rev. Endocrinol. Metab. 2011, 6, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Li, X.; Fan, W.; Yu, W.; Wang, S.; Li, Z.; Scott, E.M.; Li, X. Potassium and obesity/metabolic syndrome: A systematic review and meta-analysis of the epidemiological evidence. Nutrients 2016, 8, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masoodi, F.A.; Chauman, G.S. Use of Apple Pomace as A Source of Dietary Fibre in cakes. Plant Foods Hum. Nutr. 2002, 57, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Fan, M.; Ran, J.; Zhang, T.; Sun, H.; Dong, M.; Zhang, Z.; Zheng, H. Variation in phenolic compounds and antioxidant activity in apple seeds of seven cultivars. Saudi J. Biol. Sci. 2016, 23, 379–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Ying, D.; Guo, B.; Cheng, L.J.; May, B.; Bird, T.; Sanguansri, L.; Cao, Y.; Augustin, M. Extrusion of apple pomace increases antioxidant activity upon in vitro digestion. Food Funct. 2019, 10, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Gupta, A.; Issar, K. Effect of Packaging and Storage on Dried Apple Pomace and Fiber Extracted from Pomace. J. Food Process. Preserv. 2017, 41, e12913. [Google Scholar] [CrossRef]
- Lavelli, V.; Kerr, W. Apple pomace is a good matrix for phytochemical retention. J. Agric. Food Chem. 2012, 60, 5660–5666. [Google Scholar] [CrossRef]
- Lavelli, V.; Vantaggi, C. Rate of antioxidant degradation and color variations in dehydrated apples as related to water activity. J. Agric. Food Chem. 2009, 57, 4733–4738. [Google Scholar] [CrossRef]
- Rana, S.; Bhushan, S. Apple phenolics as nutraceuticals: Assessment, analysis and application. J. Food Sci. Technol. 2016, 53, 1727–1738. [Google Scholar] [CrossRef]
- Alkhalidy, H.; Wang, Y.; Liu, D. Dietary Flavonoids in the Prevention of T2D: An Overview. Nutrients 2018, 10, 438. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.A.; Subhan, N.; Hossain, H.; Hossain, M.; Reza, H.M.; Rahman, M.M.; Ullah, M.O. Hydroxycinnamic acid derivatives: A potential class of natural compounds for the management of lipid metabolism and obesity. Nutr. Metab. 2016, 13, 27. [Google Scholar] [CrossRef] [Green Version]
- Ćetković, G.; Čanadanović-Brunet, J.; Djilas, S.; Savatović, S.; Mandić, A.; Tumbas, V. Assessment of polyphenolic content and in vitro antiradical characteristics of apple pomace. Food Chem. 2008, 109, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Kim, Y.J.; Kim, D.-O.; Lee, H.J.; Lee, C.Y. Major Phenolics in Apple and Their Contribution to the Total Antioxidant Capacity. J. Agric. Food Chem. 2003, 51, 6516–6520. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Qin, C.; Zhang, P.; Ge, Q.; Wu, M.; Wu, J.; Wang, M.; Wang, Z. Antioxidant effect of apple phenolic on lipid peroxidation in Chinese-style sausage. J. Food Sci. Technol. 2013, 52, 1032–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raudone, L.; Raudonis, R.; Liaudanskas, M.; Viskelis, J.; Pukalskas, A.; Janulis, V. Phenolic Profiles and Contribution of Individual Compounds to Antioxidant Activity of Apple Powders. J. Food Sci. 2016, 81, C1055–C1061. [Google Scholar] [CrossRef]
- Plaza, M.; Kariuki, J.; Turner, C. Quantification of individual phenolic compounds’ contribution to antioxidant capacity in apple: A novel analytical tool based on liquid chromatography with diode array, electrochemical, and charged aerosol detection. J. Agric. Food Chem. 2014, 62, 409–418. [Google Scholar] [CrossRef]
- Diñeiro García, Y.; Valles, B.S.; Picinelli Lobo, A. Phenolic and antioxidant composition of by-products from the cider industry: Apple pomace. Food Chem. 2009, 117, 731–738. [Google Scholar] [CrossRef]
- Zhang, T.; Wei, X.; Miao, Z.; Hassan, H.; Song, Y.; Fan, M. Screening for antioxidant and antibacterial activities of phenolics from Golden Delicious apple pomace. Chem. Cent. J. 2016, 10, 47. [Google Scholar] [CrossRef] [Green Version]
- Alongi, M.; Melchior, S.; Anese, M. Reducing the glycemic index of short dough biscuits by using apple pomace as a functional ingredient. LWT 2019, 100, 300–305. [Google Scholar] [CrossRef]
- Weickert, M.O.; Pfeiffer, A.F. Impact of Dietary Fiber Consumption on Insulin Resistance and the Prevention of Type 2 Diabetes. J. Nutr. 2018, 148, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Skinner, R.C.; Warren, D.C.; Naveed, M.; Agarwal, G.; Benedito, V.A.; Tou, J.C. Apple pomace improves liver and adipose inflammatory and antioxidant status in young female rats consuming a Western diet. J. Funct. Foods 2019, 61, 103741. [Google Scholar] [CrossRef]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.K.; Cho, S.J.; Jung, U.J.; Ryu, R.; Choi, M.S. Phlorizin supplementation attenuates obesity, inflammation, and hyperglycemia in diet-induced obese mice fed a high-fat diet. Nutrients 2016, 8, 92. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.-J.; Lee, Y.-J.; Hwang, J.-H.; Kim, K.-J.; Lee, B.-Y. The inhibitory effects of quercetin on obesity and obesity-induced inflammation by regulation of MAPK signaling. J. Nutr. Biochem. 2015, 26, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.-D.; Han, C.-K.; Lee, B.-H. Loss of Body Weight and Fat and Improved Lipid Profiles in Obese Rats Fed Apple Pomace or Apple Juice Concentrate. J. Med. Food 2013, 16, 823–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boqué, N.; de la Iglesia, R.; de la Garza, A.L.; Milagro, F.I.; Olivares, M.; Bañuelos, Ó.; Soria, A.C.; Rodríguez-Sánchez, S.; Martínez, J.A.; Campión, J. Prevention of diet-induced obesity by apple polyphenols in Wistar rats through regulation of adipocyte gene expression and DNA methylation patterns. Mol. Nutr. Food Res. 2013, 57, 1473–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macagnan, F.T.; Santos, L.R.D.; Roberto, B.S.; De Moura, F.A.; Bizzani, M.; Da Silva, L.P. Biological properties of apple pomace, orange bagasse and passion fruit peel as alternative sources of dietary fibre. Bioact. Carbohydr. Diet. Fibre 2015, 6, 1–6. [Google Scholar] [CrossRef]
- Gonzalez, J.; Donoso, W.; Sandoval, N.; Reyes, M.; Gonzalez, P.; Gajardo, M.; Morales, E.; Neira, A.; Razmilic, I.; Yuri, J.A.; et al. Apple Peel Supplemented Diet Reduces Parameters of Metabolic Syndrome and Atherogenic Progression in ApoE−/− Mice. Evid. Based Complementary Altern. Med. 2015, 2015, 10. [Google Scholar] [CrossRef]
- Taylor, R. Calorie restriction for long-term remission of type 2 diabetes. Clin. Med. J. R. Coll. Physicians Lond. 2019, 19, 37–42. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority). Dietary Reference Values for nutrients Summary report. EFSA Support. Publ. 2017, 14, e15121. [Google Scholar]
- Najafian, M.; Jahromi, M.Z.; Nowroznejhad, M.J.; Khajeaian, P.; Kargar, M.M.; Sadeghi, M.; Arasteh, A. Phloridzin reduces blood glucose levels and improves lipids metabolism in streptozotocin-induced diabetic rats. Mol. Biol. Rep. 2012, 39, 5299–5306. [Google Scholar] [CrossRef]
- Niederberger, K.E.; Tennant, D.R.; Bellion, P. Dietary intake of phloridzin from natural occurrence in foods. Br. J. Nutr. 2020, 123, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Nishimuro, H.; Ohnishi, H.; Sato, M.; Ohnishi-Kameyama, M.; Matsunaga, I.; Naito, S.; Ippoushi, K.; Oike, H.; Nagata, T.; Akasaka, H.; et al. Estimated Daily Intake and Seasonal Food Sources of Quercetin in Japan. Nutrients 2015, 7, 2345–2358. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Gu, Y.; Zhang, Q.; Liu, L.; Meng, G.; Wu, H.; Xia, Y.; Bao, X.; Shi, H.; Sun, S.; et al. Estimated daily quercetin intake and association with the prevalence of type 2 diabetes mellitus in Chinese adults. Eur. J. Nutr. 2019, 58, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Saura-Calixto, F. Dietary fiber as a carrier of dietary antioxidants: An essential physiological function. J. Agric. Food Chem. 2011, 59, 43–49. [Google Scholar] [CrossRef]
- Wu, L.; Sanguansri, L.; Ann Augustin, M. Protection of epigallocatechin gallate against degradation during in vitro digestion using apple pomace as a carrier. J. Agric. Food Chem. 2014, 62, 12265–12270. [Google Scholar] [CrossRef]
- Aprikian, O.; Duclos, V.; Guyot, S.; Besson, C.; Manach, C.; Bernalier, A.; Morand, C.; Rémésy, C.; Demigné, C. Apple Pectin and a Polyphenol-Rich Apple Concentrate Are More Effective Together Than Separately on Cecal Fermentations and Plasma Lipids in Rats. J. Nutr. 2003, 133, 1860–1865. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TPC FC (mg GAE g−1) | TFC (mg QE g−1) | |
|---|---|---|
| APF1 | 7.7 ± 0.3 a | 24.8 ± 1.0 bc |
| APF2 | 6.1 ± 0.2 b | 27.4 ± 1.4 b |
| APF3 | 8.1 ± 0.3 a | 34.6 ± 2.2 a |
| APF4 | 4.6 ± 0.2 c | 18.6 ± 1.0 d |
| APF5 | 4.6 ± 0.1 c | 21.2 ± 1.3 cd |
| AE | 4.3 ± 0.2 c | 12.2 ± 0.7 e |
| F | 163.3 | 97.1 |
| p | <0.001 | <0.001 |
| mg kg−1 | APF1 | APF2 | APF3 | APF4 | APF5 | AE | F | p |
|---|---|---|---|---|---|---|---|---|
| Dihydrochalcones | ||||||||
| Phloretin | 0.77 ± 0.06 b | 0.70 ± 0.04 bc | 0.98 ± 0.03 a | 0.29 ± 0.02 d | 0.78 ± 0.00 b | 0.63 ± 0.05 c | 102.3 | <0.001 |
| Phlorizin | 215.1 ± 2.5 a | 194.5 ± 5.3 b | 227.3 ± 1.3 a | 112 ± 3.0 d | 165.8 ± 1.1 c | 158.9 ± 9.4 c | 240.4 | <0.001 |
| Total | 215.9 ± 2.5 a | 195.2 ± 5.3 b | 228.3 ± 1.3 a | 112 ± 3 d | 166.3 ± 1.3 c | 159.6 ± 9.4 c | 239.9 | <0.001 |
| Flavones | ||||||||
| Luteolin | 0.26 ± 0.01 a | 0.11 ± 0.01 c | 0.15 ± 0.01 b | 0.10 ± 0.01 c | 0.13 ± 0.01 bc | 0.11 ± 0.01 c | 89.1 | <0.001 |
| Apigenin-7-O-glucoside | 0.84 ± 0.05 b | 0.73 ± 0.02 bc | 1.01 ± 0.05 a | 0.68 ± 0.03 c | 0.47 ± 0.04 d | 0.69 ± 0.06 c | 49.6 | <0.001 |
| Apigenin | 0.48 ± 0.00 | 0.38 ± 0.04 | 0.46 ± 0.07 | 0.31 ± 0.01 | 0.41 ± 0.23 | 0.43 ± 0.21 | 0.6 | 0.675 |
| Chrysin | 0.19 ± 0.00 b | 0.18 ± 0.00 c | 0.22 ± 0.00 a | 0.13 ± 0.00 d | 0.11 ± 0.00 e | 0.12 ± 0.00 d | 702.9 | <0.001 |
| Total | 1.77 ± 0.07 a | 1.40 ± 0.07 ab | 1.84 ± 0.11 a | 1.22 ± 0.03 b | 1.12 ± 0.28 b | 1.35 ± 0.26 ab | 6.9 | 0.003 |
| Flavanones | ||||||||
| Eriodictyol | 0.18 ± 0.02 b | 0.13 ± 0.02 c | 0.26 ± 0.01 a | 0.11 ± 0.01 c | 0.21 ± 0.02 b | 0.21 ± 0.01 b | 34.0 | <0.001 |
| Naringenin | 0.24 ± 0.02 a | 0.18 ± 0.03 b | 0.21 ± 0.01 ab | 0.11 ± 0.00 c | 0.17 ± 0.00 b | 0.19 ± 0.01 ab | 20.2 | <0.001 |
| Naringin | 0.22 ± 0.01 d | 0.57 ± 0.02 a | 0.60 ± 0.01 a | 0.48 ± 0.02 b | 0.35 ± 0.02 c | 0.46 ± 0.02 b | 195.0 | <0.001 |
| Total | 0.64 ± 0.01 d | 0.88 ± 0.01 b | 1.07 ± 0.01 a | 0.70 ± 0.03 c | 0.73 ± 0.04 c | 0.86 ± 0.00 b | 177.5 | <0.001 |
| Flavonols and Flavonol Glycosides | ||||||||
| Quercetin | 14.2 ± 3.0 a | 10.9 ± 2.4 ab | 13.1 ± 3.3 ab | 7.2 ± 1.8 bc | 4.10 ± 0.74 c | 12.7 ± 1.8 ab | 8.6 | 0.001 |
| Quercetin-3-O-rhamnoside | 121.9 ± 1.3 a | 85.3 ± 4.1 c | 124.8 ± 1.7 a | 114.1 ± 3.9 b | 34.1 ± 1.3 d | 85.9 ± 1.7 c | 512.7 | <0.001 |
| Quercetin-3-O-galactoside | 165.2 ± 3.5 a | 158.5 ± 2.7 a | 149.9 ± 4.1 b | 126.7 ± 3.4 c | 121.4 ± 0.6 c | 80.8 ± 1.8 d | 339.6 | <0.001 |
| Rutin | 46.93 ± 1.91 b | 20.37 ± 0.28 e | 23.9 ± 0.2 d | 64.86 ± 1.40 a | 7.99 ± 0.09 f | 34.46 ± 0.84 c | 1163.2 | <0.001 |
| Isorhamnetin-3-O-rutinoside | 1.11 ± 0.01 a | 0.82 ± 0.02 b | 0.41 ± 0.00 c | 0.40 ± 0.04 c | 0.36 ± 0.04 c | 0.10 ± 0.01 d | 703.8 | <0.001 |
| Isorhamnetin | 12.31 ± 0.43 b | 17.62 ± 0.31 a | 4.05 ± 0.04 c | 2.08 ± 0.02 d | 1.10 ± 0.01 e | 1.16 ± 0.02 e | 3037.8 | <0.001 |
| Kaempferol | 2.46 ± 0.37 a | 0.71 ± 0.14 bc | 2.82 ± 0.37 a | 1.37 ± 0.16 b | 0.48 ± 0.02 c | 0.62 ± 0.42 bc | 37.1 | <0.001 |
| Kaempferol-7-O-glucoside | 0.05 ± 0.01 c | 0.03 ± 0.01 c | 0.70 ± 0.03 b | 1.19 ± 0.12 a | 0.11 ± 0.01 c | 0.13 ± 0.02 c | 251.7 | <0.001 |
| Total | 363.9 ± 7.8 a | 257.5 ± 7.3 c | 328.4 ± 8.6 b | 341.1 ± 7.4 b | 129.1 ± 2.5 d | 261.8 ± 4.4 c | 491.4 | <0.001 |
| Flavanonols | ||||||||
| Taxifolin | 0.16 ± 0.02 d | 0.46 ± 0.01 a | 0.33 ± 0.01 b | 0.33 ± 0.04 b | 0.24 ± 0.01 c | 0.21 ± 0.02 cd | 86.5 | <0.001 |
| Hydroxycinnamic acids | ||||||||
| Caffeic acid | 0.33 ± 0.02 a | 0.22 ± 0.03 b | 0.35 ± 0.05 a | 0.23 ± 0.01 b | 0.12 ± 0.00 c | 0.18 ± 0.00 bc | 39.7 | <0.001 |
| p-Coumaric acid | 0.32 ± 0.06 c | 0.44 ± 0.09 bc | 0.44 ± 0.03 bc | 0.76 ± 0.09 a | 0.57 ± 0.03 b | 0.51 ± 0.05 b | 17.5 | <0.001 |
| Ferulic acid | 23.80 ± 0.28 a | 23.43 ± 0.87 a | 13.24 ± 0.07 d | 23.48 ± 0.21 a | 19.48 ± 0.54 b | 15.86 ± 0.33 c | 281.1 | <0.001 |
| Sinapic acid | 7.20 ± 0.03 a | 4.29 ± 0.19 b | 2.97 ± 0.05 c | 2.78 ± 0.03 cd | 2.60 ± 0.01 d | 2.03 ± 0.05 e | 1464.3 | <0.001 |
| Chlorogenic acid | 224.4 ± 9.8 b | 214.3 ± 3.4 b | 308.3 ± 14.0 a | 89.0 ± 9.5 e | 185.7 ± 6.2 c | 126.6 ± 4.7 d | 237.4 | <0.001 |
| Total | 251.8 ± 10.1 b | 245.6 ± 3.9 b | 325.1 ± 14.1 a | 117.7 ± 9.5 e | 208.5 ± 6.4 c | 145.2 ± 4.6 d | 220.9 | <0.001 |
| Hydroxybenzoic acids | ||||||||
| Gallic acid | 4.53 ± 0.10 b | 2.22 ± 0.04 e | 4.80 ± 0.12 a | 3.20 ± 0.12 c | 3.20 ± 0.03 c | 2.54 ± 0.03 d | 450.4 | <0.001 |
| Protocatechuic acid | 28.61 ± 1.01 a | 7.29 ± 0.55 de | 21.15 ± 0.73 b | 6.58 ± 0.42 e | 17.22 ± 0.24 c | 8.69 ± 0.01 d | 686.9 | <0.001 |
| Ellagic acid | 19.70 ± 2.90 b | 14.17 ± 0.27 b | 24.04 ± 5.64 b | 22.88 ± 8.49 b | 22.49 ± 2.30 b | 75.2 ± 11.8 a | 35.5 | <0.001 |
| p-Hydroxybenzoic acid | 2.51 ± 0.10 b | 2.15 ± 0.38 b | 2.24 ± 0.15 b | 2.92 ± 0.53 b | 1.16 ± 0.17 c | 5.80 ± 0.09 a | 89.9 | <0.001 |
| Total | 55.4 ± 3.4 b | 25.8 ± 0.5 d | 52.2 ± 5.2 bc | 35.6 ± 9.4 cd | 44.1 ± 1.9 bcd | 92.3 ± 11.7 a | 35.6 | <0.001 |
| Coumarins | ||||||||
| Aesculin | 9.38 ± 0.37 b | 8.80 ± 0.36 bc | 10.67 ± 0.56 a | 7.96 ± 0.11 c | 9.68 ± 0.28 b | 5.53 ± 0.16 d | 81.8 | <0.001 |
| Others | ||||||||
| Resveratrol | 0.16 ± 0.01 d | 0.89 ± 0.02 a | 0.82 ± 0.01 b | 0.24 ± 0.02 c | 0.22 ± 0.02 c | 0.19 ± 0.03 cd | 869.1 | <0.001 |
| Pterostilbene | 0.19 ± 0.01 d | 0.90 ± 0.00 a | 0.70 ± 0.01 b | 0.35 ± 0.02 c | 0.29 ± 0.02 c | 0.20 ± 0.05 d | 478.3 | <0.001 |
| Pinocembrin | 0.39 ± 0.01 a | 0.32 ± 0.00 b | 0.22 ± 0.00 e | 0.25 ± 0.00 d | 0.29 ± 0.00 c | 0.22 ± 0.00 e | 938.6 | <0.001 |
| Total | ||||||||
| 899.9 ± 3.9 b | 736.1 ± 7.2 c | 948.6 ± 77.0 a | 619 ± 10 e | 560.5 ± 4.9 f | 668.6 ± 19.9d | 736.5 | <0.001 | |
| ABTS (mmol TE 100 g−1) | DPPH (mmol TE 100 g−1) | HPMC (%/mg) | RACI | |
|---|---|---|---|---|
| APF1 | 10.0 ± 0.7 a | 3.8 ± 0.2 ab | 1.20 ± 0.04 b | 0.63 |
| APF2 | 9.2 ± 0.9 a | 3.3 ± 0.3 bc | 1.20 ± 0.04 b | 0.35 |
| APF3 | 9.5 ± 1.0 a | 4.5 ± 0.4 a | 1.74 ± 0.08 a | 1.42 |
| APF4 | 3.6 ± 0.5 b | 2.6 ± 0.2 cd | 1.00 ± 0.04 c | −0.68 |
| APF5 | 3.4 ± 0.3 b | 2.9 ± 0.4 bcd | 0.90 ± 0.04 cd | −0.68 |
| AE | 3.1 ± 0.4 b | 2.2 ± 0.5 d | 0.86 ± 0.04 d | −1.03 |
| F | 74.7 | 17.1 | 130.5 | |
| p | <0.001 | <0.001 | <0.001 |
| Variables | DPPH | HPLC | TPC | TFC | HPMC | RACI |
|---|---|---|---|---|---|---|
| ABTS | 0.86 | 0.89 | 0.94 | 0.82 | 0.77 | 0.93 |
| DPPH | 0.89 | 0.96 | 0.95 | 0.93 | 0.98 | |
| HPLC | 0.97 | 0.75 | 0.86 | 0.93 | ||
| TPC | 0.86 | 0.88 | 0.97 | |||
| TFC | 0.92 | 0.94 | ||||
| HPMC | 0.95 |
| Parameter | HFSD + APF10 | HFSD | SPRD | SPRD + APF10 | F | p |
|---|---|---|---|---|---|---|
| Glycemia (mmol/L) | 7.6 ± 1.5 b | 12.4 ± 2.1 a | 8.9 ± 2.3 b | 8.9 ± 0.6 b | 8.3 | 0.001 |
| Body weight gain (g) | 4.5 ± 1.3 b | 11.5 ± 2.3 a | 11.1 ± 1.3 a | 6.7 ± 1.9 b | 22.5 | <0.001 |
| Water intake (mL/d) | 8.56 ± 0.36 | 8.44 ± 0.42 | 8.56 ± 0.36 | 8.39 ± 0.23 | 0.28 | 0.838 |
| Food intake (g/d) | 1.99 ± 0.03 b | 1.97 ± 0.05 b | 2.12 ± 0.03 a | 2.10 ± 0.04 a | 20.5 | <0.001 |
| Energy intake (kcal/d) | 15.2 ± 0.2 a | 15.0 ± 0.3 a | 6.4 ± 0.1 b | 6.3 ± 0.1 b | 3559.7 | <0.001 |
| FER1 ** (g/g) | 0.015 ± 0.004 b | 0.039 ± 0.008 a | 0.035 ± 0.004 a | 0.021 ± 0.006 b | 19.2 | <0.001 |
| FER2 *** (×10−3 g/kcal) | 2.0 ± 0.6 c | 5.1 ± 1.0 b | 11.6 ± 1.4 a | 7.1 ± 2.0 b | 53.3 | <0.001 |
| AUC (mM*h) | 19.2 ± 2.0 b | 29.0 ± 3.6 a | 16.3 ± 3.7 b | 19.9 ± 4.5 b | 14.3 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorjanović, S.; Micić, D.; Pastor, F.; Tosti, T.; Kalušević, A.; Ristić, S.; Zlatanović, S. Evaluation of Apple Pomace Flour Obtained Industrially by Dehydration as a Source of Biomolecules with Antioxidant, Antidiabetic and Antiobesity Effects. Antioxidants 2020, 9, 413. https://doi.org/10.3390/antiox9050413
Gorjanović S, Micić D, Pastor F, Tosti T, Kalušević A, Ristić S, Zlatanović S. Evaluation of Apple Pomace Flour Obtained Industrially by Dehydration as a Source of Biomolecules with Antioxidant, Antidiabetic and Antiobesity Effects. Antioxidants. 2020; 9(5):413. https://doi.org/10.3390/antiox9050413
Chicago/Turabian StyleGorjanović, Stanislava, Darko Micić, Ferenc Pastor, Tomislav Tosti, Ana Kalušević, Slavica Ristić, and Snežana Zlatanović. 2020. "Evaluation of Apple Pomace Flour Obtained Industrially by Dehydration as a Source of Biomolecules with Antioxidant, Antidiabetic and Antiobesity Effects" Antioxidants 9, no. 5: 413. https://doi.org/10.3390/antiox9050413