1. Introduction
Diabetes pathology is associated with hyperglycemia and an enhanced risk of developing cardiovascular disease [
1]. Several metabolic and signaling pathways become dysfunctional and contribute to chronic disorders associated with diabetes. For instance, oxidative stress and glycation leading to advanced glycation end-products (AGEs) formation, were identified as key mediators of multiple complications occurring in diabetes [
2]. The generation of AGEs results from a complex process, named glycoxidation and involves a condensation reaction between proteins and glucose, followed by Amadori rearrangement, cyclization, polymerization, cleavage and oxidation process [
3]. Glycoxidation process can also concern protein reaction with highly reactive α-oxoaldehydes derivatives such as methylglyoxal (MGO) occurring in glucose autoxidation [
4]; AGEs derived from these precursors are responsible for several organ and cell toxicities [
5]. Plasmatic proteins and blood cells are the major targets of oxidation and glycation affecting mainly long half-life and abundant blood proteins such as albumin and hemoglobin [
6]. Albumin was considered as a preferential protein target for glycoxidation because of its long half-life, its high abundance in serum (35 g/L), its continuous exposure to oxidative stress and hyperglycemia and the presence of multiple glycation sites [
6]. The glycoxidation process of this globular protein exerts adverse effects on its structural and functional integrity, particularly in terms of conformational changes and redox status, which may favor the formation of cross β-structure rich amyloid fibril [
7,
8]. Our group recently evidenced impaired drug binding capacity of albumin when glycoxidized [
9,
10]. Glycoxidized albumin was associated with metabolic disorders observed in diabetes mellitus, such as retinopathy, nephropathy and coronary artery disease [
11].
During their life span, erythrocytes are also continuously exposed to glucose and oxidative stress. In hyperglycemic condition, red blood cells are particularly concerned by hemoglobin glycation and AGEs abundant accumulations [
12]. In red blood cells, glycoxidation phenomenon can lead to an increased oxidative stress, aggregation, membrane phospholipids asymmetry resulting from eryptosis and to a decreased deformability and elasticity capacity [
13]. These hemorrheologic impairments, associated with an induced early senescence phenomenon, render red blood cells (RBC) more prone to being phagocytized and potentially participate in atheroma/vascular complications [
14,
15,
16].
Among the potential therapeutic interventions to manage diabetes and prevent for associated complications, polyphenol-rich diet from natural products appears to be a high interest approach [
17]. Plant polyphenols constitute the main dietary source of antioxidants which exhibit several other beneficial biological properties including antibacterial, antiviral and anti-inflammatory activities [
18,
19].
Endemic to Mascarene and Madagascar,
Antirhea borbonica (Rubiaceae) is a tree mainly found in the wild and humid forests of Reunion island, a French overseas territory. Formerly used in Reunion, the leaves were crushed and applied in poultices to improve healing and stop bleeding or consumed as a decoction for their astringent properties to fight against diarrhea, dysentery and bladder problems [
20]. Registered in the French Pharmacopoeia, this protected species is cultivated and sold (≈16 €/100 g) on a small scale mainly at La Reunion island [
20]. Today, leaf decoctions remain consumed by Mauritians for their astringent properties [
21,
22] while the Reunionese use it in the form of herbal teas to treat diabetes mellitus and hypercholesterolemia [
23].
Recently, published data from our group evidenced polyphenols derived from the medicinal plant
Antirhea borbonica exhibit robust anti-inflammatory properties at the preadipocyte and adipocyte levels [
24,
25]. The benefits of this plant in terms of antioxidative and antiglycative properties remained uncovered. In this study, we hypothesized that the
Antirhea borbonica (
Ab) aqueous plant extracts exert antiglycation and antioxidant protective effects for albumin and RBC subjected to dicarbonyl-induced toxicity. For albumin, the main plasmatic protein, multistage glycation markers (ketoamine, free amine level and fluorescent AGE), along with oxidative and conformational parameters (thiols, β-aggregation and tryptophan fluorescence), were investigated. For the main circulating blood cells, morphology, deformability, oxidative stress and eryptosis parameters were evaluated. Finally, the antioxidant effects of
Ab in red blood cells were also investigated in an in vivo model of MGO-injected zebrafish.
In summary, Ab aqueous extracts revealed to be rich in polyphenols and potent inhibitor of oxidative damages for albumin and erythrocytes submitted to MGO-induced damages. Antioxidant capacities of Ab extract on erythrocytes were retrieved in vivo in zebrafish previously infused with MGO. These results reveal antioxidant and antiglycant properties of Ab that could be used for dietary modulation of oxidative stress and glycation in hyperglycemic situations.
2. Materials and Methods
2.1. Plant Materials and Preparation of Plant Extracts
Leaves of Antirhea borbonica J.F Gmel (Bois d’osto; Saint-Joseph de La Réunion; REF: BOSJDTCA171218AA) obtained from the Cooperative des Huiles Essentielles de Bourbon (CAHEB) (Saint-Pierre de La Réunion, France) were dried, crushed and conserved at –20 °C until extraction. Aqueous plant extract (or herbal tea) (4 g/L) was prepared by infusion technique. Briefly, 1 g of crushed plant was mixed with 250 mL of boiled Milli-Q water (or fish water) for 10 min. The herbal tea preparation was filtered (with 20 µm membrane), aliquoted and stored at –80 °C before use. For zebrafish treatment, the fresh herbal tea was directly diluted in 2 L of fish water to reach a final concentration of 0.5 g/L.
2.2. Identification of Polyphenols in Medicinal Plant Extract
Polyphenols extracted from Antirhea borbonica infusion were identified by Ultra-High Performance Liquid Chromatography coupled with diode array detection and HESI-Orbitrap mass spectrometer (Q Exactive Plus, Thermo Fisher, Waltham, MA, United States). Briefly, 10 µL of sample was injected using an UHPLC system equipped with a Thermo Fisher Ultimate 3000 series WPS-3000 RS autosampler and then separated on a PFP column (2.6 µm, 100 mm × 2.1 mm, Phenomenex, Torrance, CA, United States). Elution of the column was conducted by using a gradient combination of 0.1% formic acid in water (A) and 0.1% formic acid in acetonitrile (B) at the flow rate of 0.450 mL/min, with 5% B at 0–0.1 min, 35% B at 0.1–7.1 min, 95% B at 7.1–7.2 min, 95% B at 7.2–7.9 min, 5% B at 7.9–8 min and 5% B at 8–10 min. The column temperature was held at 30 °C and the detection wavelength was set to 280 nm and 320 nm.
For the mass spectrometer conditions, a Heated Electrospray Ionization Source II (HESI II) was used. Nitrogen was used as drying gas. The mass spectrometric conditions were optimized as follows—spray voltage = 2.8 kV, capillary temperature = 350 °C, sheath gas flow rate = 60 units, aux gas flow rate = 20 units and S lens RF level = 50.
Mass spectra were registered in full scan mode from m/z 100 to 1500 in negative ion mode at a resolving power of 70,000 FWHM (full width at half maximum) at m/z 400. The automatic gain control (AGC) was set at 1e6. The Orbitrap performance in negative ionization mode was evaluated weekly and external calibration of the mass spectrometer was performed with a LTQ ESI negative ion calibration solution (Pierce™, Waltham, MA, United States). Identification of the compounds of interest was based on their exact mass, retention time and MS/MS analysis. Data were acquired and processed by XCalibur 4.0 software (Thermo Fisher Scientific Inc., Waltham, MA, United States).
2.3. Quantification and Identification of Polyphenols in Medicinal Plant Extracts
Polyphenol contents in herbal tea preparation of
Antirhea borbonica were determined using the Folin-Ciocalteu test [
26]. Briefly, 25 μL plant extract (1:10 dilution), 125 μL Folin-Ciocalteu’s phenol reagent (Sigma-Aldrich, Darmstadt, Germany, 1:10 dilution in water) and 100 μL sodium carbonate solution (75 g/L) were mixed in a ninety six-well microplate and sequentially incubated at 50 °C for 5 min and then at 4 °C for 5 min, in the dark before absorbance measurement at 760 nm (FLUOstar Optima, BMG Labtech, Ortenberg, Germany). The total phenolic content was calculated with respect to the gallic acid (Sigma-Aldrich, Darmstadt, Germany) calibration curve (calibration range 12.5–300 µM). Results were expressed as mg gallic acid equivalent (GAE) per g plant powder.
Total flavonoids were also measured using the aluminum chloride (AlCl
3) colorimetric assay and were adapted from Zhishen et al. [
27]. For this measurement, 100 μL of herbal tea extract were mixed in a 96-well microplate with 6 μL of 5% aqueous sodium nitrite (NaNO
2) solution. Five minutes later, 6 μL of 10% aqueous AlCl
3 were added to the preparation before vortex. Following 1 min incubation, forty microliters of 1 M NaOH were added before absorbance measurement at 510 nm. The total flavonoid content was calculated with respect to the (–)-epicatechin (Sigma, Darmstadt, Germany) calibration curve (calibration range 6.25–300 µM) and the results were expressed as mg (–)-epicatechin equivalent (EE) per g plant powder.
2.4. Determination of Antioxidant Capacity of Plant Extracts
The antioxidant activity of Antirhea borbonica plant extract was investigated by using the oxygen radical absorbance capacity (ORAC) and the 2,2-Diphenyl-1-picrylhydrazyl (DPPH) radical scavenging assays.
The ORAC assay using fluorescein as the fluorescent probe measures the antioxidant capacity of samples to protect fluorescein from 2,2′-azobis [2-methyl-propionamidin] dihydrochloride AAPH-induced oxidative damage. Briefly, 25 µL of polyphenol extract (at different dilution from 1/2 to 1/200 (in phosphate buffer 75 mM, pH 7.4)) were incubated with 150 µL of fluorescein solution (80 nM). The fluorescence kinetic of fluorescein (Sigma, Darmstadt, Germany) was followed for 2 h at 37 °C at an excitation wavelength of 485 nm and an emission wavelength of 530 nm, after adding 25 µL of AAPH (Sigma, Darmstadt, Germany) solution (150 mM) in each well. The results were based on the area under the curve of fluorescence decay over time and compared with the Trolox calibration curve (concentration range: 6–50 μM). Free radical-scavenging activities of polyphenol-rich plant extracts were expressed as mM Trolox (Sigma, Darmstadt, Germany) equivalent.
DPPH protocol is based on that described by Yang et al. with few modifications [
28]. Briefly, 200 µL of 0.3 mM methanolic solution of DDPH were incubated in a 96-well microplate with either 40 µL of herbal tea extract (about 12 mM GAE, 40 µL of 10 mM gallic acid (Sigma, Darmstadt, Germany) (dilution in DMSO) used as standard polyphenol or 40 µL of 10 mM ascorbic acid (Sigma, Darmstadt, Germany) (dilution in H
2O) and 40 µL of 8.5 mM caffeic acid (Sigma, Darmstadt, Germany) as standard antioxidants. After 30 min incubation in the dark at 25 °C, absorbance was measured at 517 nm and free radical-quenching activity of DPPH (percentage) was calculated by using the following formula:
where A
0 is the absorbance of the reaction mixture and A
1 the absorbance of the reaction mixture in the presence of the plant extract.
2.5. BSA and AGE Preparation and Biochemical Characterization
Glycation of bovine serum albumin (Sigma-Aldrich, Darmstadt, Germany) was performed by incubating 15 mL of BSA (20 g/L in PBS) in the presence of 110 µL of methylglyoxal 40% (MGO, final concentration 7.5 mM) and completed with 15 mL phosphate-buffered saline (PBS; 0.2 M, pH 7.4) containing or not Antirhea borbonica (Ab) plant extract (final concentration 360 µM of GAE). The incubation was performed for BSA (wo. MGO/wo. Ab), BSA + Ab (wo. MGO/w. Ab), BSA + MGO (w. MGO/wo. Ab) and BSA + MGO + Ab (w. MGO/w. Ab) during 7 days under sterile condition (filtration with 0.2 µm Millipore membrane) at 37 °C in the dark and under agitation. All the incubations were performed in triplicates. After incubation, methylglyoxal and plant extract in excess were removed by extensive dialysis against PBS. Protein content of albumin preparations were checked by using bicinchoninic acid assay (BCA) and albumin samples were analyzed by native PAGE (12.5% of acrylamide) and denaturing PAGE (12.5% of acrylamide) and stained with Coomassie blue. Then, samples were stored at –80 °C for further analysis. The antiglycation and antioxidant potential of herbal extract was determined by assessing the level of fructosamines, fluorescent AGE, free amino groups, free sulphydryl groups, tryptophan quenching and β-amyloid aggregation.
2.6. Fructosamine and Fluorescent AGE Determination
Fructosamine content in our preparations was determined according to a protocol previously described by our group [
9]. Briefly, the reaction consists of the reduction of nitroblue tetrazolium (NBT) and the consequent change in absorbance is measured at 530 nm. Results were expressed in nanomol of 1-deoxy-1-morpholinofructose (DMF) according to a calibration curve using a synthetic ketoamine as a standard.
The fluorescence emission intensity of AGE product was obtained with 370 nm excitation wavelength using a spectrophotometer (FluoroMax-4, Horiba, Kyoto, Japan) with excitation and emission slits set at 5 nm [
29]. Samples are diluted at 1.5 mg/mL in 50 mM sodium phosphate buffered saline (PBS, pH 7.4). Percentage of AGE formation was determined using the following formula:
where AGE% represents the relative percent of AGE, ImaxG is maximal fluorescence intensity of glycated albumin and ImaxG0 is the maximal fluorescence intensity of nonglycated albumin (BSA).
2.7. Free Primary Amino and Sulphydryl Group Determination
Quantification of primary free amino groups in our albumin preparation used 2,4,6-trinitrobenzene sulfonate (TNBS) reagent which forms color when complexing with free amino groups [
9]. A calibration curve was established by using increasing concentrations of L-glycine up to 2 mM (Sigma-Aldrich, Darmstadt, Germany). Absorbance was read at 420 nm and results were expressed as mol of amine/mol of protein.
Ellman’s assay containing 5,5-dithiobis(2-nitrobenzoic acid (DTNB) was used to determine thiol group number in albumin preparations. Protocol was well described in a previous study [
9]. Briefly and for each assay, a calibration curve was established by using increasing concentrations of L-cysteine up to 100 nmol (Sigma-Aldrich, Darmstadt, Germany). Thiol group numbers for each albumin preparation was determined in duplicate on two different sample dilutions after absorbance measurement at 412 nm. Results were expressed as the number of free -SH group per mol of BSA.
2.8. Quenching of Intrinsic and Thioflavin T Fluorescence Determination
The intrinsic fluorescence of albumin is mainly attributed to both tryptophan residues present in BSA molecule. The maximum emission of intrinsic fluorescence was determined for our albumin samples (1.5 mg/mL in PBS) from fluorescent emission spectra obtained with Horiba FluoroMax-4 spectrophotometer (250–500 nm range) under excitation wavelength at 270 nm (slit, 5 nm) [
30]. The relative percent of quenching of tryptophan fluorescence was calculated using the following formula:
where Imax
Trp is maximal fluorescence intensity of albumin sample and Imax
Trp0 is the maximal fluorescence intensity of native non-glycated albumin (BSA).
For β-aggregation determination, thioflavin T (ThT) a specific fluorescent probe for amyloid cross β structure was used [
29]. Albumin samples (2.5 µM) were incubated with 30 µM thioflavin T solution (dilution in H
2O) for 1 h at room temperature. The thioflavin emission spectra were obtained in the range of 250–600 nm under excitation at 435 nm (slit, 5 nm). The relative percent of β-amyloid formation was calculated using the following formula:
where Imax
ThT is maximal thioflavin T fluorescence intensity of albumin samples and Imax
ThT0 is the maximal thioflavin fluorescence intensity of nonglycated albumin (BSA).
All fluorescence spectra were corrected for the respective different absorption.
2.9. Erythrocyte Preparations
Erythrocytes were obtained from the French blood national agency (EFS-LR agreement number # 2018001378). Concentrated red blood cells were washed 3 times with sterile isotonic solution (NaCl 0.15 M, pH 7) and prepared at 20% hematocrit in phosphate buffered saline solution/5 mM glucose (PBS/0.1% glucose) in the presence or absence of Ab plant extract (70 µM GAE) and in the absence or presence of 7.5 mM of methylglyoxal solution. After 24 h of incubation at 37 °C, erythrocytes were washed 3 times with NaCl 0.15 M before a direct use by FACS or ektacytometry.
2.10. Zebrafish Maintenance and Treatment
Adult AB wild type zebrafish (3 to 6 months) were housed in the zebrafish facility of the CYROI/DéTROI (La Réunion) and maintained under standard conditions of temperature (28.5 °C), photoperiod (14 h dark/10 h light), pH (7.4) and conductivity (400 μS). Zebrafish were fed daily with commercially available dry (GEMMA 300, Planktovie). All animal experiments were done in CYROI (UMR 1188) and conducted in accordance with the French and European Community Guidelines for the Use of Animals in Research (86/609/EEC and 2010/63/EU) and approved by the local Ethics Committee for animal experimentation of CYROI and the French Government (APAFIS # 2019110510533837_v5).
Zebrafish were divided into three groups—(1) control fish, (2) MGO-treated fish and (3) MGO-treated fish +
Ab. Prior to MGO injection, the third group was treated for 48 h with
Antirhea borbonica. The herbal tea treatment was renewed every 24 h. The two other groups were placed in normal fish water for the 48 h. After 48 h, the first group of fish was injected with 1X PBS (Control fish), the second one with 2.5 µL of MGO (100 mg/kg of body weight) and the third one was injected with 2.5 µL of MGO (100 mg/kg of body weight) and treated one more day with
Ab in order to investigate the effects of
Antirhea borbonica on red blood cells glycation (See
Figure S1).
Twenty-four hours post-injection, the fish were euthanized using tricaine (MS 222-Sigma). Fish were gently dried with a tissue and one eye was removed allowing the ocular cavity to fill with blood. The blood from 2 fish was collected into 50 µL of 1X PBS containing EDTA in order to avoid blood clotting. For each independent experiment, three pools of blood collected from 2 fish were performed for each condition (Control, MGO, MGO + Ab) and three independent experiments were performed.
2.11. Mass Spectroscopy for Hemoglobin Glycation Level Determination
Glycation of both α and β hemoglobin subunits was analyzed by matrix-assisted desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) for mass shift determination as previously described [
31]. Mass spectra were obtained in three independent experiments. On each spectrum, the four main peaks for α and β hemoglobin subunits and their glycated forms were identified. For each peak, the mass (
m/z) and Δ
mass between non-glycated and glycated subunits were obtained. Relative intensity of each peak was calculated as follow—relative % glycation = (intensity glycated-hemoglobin/intensity hemoglobin) × 100.
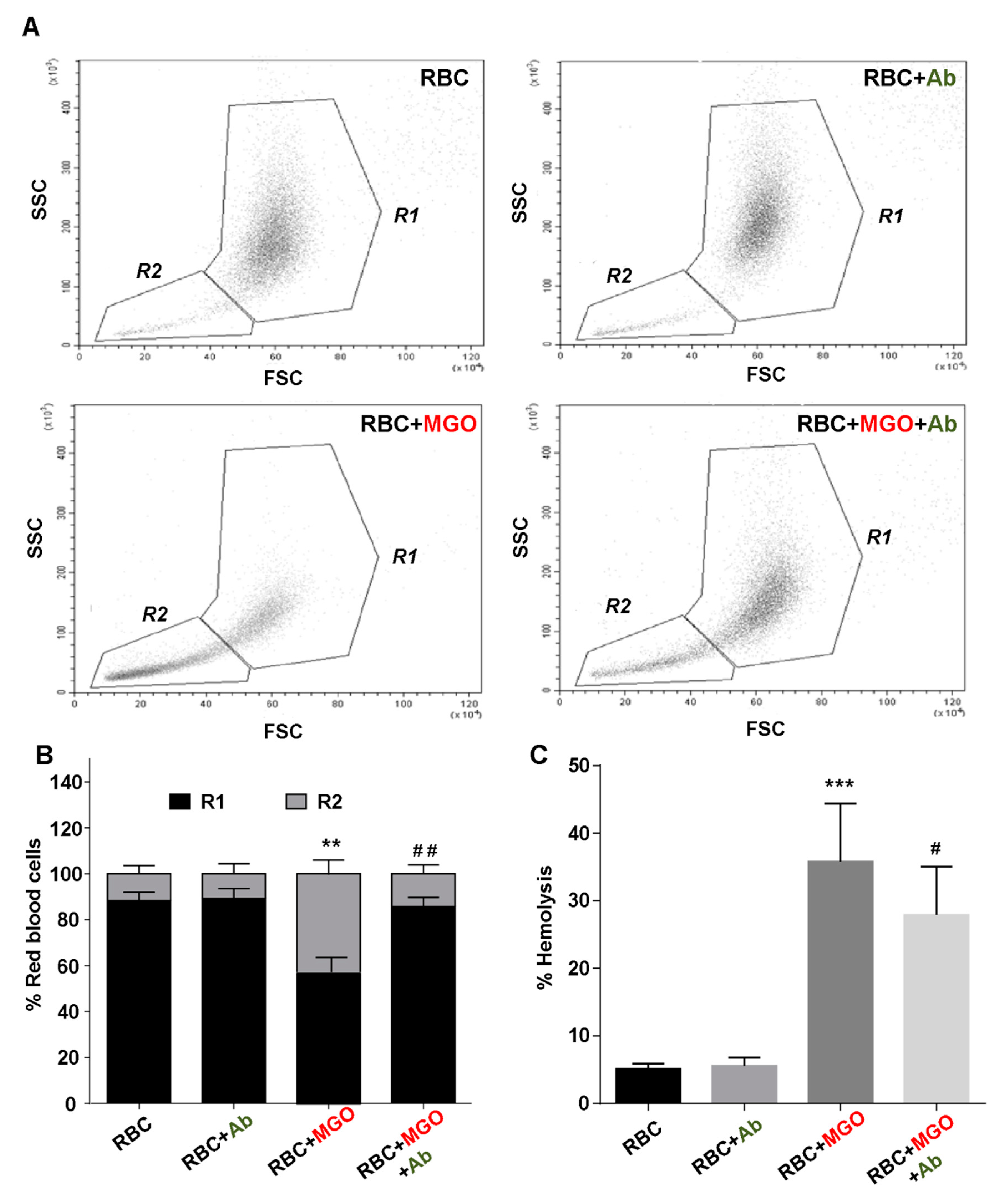
2.12. Erythrocyte Sensitivity to Hemolysis Determination
After 24 h incubation, erythrocyte preparations were washed with NaCl 0.15 M. After centrifugation at 2000× g for 10 min, absorbance of supernatant was measured at 450 nm on a microplate reader (FLUOstar Optima, BMG Labtech, Ortenberg, Germany). The percentage of free hemoglobin resulting from lysis of the cells was calculated in percentage to total hemolysis measured on 10% hematocrit erythrocyte preparations incubated with deionized water.
2.13. Flow Cytometry Assays
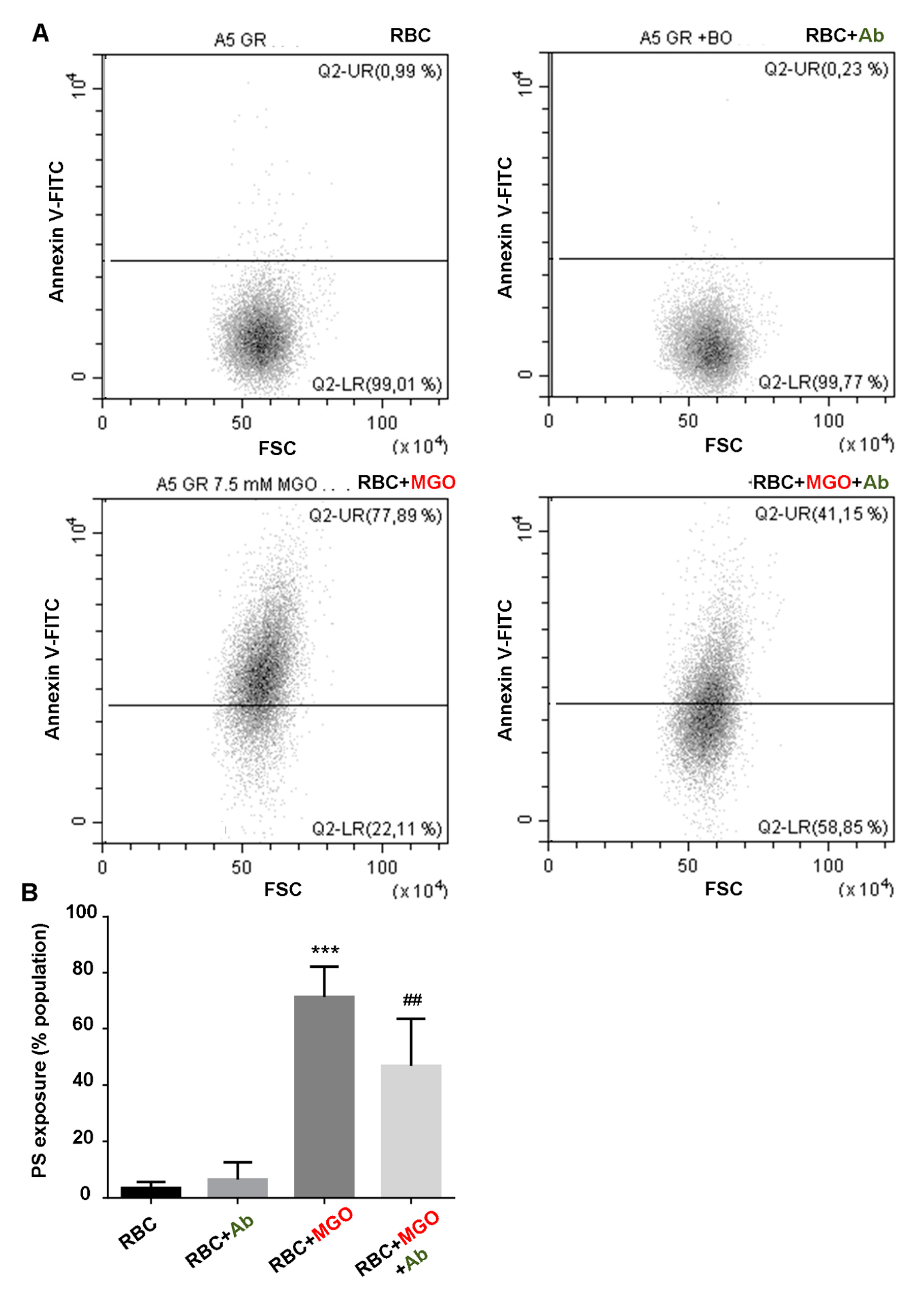
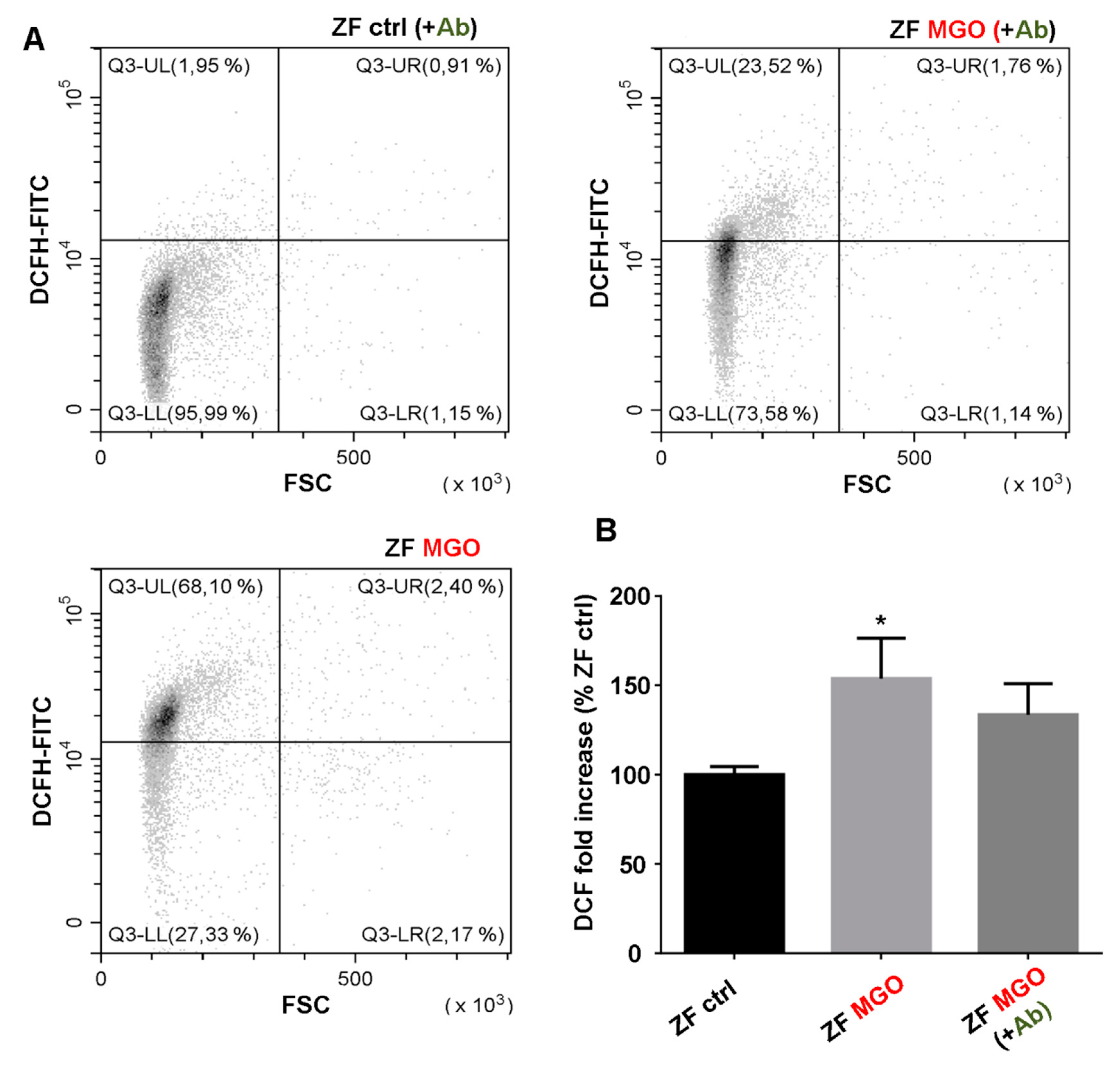
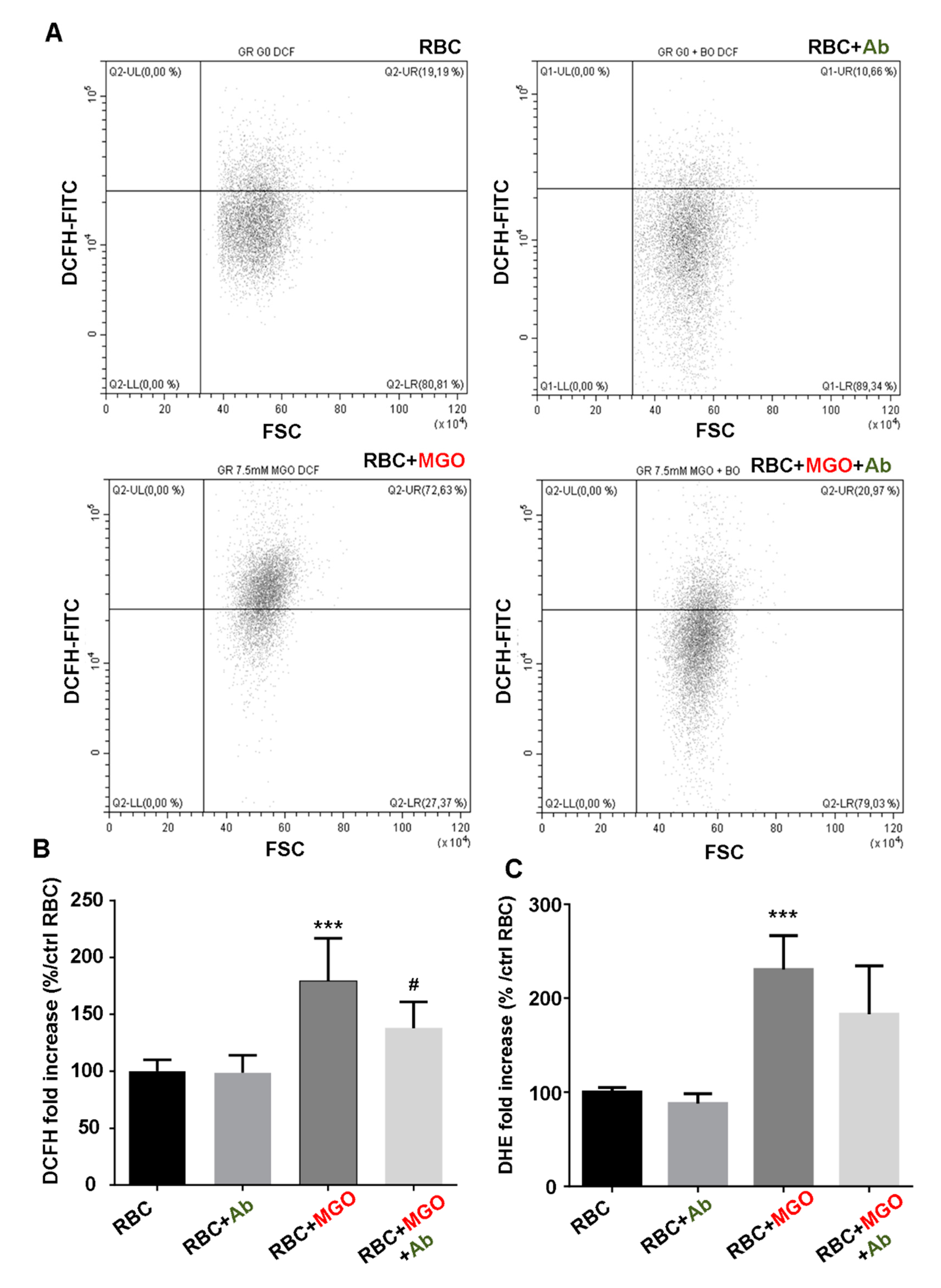
Erythrocyte shape, eryptosis and intracellular reactive oxygen species (ROS) production in our different erythrocyte preparations (from human and zebrafish) were determined by flow cytometry using Beckman Coulter’s CytoFLEX and Cytexpert software (v2.1, Beckman Coulter, Brea, CA, United States). A specific erythrocyte cell population was selected by gating and could be characterized by its typical location in a forward scatter (FSC) versus a side scatter (SSC) parameter graph. For phosphatidylserine exposure determination, 100 µL of erythrocytes (1/50 dilution) were preliminary incubated with 2 μg/mL Annexin V-FITC in binding buffer (BioLegend, San Diego, CA, United States) for 30 min at RT before flow cytometry analysis. Annexin V protein exhibits a high affinity for phosphatidylserine (PS) and was measured with an excitation wavelength of 488 nm and an emission wavelength of 530 nm. For the evaluation of ROS production, 100 µL of erythrocytes (1/50 dilution) were incubated with 2 µM of fluorescent probes, dichlorodihydrofluoresceindiacetate (DCFH-DA; Sigma-Aldrich, Darmstadt, Germany) or dihydroethidium (DHE; Sigma-Aldrich, Darmstadt, Germany) for 30 min at RT.
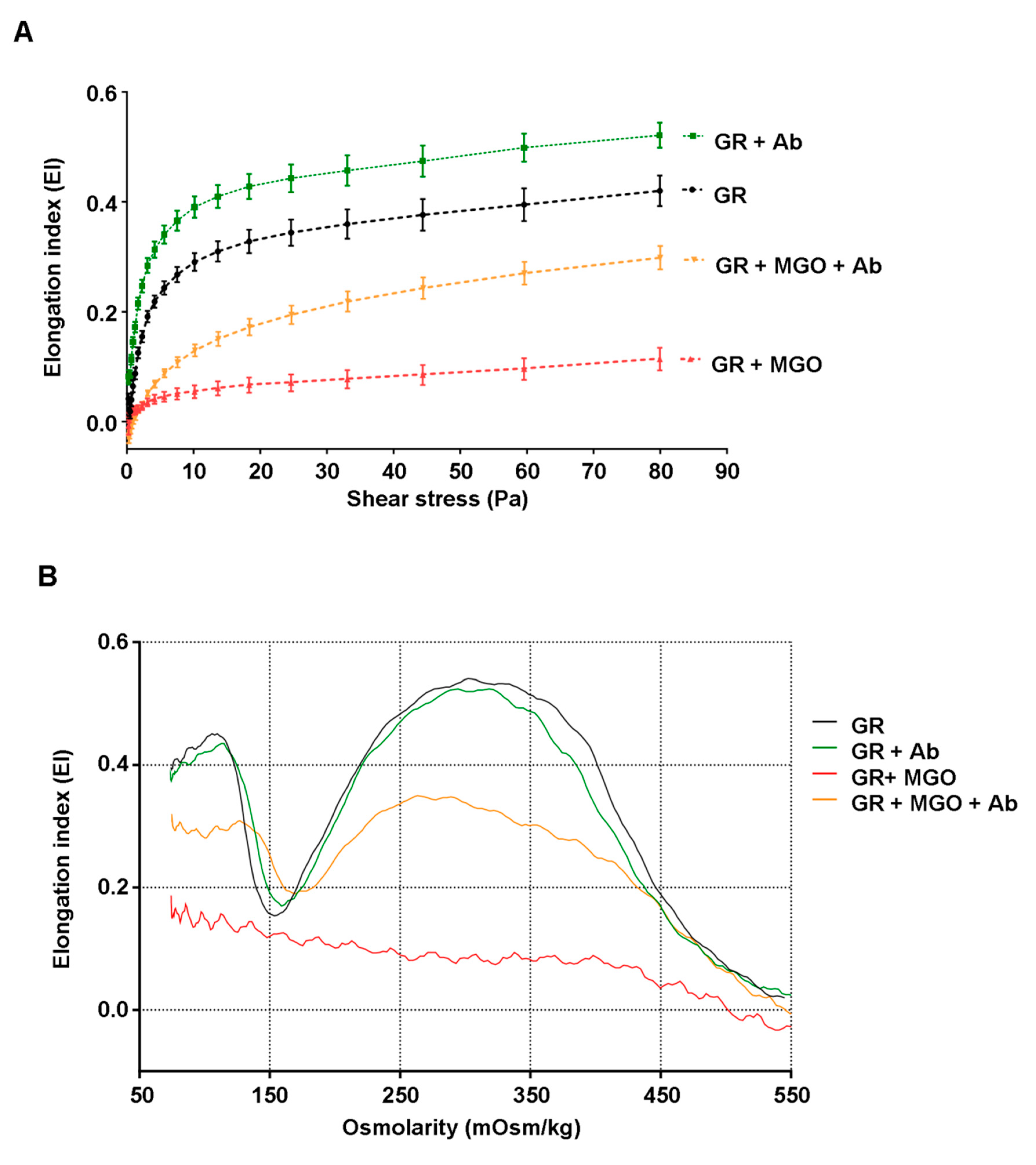
2.14. Shear Stress Gradient Ektacytometry
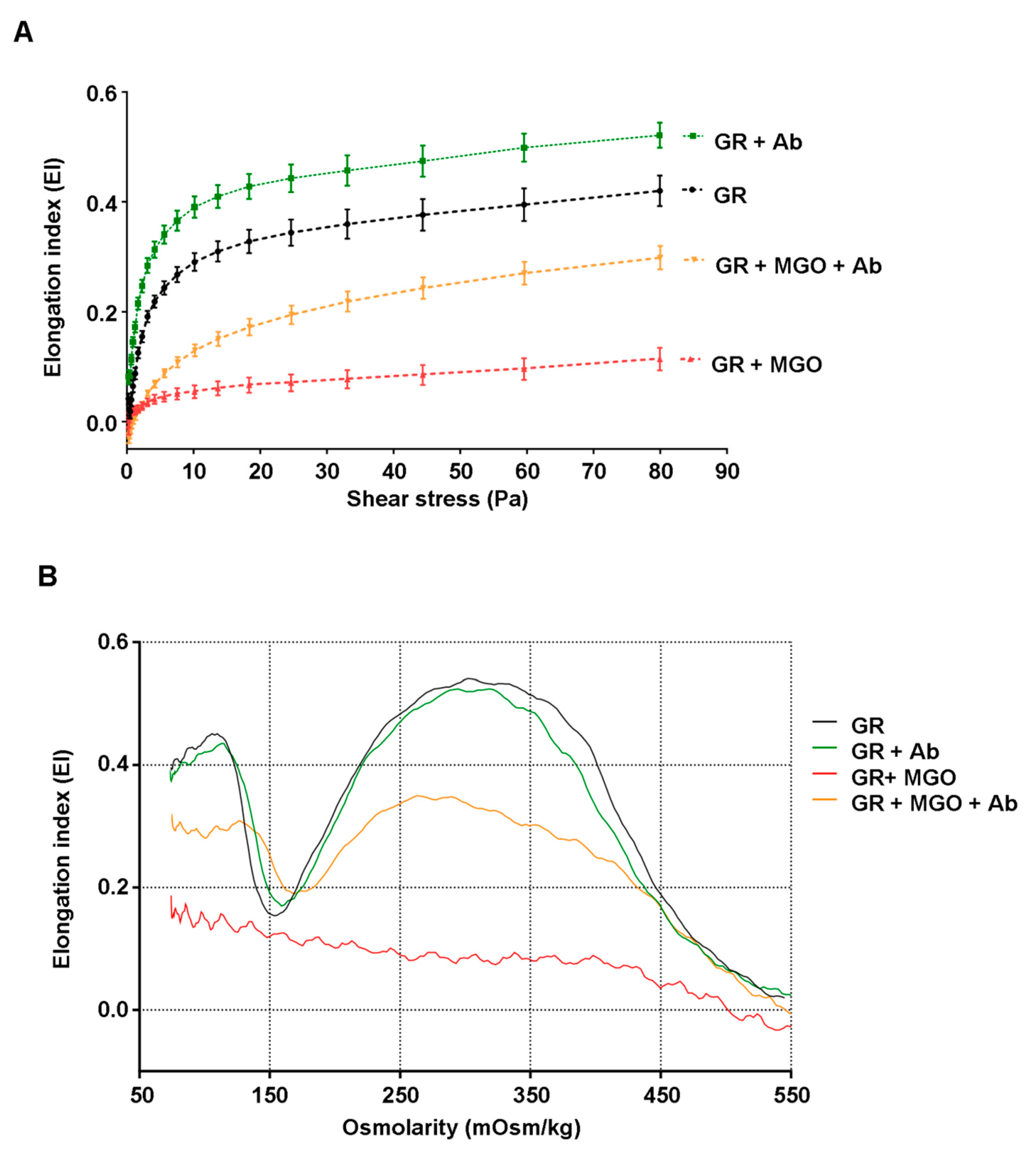
The determination of erythrocytes membrane deformability was performed using an ektacytometer (LORCCA MaxSis, Mechatronics, Zwaag, The Netherlands), which measures the elongation of red blood cells as a function of an increasing shear stress (elongation curve) or osmotic gradient (Osmoscan curve).
To obtain elongation curves, red blood cells suspended at 20% hematocrit were diluted 200 times in an iso-osmolar solution of polyvinylpyrrolidone buffer (PVP, viscosity 28.6 mPa/s). Deformation was expressed as an elongation index (EI) calculated for 19 shear-stress intensities between 0.30 and 80 Pa (increasing rotation speed) as follow—EI = (A – B)/(A + B). In this formula, A and B represent the length and the width of the ellipsoid diffraction pattern, respectively. The deformability curve obtained by plotting the calculated values for EI versus the shear-stress (SS) [
32] was analyzed using the Lineweaver-Burke model which links the shear-stress and the elongation index according to the following equation:
From this equation, the maximum elongation index (EI
max) and the shear stress at half maximal deformation (SS
1/2) can be calculated. Both parameters appeared to be relevant indicators of erythrocyte deformability capacity [
32].
2.15. Osmotic Gradient Ektacytometry
For the acquisition of the osmoscan curve, 200 µL of red blood cells suspended at 20% hematocrit were suspended in 5 mL PVP buffer. A constant shear stress (30 Pa) was applied during the EI measurement with a gradual increase in osmolality from 80 to 500 mOsmol/kg. The resulting osmotic gradient curves reflect red blood cells deformability as a continuous function of suspending medium osmolality. Different parameters obtained from these curves were analyzed, such as the minimal elongation index value measured at low-osmotic environment (Ei
os-min) and maximum elongation index values corresponding to the maximal deformability obtained near the isotonic osmolality (Ei
os-max). From these parameters, the ratio of maximal and minimal EI values was calculated—rEI = Ei
os-max/Ei
os-min. This ratio value can be interpreted as the amplitude of deformability of erythrocytes [
33].
2.16. Statistical Analysis
Data were expressed as the mean ± standard error of the mean (SEM) from at least three independent experiments performed in triplicate. Statistical analyses were performed with Prism (GraphPad Software Inc., San Diego, CA, USA). Statistical significance was determined using one-way ANOVA followed by Dunnett’s test, with a p-value < 0.05 required for significance.
4. Discussion
Glycoxidation is a physiological complex process which is amplified in hyperglycemic conditions. The glycation reaction corresponds to the initial precursor of this process to other even more harmful phenomena such as oxidative stress, inflammation and platelet aggregation. The AGEs accumulation, resulting from this critical process, is involved in progression of diabetes complication such as vascular complications [
38].
Albumin and erythrocytes represent abundant circulating components which render them very sensitive to glycoxidative modifications [
6]. Albumin and erythrocytes in their native form exert many beneficial properties but when glycoxidized they become deleterious actors that were involved in the development of vascular complications. Indeed, serum albumin, primary glycoxidation target, constitutes an important provider of circulating AGEs and therefore of increased plasmatic oxidative stress. The resulting oxidative damage thus affects the main molecular and cellular actors of atherogenesis, such low-density-lipoprotein (LDL), the main contributor of atheroma plaque development and the weakened endothelium that will facilitate macrophage and lipid infiltration [
39,
40]. Erythrocyte is now considered as a new culprit contributing to the pathogenesis of atherosclerosis by directly participating in intraplaque hemorrhage and/or thrombus formation [
41]. Consequently, inhibition of glycation, oxidation and glycoxidation appears to be the basis of antiatherogenic and antithrombotic strategies to be considered and the discovery and investigation of AGEs inhibitors would offer a potential therapeutic approach for prevention of diabetic complications. Some potential synthetic AGE inhibitors and antiglycant agents were discovered and tested such as metformin or aminoguanidine [
42]. But, because of potential toxic and side effects of these synthetic molecules, antioxidant compounds from natural products such as polyphenols were preferred as safe and promising agents for their antiglycation activities and for prevention of AGEs formation.
It appeared that our
Ab aqueous plant extract exhibited a higher polyphenols and flavonoids content (5.2% GAE (
w/w) /2.1% EE (
w/w)) than the acetonic extract of the same plant investigated by Marimoutou et al. [
25]. In addition, major phenolic acids including caffeic acids derivatives and flavonoids like quercetin derivatives known to be abundant in medicinal plants were identified in our plant extract [
43]. The two major compounds of this extract are caffeic acids derivatives—dicaffeoylquinic acid and chlorogenic acid. The antioxidant activity and ROS trapping capacity of dicaffeoylquinic acid has been reported in the treatment of oxidative damage induced by ROS such as age-related diseases and certain degenerative diseases [
44].
Chlorogenic acid delays the intestinal absorption of glucose and therefore its passage through the bloodstream [
45]. This phenolic acid also protects LDL from oxidation and thus limits the formation of atheroma plaque, a major vascular complication associated with diabetes [
46].
This high polyphenol level could explain the strong free radical-scavenging activity of the plant extracts shown with DPPH and ORAC assays. Indeed, polyphenols were identified as down regulators of ROS production and pro-inflammatory cytokines from H
2O
2 exposed preadipocyte cells and LPS exposed adipocyte [
24,
25]. This beneficial extract prepared in the form of herbal tea and used as therapeutic human diet can also act on the main circulating cells and plasmatic protein. Indeed, many studies showed that several polyphenols including caffeic acid or quercetin derivatives can be absorbed, metabolized after a specific human diet and be detected in plasma the first few hours after ingestion [
47,
48].
In our study, aqueous extract of
Ab significantly reduced albumin glycation at several stages of this deleterious process. Indeed, the inhibitory effects of this extract acts in the early glycation products (ketoamines) and also in the advanced products (AGEs) formation. Effect of
Antirhea borbonica was not limited to glycation process inhibition. Indeed, β-structure aggregates formation was found not only with MGO-glycation but also with
Ab extract. This conformational change of albumin into β-sheet structure induced by the plant suggests a direct impact of
Ab on the tertiary structure of the protein. This hypothesis is reinforced by the results of tryptophan fluorescence showing an important tryptophan fluorescence quenching of albumin in the presence of the plant. As attested by tryptophan fluorescence quenching phenomenon, a partial unfolding occurs in the presence of MGO or
Antirhea borbonica, similarly to what was already observed in case of aggregation process of native BSA [
49]. In our experimental conditions, this change of tertiary structure could be associated with increasing exposure of hydrophobic pockets usually detected with ANSA fluorescent probe [
49]. This increased accessibility of the hydrophobic regions induced by the α-dicarbonyl agent or the plant appears to be favorable for the aggregation process.
These last results showing the direct impact of the rich polyphenol plant extract on albumin structure are consistent with numerous studies showing the affinity of certain polyphenols for albumin, particularly dicaffeoylquinic acid and chlorogenic acid [
50,
51]. Indeed, numerous polyphenols including phenolic acids or flavonoids are known to exhibit high affinities to serum albumin [
52]. In the present study, caffeic acid induced same effects on albumin as
Antirhea borbonica in terms of tryptophan fluorescence quenching and β-structure aggregates formation (
Figure S8). These latest data raise the question of the potential impact of Ab herbal tea on the consequences of such an increase in albumin aggregation.
Several hypotheses can be put forward to explain the inhibition action of
Ab extract on MGO-induced glycation of albumin. First, polyphenols could directly react with methylglyoxal and then neutralizing its glycating and oxidizing capacity. Indeed, many phenolic acids were found to be able to bind and reduce methylglyoxal molecules [
53]. Second, polyphenols and the glycative agent may compete for the sensitive sites of the protein. Indeed, a very recent study by Tagliazucchi et al. showed that some phenolic acids could effectively bind albumin at key glycation sites [
54].
It is clear that glycation impairs the protective antioxidant properties of albumin [
55]. By limiting glycation and thus AGEs formation, the
Ab extract may protect albumin antioxidant properties. It would be interesting to test the protective effect of
Ab extract against glycated albumin mediated toxicity to erythrocytes [
56].
The observed beneficial effects of
Antirhea borbonica extract to protect albumin from glycation are comforted by the results highlighting its capacity to prevent red blood cells from oxidative injury. Indeed, MGO-induced oxidative stress damages in erythrocytes represent a determinant starting point that initiate membrane fragility and eryptotic process of red blood cells. This membrane fragility results in increased oxidative mediated hemolysis induced by the α-dicarbonyl agent. We evidenced an effective protection of
Ab for reducing ROS production, preserving membrane integrity and function and slowing down the senescence process. An enzymatic defense system in erythrocytes could be involved in the protective effect of
Ab. Indeed, alteration of this protective mechanism in ROS-damaged erythrocytes were already evidenced in H
2O
2 or glucose-exposed red blood cells [
57]. In addition, several studies report that polyphenols, such as Silymarin and polyphenol-rich extracts from
Achillea species, could significantly stimulate activities of the antioxidant enzymatic system such as catalase or peroxidase [
58,
59]. Thus, the return to redox homeostasis with phenolic compounds contributes to reduce lipid peroxidation and preserve membrane integrity [
60].
Not only
Ab extract prevent erythrocytes from oxidative stress and eryptosis and also protect their membrane integrity and hence preserving erythrocyte morphology and deformability capacity. Deformation capacity constitutes a determinant rheological parameter for erythrocytes, which have to pass through tiny capillaries. Then, a relative rigidity of the RBC membrane may provoke significant disruption in the microcirculation efficiency and thus represents a putative contributor to microvasculature occlusion [
61].
Here, and for the first time, methylglyoxal as a glycoxidizing agent, was shown to induce severe erythrocyte membrane stiffness in terms of cell deformability capacity following a change in shear stress and also when osmolarity deviates from physiological conditions. In our experimental conditions, if MGO-glycated erythrocytes totally lost their ability to be deformed,
Ab extract appears to be able to significantly preserve this capacity. RBC membrane disorders causing this loss of deformability may be due to the alteration of specific cytoskeleton proteins such spectrin, ankyrin or protein band.3 [
62]. These proteins ensure cohesion between the membrane lipid bilayer and cytoskeleton, thus providing structural and functional integrity of RBC. In oxidative stress situation induced by H
2O
2 or AAPH, α- and β-spectrin and band.3 proteins in RBC were shown to be particularly prone to oxidative damages [
63,
64]. Paiva-Martins et al. showed that many olive oil polyphenolic compounds had the capacity to protect RBC from oxidative injury and suggested that this protection could be mediated by the interaction of 3,4-DHPEA-EDA and hydroxytyrosol with RBC membrane proteins, improving the stability of erythrocytes [
64,
65]. In our study, methylglyoxal may primarily affect the key proteins, ankyrin, spectrin and band.3, tightly responsible for erythrocyte flexibility and stability.
The direct consequence of membrane flexibility protection exerted by the
Ab extract is an increased hemolysis resistance and an improvement in the deleterious morphological change of RBC. Although
Ab extracts exert potent cytoprotective effects, it does not seem to impact hemoglobin, the main erythrocyte protein. Indeed, by mass spectrometry, we can observe a significant increase in hemoglobin glycation in the presence of methylglyoxal suggesting that the α-dicarbonyl reagent can easily penetrate the membrane and also cause intracellular damage. However, in our experimental conditions,
Ab extract does not seem to be able to counteract the glycoxidant action of MGO. This conversely action exerted by
Antirhea borbonica on the membrane versus the intracellular level supports the hypothesis raised above concerning the absence of a direct neutralizing action of the plant on the glycoxidizing agent. The results obtained on the zebrafish model treated with
Antirhea borbonica support this hypothesis. According to several studies, the hyperglycemia model of zebrafish treated with methylglyoxal appears to be attractive for understanding underlying mechanisms associated with diabetic complications such as microvascular complications [
66]. Until now, methylglyoxal in zebrafish was rather tested in the embryonic or larval stage than in adult fish [
67]. In our study, a 24 h treatment with MGO through intraperitoneal injection was sufficient to cause intracellular oxidative stress in zebrafish erythrocytes without inducing eryptosis. Furthermore, a 48 h preliminary bathing treatment with
Ab seems to protect red blood cells against MGO induced oxidative stress. These preliminary results on zebrafish blood seem very promising and further ongoing projects are highly warranted and under progress to decipher the beneficial effect of
Antirhea borbonica on a diet induced obese zebrafish model. In summary, in the present work, we showed not only that
Antirhea borbonica extract preserve in vitro erythrocyte from MGO-induced damages and also acts as an in vivo protecting agent in a zebrafish model recently developed in our group [
36,
68]. If these results cannot be extrapolated to what could be encountered in pathological conditions, several points may in some way relate our in vitro experimental conditions to in vivo situations. Actually, if albumin and erythrocyte glycation was performed by using supraphysiological concentration of the glycating agent, such concentration was chosen to operate a rapid glycation model for long half-life proteins, 21 days and 120 days for albumin and hemoglobin, respectively. These vitro glycation models appeared relevant to what could be encountered in vivo as most of oxidative damages, identified in our in vitro glycated albumin and erythrocytes preparations, were retrieved in albumin and erythrocytes isolated from diabetics [
10,
15,
69].
In addition, the protective effect of Ab was observed in vivo in zebrafish which water bath comprised 0.5 g of plant extract in 2 L. In term of comparison, when used as an herbal tea, 4 g of the plant tea leaves are used for infusion into 1 L [
23].

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}