Microorganisms and Biostimulants Impact on the Antioxidant Activity of Buckwheat (Fagopyrum esculentum Moench) Sprouts

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Experiment Set-Up, and Sample Preparation
2.2. Extracts Preparation
2.3. Determination of Antioxidant Activity
2.3.1. Ferric Reducing Antioxidant Power (FRAP) Assay
2.3.2. 1,1-Diphenyl-2-Picrylhydrazyl (DPPH) Assay
2.3.3. Photochemiluminescence (PCL) Assay
2.3.4. Determination of Total Polyphenols (TP) Content
2.3.5. Determination of Organic Acids Content
2.3.6. Determination of Rutoside Content
2.4. Statistical Analyses
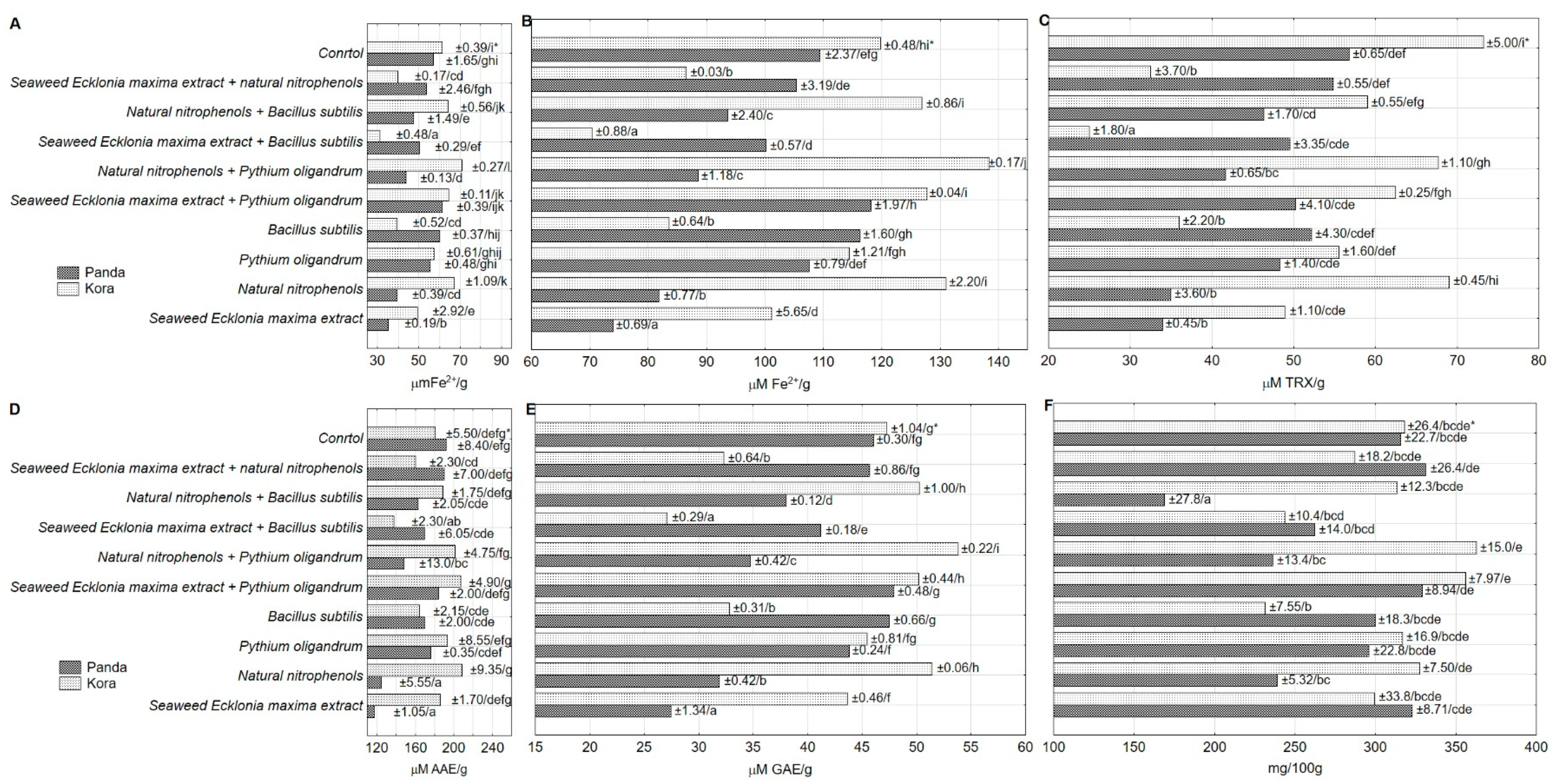
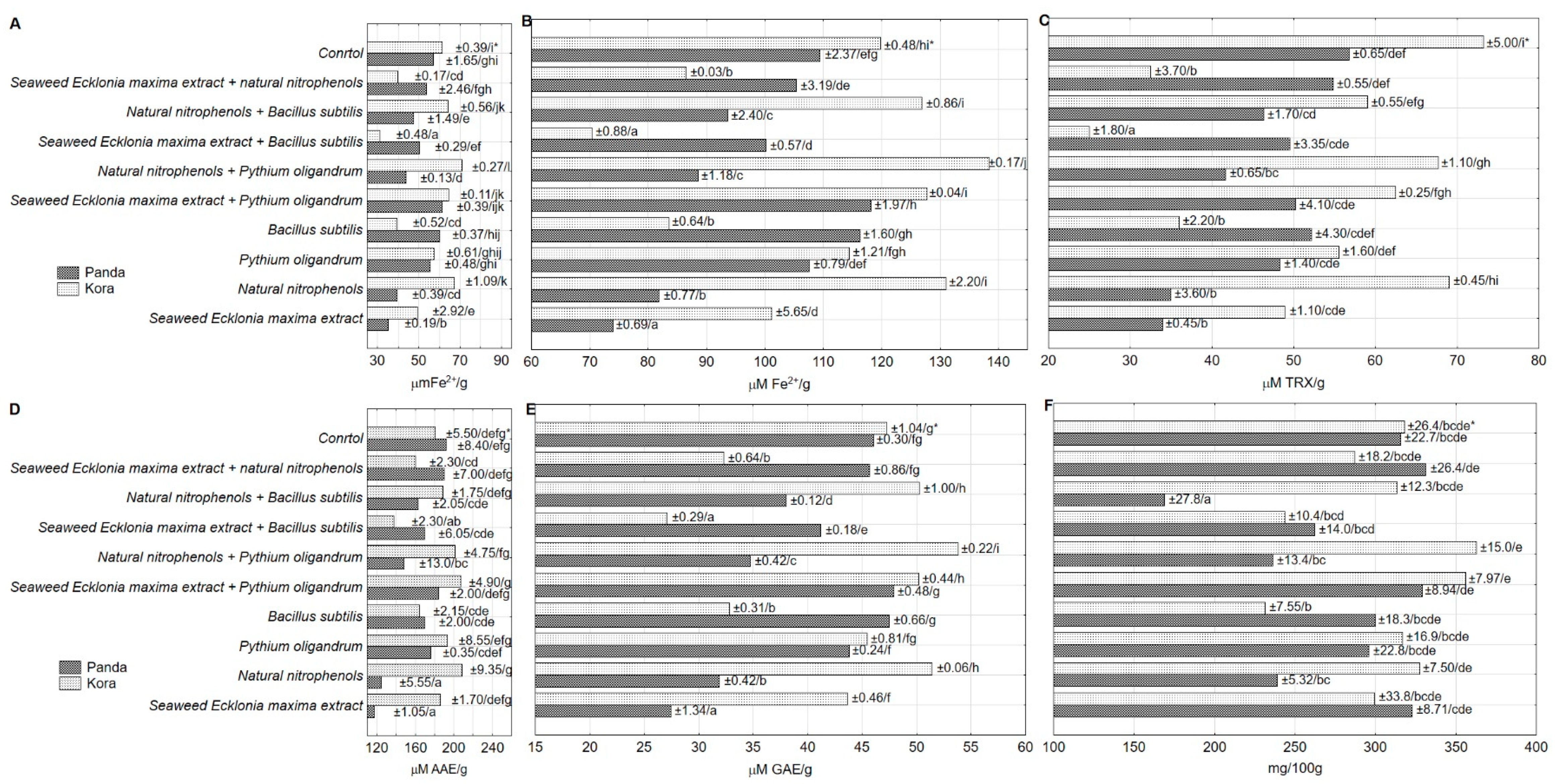
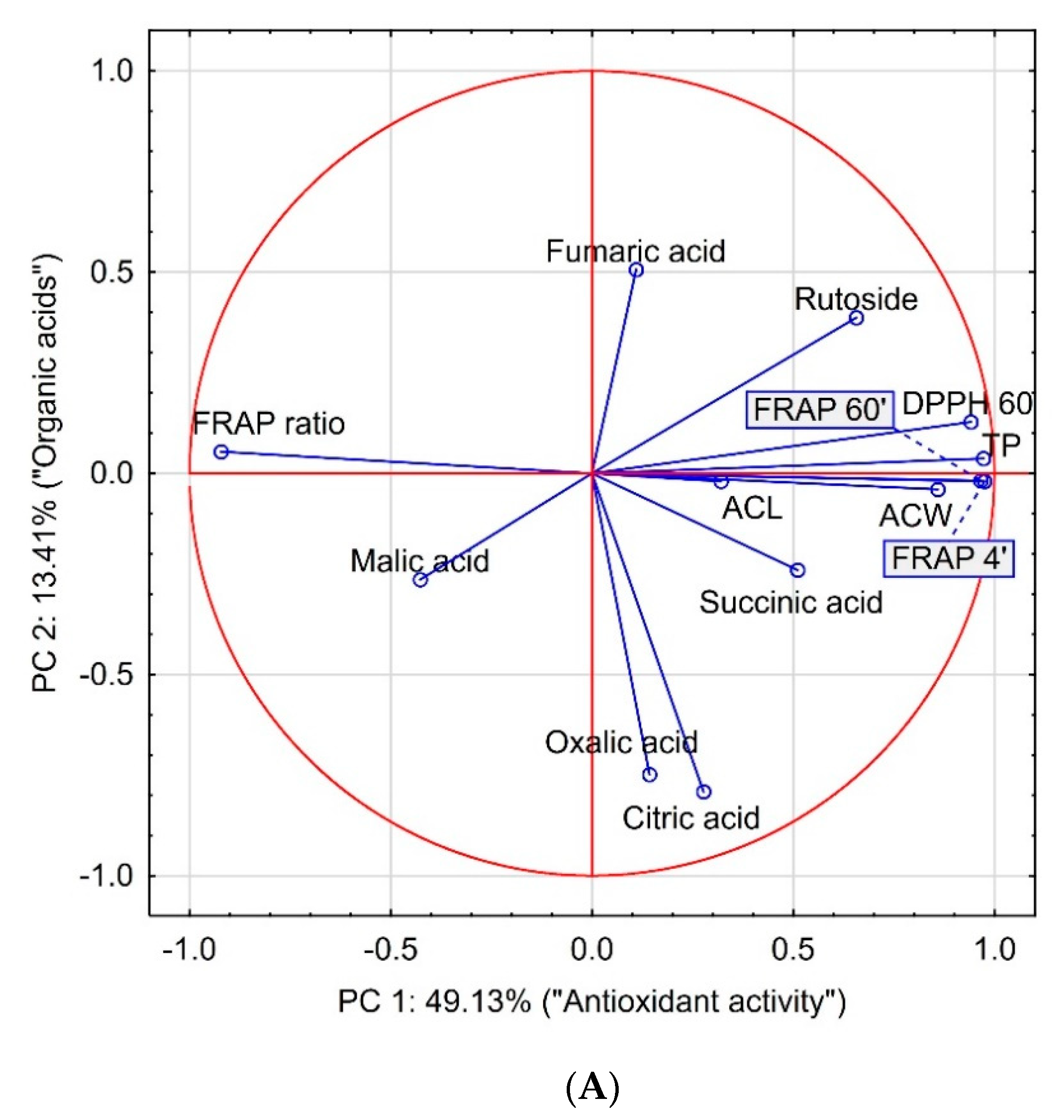
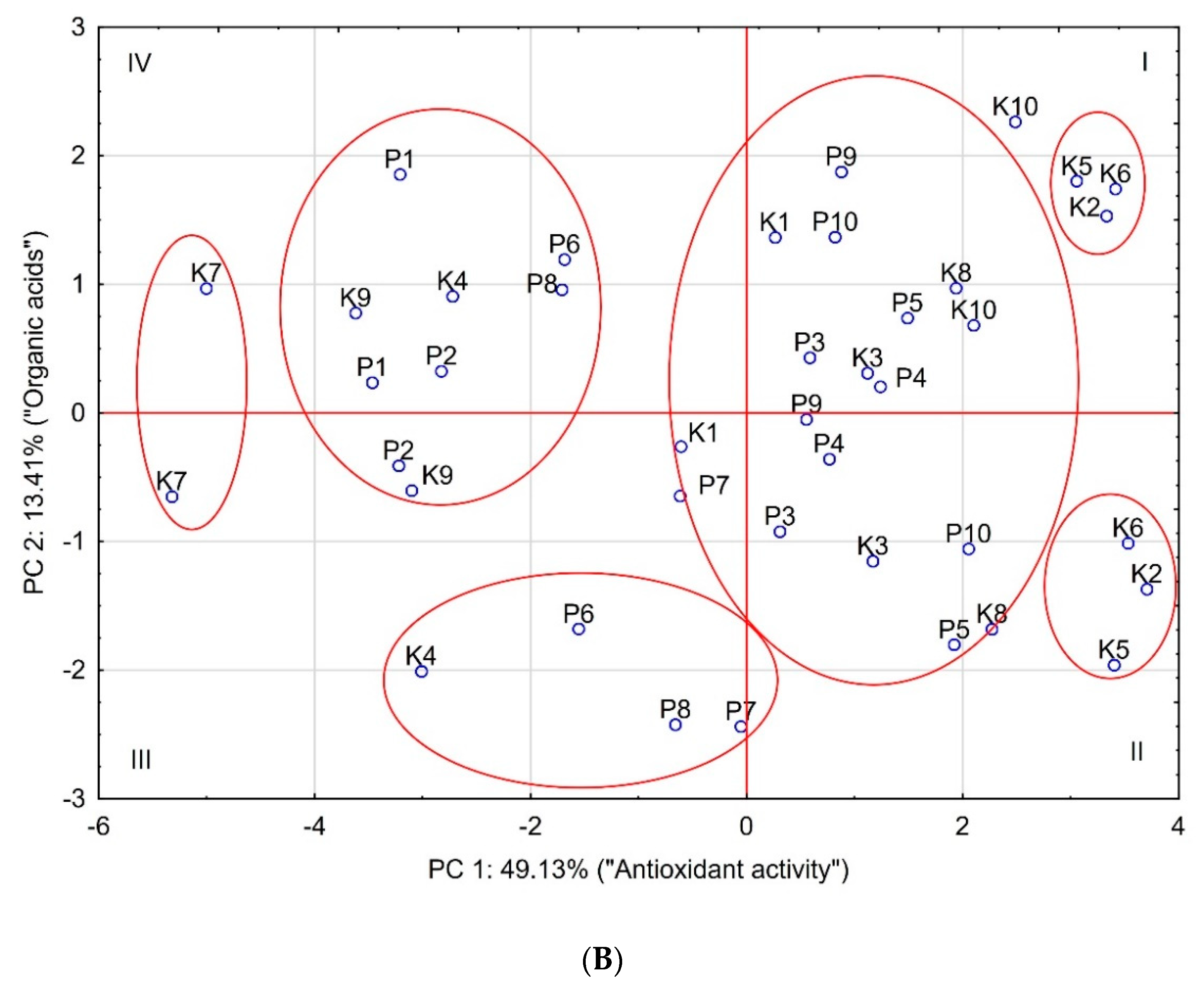
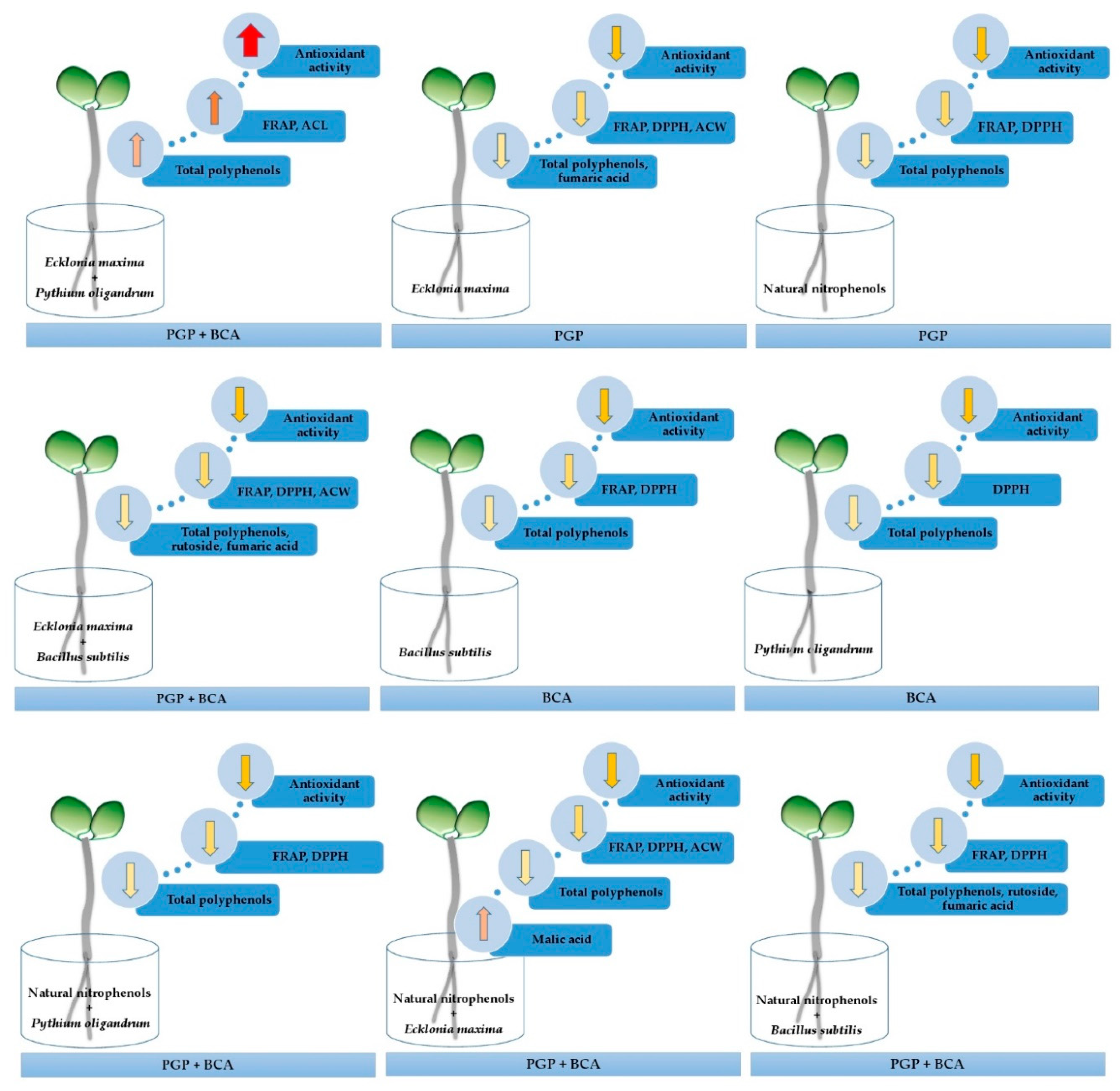
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gimenez-Batista, J.A.; Zieliński, H. Buckwheat as functional food and its effects on health. J. Agric. Food Chem. 2015, 63, 7896–7913. [Google Scholar] [CrossRef] [PubMed]
- World Health Statistic 2018: Monitoring Health for the SDGs, Sustainable Development Goals; World Health Organization: Geneva, Switzerland, 2018; ISBN 978-92-4-156558-5.
- Benincasa, P.; Falcinelli, B.; Lutts, S.; Stagnari, F.; Galieni, A. Sprouted grains: A comprehensive revive. Nutrients 2019, 11, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomé-Sánchez, I.; Martín-Diana, A.B.; Peñas, E.; Bautista-Expósito, S.; Frias, J.; Rico, D.; González-Maillo, L.; Martinez-Villaluenga, C. Soluble phenolic composition tailored by germination conditions accompany antioxidant and anti-inflammatory properties of wheat. Antioxidants 2020, 9, 426. [Google Scholar] [CrossRef] [PubMed]
- Jing, R.; Li, H.Q.; Hu, C.L.; Jiang, Y.P.; Qin, L.P.; Zheng, C.J. Phytochemical and pharmacological profiles of three Fagopyrum buckwheats. Int. J. Mol. Sci. 2016, 17, 589. [Google Scholar] [CrossRef] [Green Version]
- Kim, L.S.; Kim, K.S.; Park, C.H. Introduction and nutritional evaluation of buckwheat sprout as a new vegetable. Food Res. Int. 2004, 37, 319–327. [Google Scholar] [CrossRef]
- Márton, M.; Mándoki, Z.; Csapó-Kiss, Z.; Csapó, J. The role of sprouts in human nutrition. A review. Acta Univ. Sapientiae Aliment. 2010, 3, 81–117. [Google Scholar]
- Ganeshpurkar, A.; Saluja, A.K. The pharmacological potential of rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Meier, F.; Lo, A.J.; Yuan, W.; Sze, V.L.P.; Chung, L.H.; Yuk, G.H. Overview of recent events in the microbiological safety of sprouts and new intervention technologies. Compr. Rev. Food Sci. Food Saf. 2013, 12, 265–280. [Google Scholar] [CrossRef]
- Zou, T.; Xu, N.; Hu, G.; Pang, J.; Hu, H. Biofortification of soybean with zinc and bioaccessibility of zinc in the sprouts. J. Sci. Food Agric. 2014, 94, 3053–3060. [Google Scholar] [CrossRef]
- Liu, H.K.; Kang, Y.F.; Zhao, X.Y.; Liu, Y.P.; Zhang, X.W.; Zhang, S.J. Effect of elicitation on bioactive compounds and biological activities of sprouts. J. Funct. Foods 2019, 53, 136–145. [Google Scholar] [CrossRef]
- Benincasa, P.; D’Amato, R.; Falcinelli, B.; Troni, E.; Fontanella, M.C.; Frusciante, S.; Guiducci, M.; Beone, G.M.; Businelli, D.; Diretto, G. Grain endogenous selenium and moderate salt stress work as synergic elicitors in the enrichment of bioactive compounds in maize sprouts. Agronomy 2020, 10, 735. [Google Scholar] [CrossRef]
- Briatia, X.; Jomduang, S.; Park, C.H.; Lumyong, S.; Kanpiengjai, A.; Khanongnuch, C. Enhancing growth of buckwheat sprouts and mircogreens by endophytic bacterium inoculation. Int. J. Agric. Biol. 2017, 19, 374–380. [Google Scholar] [CrossRef]
- Zhong, L.; Niu, B.; Tang, L.; Chen, F.; Zhao, G.; Zhao, J. Effects of polysaccharide elicitors from endophytic Fusarium oxysporum Fat9 on the growth, flavonoid accumulation and antioxidant property of Fagopytum tataricum sprout cultures. Molecules 2016, 21, 1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Gupta, R.; Pandey, R. Rice seed priming with picomolar rutin enhance rhizospheric Bacillus subtilis CIM colonization and plant growth. PLoS ONE 2016, 11, e01466013. [Google Scholar] [CrossRef]
- Witkowicz, R.; Biel, W.; Chłopicka, J.; Galanty, A.; Gleń-Karolczyk, K.; Skrzypek, E.; Krupa, M. Biostimulants and microorganisms boost the nutritional composition of buckwheat (Fagopyrum esculentum Moench) sprouts. Agronomy 2019, 9, 469. [Google Scholar] [CrossRef] [Green Version]
- Frederiks, C.; Wesseler, J.H.H. A comparison of EU and US regulatory frameworks for the active substance registration of microbial biological control agents pest. Manag. Sci. 2019, 75, 87–103. [Google Scholar] [CrossRef] [Green Version]
- Jardin, P. Plant Biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Przybysz, A.; Gawrońska, H.; Kowalkowski, Ł.; Szalacha, E.; Gawroński, S.W. The biostimulant Asahi SL protects the growth of Arabidopsis thaliana L. plants when cadmium is present. Acta Sci. Pol. Hortorum Cultus. 2016, 15, 37–48. [Google Scholar]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Tava, A. Microalgal biostimulants and biofertilisers in crop production. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant Biostimulants: Importance of the quality and yield of horticultural crops and the improvement of plant tolerance to abiotic stress—A review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Woo, S.L.; Comite, E.; El-Nakhel, C.; Rouphael, Y.; Fusco, G.M.; Borzacchiello, A.; Lanzuise, S.; Vinale, F. Application of Trichoderma harzianum, 6-pentyl-α-pyrone and plant biopolymer formulations modulate plant metabolism and fruit quality of plum tomatoes. Plants 2020, 9, 771. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1996, 28, 25–30. [Google Scholar] [CrossRef]
- Popov, I.; Lewin, G. Antioxidative homeostasis: Characterisation by means of chemiluminescent technique in methods in enzymology. In Oxidants and Antioxidants; Packer, L., Ed.; Academic Press: Cambridge, MA, USA, 1999; pp. 96–100. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticult. 1995, 16, 144–153. [Google Scholar]
- Dobrowolska-Iwanek, J.; Zagrodzki, P.; Prochownik, E.; Jarkiewicz, A.; Paśko, P. Influence of brassica sprouts on short chain fatty acids concentration in stools of rats with thyroid dysfunction. Acta Pol. Pharm. 2019, 76, 1005–1014. [Google Scholar] [CrossRef]
- Paśko, P.; Galanty, A.; Żmudzki, P.; Gdula-Argasińska, J.; Zagrodzki, P. Influence of different light conditions and time of sprouting on harmful and beneficial aspects of rutabaga sprouts in comparison to their roots and seeds. J. Sci. Food Agric. 2019, 99, 302–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tibco. 2020. Available online: https://www.scribd.com/document/321061529/STATISTICA-Electronic-Manual (accessed on 22 June 2020).
- Kim, H.J.; Park, K.J.; Lim, J.H. Metabolomic analysis of phenolic compounds in buckwheat (Fagopyrum esculentum M.) sprouts treated with methyl jasmonate. J. Agric. Food Chem. 2011, 59, 5707–5713. [Google Scholar] [CrossRef] [PubMed]
- Świeca, M. Potentially bioaccessible phenolics, antioxidant activity and nutritional quality of young buckwheat sprouts affected by elicitation and elicitation supported by phenylpropanoid pathway precursor feeding. Food Chem. 2015, 192, 625–632. [Google Scholar] [CrossRef]
- Colonna, E.; Rouphael, Y.; Barberi, G.; De Pascale, S. Nutritional quality of ten leafy vegetables at two light intensitis. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef]
- Chłopicka, J.; Folta, M.; Barton, H.; Sitek, A. Badanie aktywności antyoksydacyjnej oraz polifenoli w aspekcie jakościowym i ilościowym w kiełkach gryki (Antioxidant activity, qualitative and quantitative analysis of polyphenols in buckwheat sprouts). Bromat. Chem. Toksykol. 2014, 47, 336–341. [Google Scholar]
- Kreft, S.; Janes, D.; Kreft, I. The content of fagopyrin and polyphenols in common and tartary buckwheat sprouts. Acta Pharm. 2013, 63, 553–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.J.; Lim, J.H.; Park, K.J.; Kang, S.; Kim, D.S.; Park, S. Methyl jasmolate treated buckwheat sprout powder enhances glucose metabolism by potentiating hepatic insulin signalling in estrogen-deficient rats. Nutrition 2016, 32, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Sytar, O.; Hemmerich, I.; Zivcak, M.; Rauh, C. Comparative analysis of bioactive phenolic compounds composition from 26 medicinal plants. Saudi J. Biol. Sci. 2018, 25, 631–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziadek, K.; Kopeć, A.; Piątkowska, E.; Leszczyńska, T.; Pisulewska, E.; Witkowicz, R.; Bystrowska, B.; Francik, R. Identification of polyphenolic compounds and determination of antioxidant activity in extracts and infusion of buckwheat leaves. Eur. Food Res. Technol. 2018, 244, 333–343. [Google Scholar] [CrossRef]
- Suzuki, T.; Kim, S.J.; Takigawa, S.; Mukasa, Y.; Hashimoto, N.; Saito, K.; Noda, T.; Matsuura-Endo, C.; Zaidul, I.S.M.; Yamauchi, H. Changes in rutin concentration an flavonol-3-glucosidase activity during seedling growth in tartary buckwheat (Fagopyrum tataricum Gaertn.). Can. J. Plant. Sci. 2007, 87, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.C.; Sun, J.T. Changes in phenolic content, phenylalanine ammonia-lysyne (PAL) activity, and antioxidant capacity of two buckwheat sprouts in relation to germination. J. Funct. Foods 2014, 7, 298–304. [Google Scholar] [CrossRef]
- Nam, T.G.; Kim, D.O.; Eom, S.H. Effects of light sources on major flavonoids and antioxidant activity in common buckwheat sprouts. Food Sci. Biotechnol. 2018, 27, 169–176. [Google Scholar] [CrossRef]
- Qin, P.; Wei, A.; Zhao, D.; Yao, Y.; Yang, X.; Dun, B.; Ren, G. Low concentration of sodium bicarbonate improves the bioactive compounds levels and antioxidant an α-glucosidase inhibitory activities of tartary buckwheat sprouts. Food Chem. 2017, 224, 124–130. [Google Scholar] [CrossRef]
- Peng, L.X.; Zou, L.; Su, Y.M.; Fan, Y.; Zhao, G. Effects of light on growth, levels of anthocyanin, concentration of metabolites in Fagopyrum tataricum sprouts cultures. Int. J. Food Sci. Tech. 2015, 50, 1382–1389. [Google Scholar] [CrossRef]
- Weber, N.; Schmitzer, V.; Jakopic, J.; Stamper, F. First fruit in season: Seaweed extract and silicon advance organic strawberry (Fragaria × ananassa Duch.) fruit formation and yield. Sci. Hortic-Amsterdam. 2018, 242, 103–109. [Google Scholar] [CrossRef]
- Pereira, C.; Dias, M.I.; Petropoulos, S.A.; Plexida, S.; Chrysargyris, A.; Tzortzakis, N.; Calhelha, R.C.; Ivanov, M.; Stojković, D.; Soković, M.; et al. The effects of biostimulants, biofertilizers and water-stress on nutritional value and chemical composition of two spinach genotypes (Spinacia oleracea L.). Molecules 2019, 24, 4494. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor * | Antioxidant Activity | |||||
|---|---|---|---|---|---|---|
| FRAP 4′ (µM Fe2+/g) | FRAP 60′ (µM Fe2+/g) | FRAP Ratio (FRAP 60′/FRAP 4′) | DPPH (μM TRX/g) | ACW (μM AAE/g) | ACL (μM TRX/g) | |
| Cultivar ** | ||||||
| Panda | 50.44 ± 1.38 a | 99.50 ± 2.29 a | 1.973 ± 0.01 a | 46.88 ± 1.79 a | 163.5 ± 5.82 a | 1.132 ± 0.03 a |
| Kora | 54.52 ± 2.99 b | 110.00 ± 5.08 b | 2.018 ± 0.02 b | 52.94 ± 3.68 b | 182.7 ± 5.14 b | 1.250 ± 0.07 b |
| Treatment *** | ||||||
| Seaweed Ecklonia maxima extract | 39.83 ± 3.10 a | 83.03 ± 5.94 a | 2.085 ± 0.02 c | 41.23 ± 4.34 ab | 151.7 ± 19.9 a | 1.288 ± 0.07 a |
| Natural nitrophenols | 48.82 ± 5.83 c | 98.22 ± 10.4 c | 2.012 ± 0.03 bc | 52.03 ± 9.94 c | 166.6 ± 24.8 ab | 1.235 ± 0.15 a |
| Pythium oligandrum | 56.18 ± 0.50 e | 109.84 ± 1.58 de | 1.955 ± 0.02 ab | 51.90 ± 2.25 c | 184.6 ± 6.05 bc | 1.155 ± 0.06 a |
| Bacillus subtilis | 53.36 ± 4.39 d | 105.38 ± 6.99 d | 1.975 ± 0.05 ab | 44.10 ± 10.2 b | 166.8 ± 2.02 ab | 0.993 ± 0.06 a |
| Seaweed Ecklonia maxima extract + Pythium oligandrum | 62.47 ± 0.72 f | 121.31 ± 2.38 f | 1.942 ± 0.02 a | 56.33 ± 3.91 c | 195.6 ± 6.82 c | 1.568 ± 0.24 b |
| Natural nitrophenols + Pythium oligandrum | 52.84 ± 5.69 d | 105.22 ± 10.5 d | 1.991 ± 0.02 ab | 54.68 ± 7.54 c | 174.8 ± 16.3 b | 1.115 ± 0.06 a |
| Seaweed Ecklonia maxima extract + Bacillus subtilis | 43.92 ± 4.06 b | 90.25 ± 6.29 b | 2.055 ± 0.06 c | 37.28 ± 7.26 a | 153.8 ± 9.59 a | 1.215 ± 0.06 a |
| Natural nitrophenols + Bacillus subtilis | 53.03 ± 3.62 d | 104.73 ± 7.19 d | 1.975 ± 0.02 ab | 52.68 ± 3.75 c | 175.8 ± 7.50 b | 1.063 ± 0.03 a |
| Seaweed Ecklonia maxima extract + natural nitrophenols | 49.07 ± 3.30 c | 99.10 ± 4.47 c | 2.020 ± 0.05 bc | 43.68 ± 6.63 b | 175.1 ± 10.1 b | 1.062 ± 0.03 a |
| Control | 58.48 ± 1.40 e | 112.91 ± 2.64 e | 1.931 ± 0.02 a | 64.98 ± 5.18 d | 186.1 ± 5.22 bc | 1.218 ± 0.07 a |
| Factor * | Ingredient | ||||||
|---|---|---|---|---|---|---|---|
| Total Polyphenols (mg GAE/g) | Rutoside (mg/100 g) | Oxalic Acid (mg/100 g) | Malic Acid (mg/100 g) | Citric Acid (mg/100 g) | Succinic Acid (mg/100 g) | Fumaric Acid (mg/100 g) | |
| Cultivar ** | |||||||
| Panda | 40.42 ± 1.56 a | 280.1 ± 9.51 a | 144.0 ± 6.51 a | 527.2 ± 20.8 a | 388.1 ± 32.4 a | 490.7 ± 42.9 a | 81.38 ± 2.22 b |
| Kora | 43.42 ± 2.04 b | 305.6 ± 8.12 b | 124.5 ± 6.35 a | 622.7 ± 40.3 b | 343.1 ± 44.8 a | 537.4 ± 34.1 a | 74.87 ± 1.61 a |
| Treatment *** | |||||||
| Seaweed Ecklonia maxima extract | 35.52 ± 4.71 b | 311.2 ± 16.8 cd | 123.4 ± 17.8 a | 572.9 ± 118 abc | 217.0 ± 16.6 a | 368.2 ± 58.2 a | 74.94 ± 3.38 ab |
| Natural nitrophenols | 41.67 ± 5.63 d | 283.3 ± 17.3 abc | 119.2 ± 15.4 a | 517.6 ± 42.0 ab | 446.3 ± 107 a | 548.5 ± 102 a | 79.92 ± 4.50 bc |
| Pythium oligandrum | 44.62 ± 0.58 e | 306.4 ± 13.7 cd | 140.0 ± 13.6 a | 608.9 ± 42.4 bc | 391.8 ± 120 a | 492.2 ± 79.7 a | 76.57 ± 2.56 abc |
| Bacillus subtilis | 40.16 ± 4.26 c | 265.6 ± 15.9 abc | 144.0 ± 19.8 a | 564.8 ± 33.9 abc | 373.4 ± 85.3 a | 482.0 ± 106 a | 76.48 ± 4.43 abc |
| Seaweed Ecklonia maxima extract + Pythium oligandrum | 49.03 ± 0.71 g | 342.4 ± 7.52 d | 149.4 ± 18.6 a | 508.9 ± 23.6 ab | 428.0 ± 100 a | 629.4 ± 95.6 a | 76.55 ± 2.66 abc |
| Natural nitrophenols + Pythium oligandrum | 44.28 ± 5.49 e | 299.4 ± 25.7 abc | 136.9 ± 17.0 a | 471.3 ± 32.6 ab | 390.6 ± 118 a | 525.2 ± 97.6 a | 80.65 ± 3.65 bc |
| Seaweed Ecklonia maxima extract + Bacillus subtilis | 34.16 ± 4.07 a | 253.0 ± 8.81 ab | 125.9 ± 20.8 a | 683.8 ± 73.2 c | 340.8 ± 109 a | 438.1 ± 29.7 a | 66.53 ± 2.93 a |
| Natural nitrophenols + Bacillus subtilis | 44.14 ± 3.55 e | 241.1 ± 30.7 a | 130.9 ± 12.5 a | 449.7 ± 25.4 a | 397.9 ± 108 a | 629.6 ± 96.7 a | 70.23 ± 1.73 ab |
| Seaweed Ecklonia maxima extract + natural nitrophenols | 38.99 ± 3.88 c | 309.3 ± 17.0 cd | 130.4 ± 8.44 a | 835.2 ± 119 d | 339.4 ± 73.0 a | 419.4 ± 28.8 a | 87.27 ± 6.07 c |
| Control | 46.62 ± 0.56 f | 316.9 ± 16.1 cd | 131.6 ± 13.9 a | 561.0 ± 26.0 abc | 308.8 ± 69.0 a | 582.4 ± 83.8 a | 87.79 ± 1.91 c |
| Item | FRAP 4′ | FRAP 60′ | TP | DPPH | ACW | ACL | FRAP Ratio | Rutoside | Oxalic Acid | Malic Acid | Citric Acid | Succinic Acid |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FRAP 60′ | 0.9978 p = 0.000 | 1.0000 p = --- | ||||||||||
| TP | 0.9645 p = 0.000 | 0.9609 p = 0.000 | 1.0000 p = --- | |||||||||
| DPPH | 0.8990 p = 0.000 | 0.8853 p = 0.000 | 0.9063 p = 0.000 | 1.0000 p = --- | ||||||||
| ACW | 0.8182 p = 0.000 | 0.8207 p = 0.000 | 0.8837 p = 0.000 | 0.8100 p = 0.000 | 1.0000 p = --- | |||||||
| ACL | 0.2682 p = 0.094 | 0.2730 p = 0.088 | 0.2000 p = 0.216 | 0.2900 p = 0.069 | 0.2550 p = 0.112 | 1.0000 p = --- | ||||||
| FRAP Ratio | −0.8897 p = 0.000 | −0.8609 p = 0.000 | −0.8847 p = 0.000 | −0.8715 p = 0.000 | −0.7190 p = 0.000 | −0.1697 p = 0.295 | 1.0000 p = --- | |||||
| Rutoside | 0.5990 p = 0.000 | 0.6033 p = 0.000 | 0.6341 p = 0.000 | 0.6280 p = 0.000 | 0.5409 p = 0.000 | 0.3053 p = 0.055 | −0.4832 p = 0.002 | 1.0000 p = --- | ||||
| Oxalic acid | 0.1230 p = 0.449 | 0.1091 p = 0.503 | 0.0798 p = 0.625 | 0.0580 p = 0.722 | 0.0127 p = 0.938 | 0.1425 p = 0.380 | −0.2425 p = 0.132 | −0.0552 p = 0.735 | 1.0000 p = --- | |||
| Malic acid | −0.3870 p = 0.014 | −0.3593 p = 0.023 | −0.3551 p = 0.025 | −0.4268 p = 0.006 | −0.1180 p = 0.468 | −0.1603 p = 0.323 | 0.5141 p = 0.001 | −0.1751 p = 0.280 | 0.0488 p = 0.765 | 1.0000 p = --- | ||
| Citric acid | 0.2353 p = 0.144 | 0.2243 p = 0.164 | 0.2286 p = 0.156 | 0.1486 p = 0.360 | 0.2797 p = 0.080 | 0.0165 p = 0.920 | −0.3043 p = 0.056 | −0.0617 p = 0.705 | 0.5888 p = 0.000 | −0.0324 p = 0.842 | 1.0000 p = --- | |
| Succinic acid | 0.4981 p = 0.001 | 0.4889 p = 0.001 | 0.4471 p = 0.004 | 0.3939 p = 0.012 | 0.3618 p = 0.022 | 0.1381 p = 0.396 | −0.4667 p = 0.002 | 0.1369 p = 0.399 | 0.0604 p = 0.711 | −0.1160 p = 0.476 | 0.2256 p = 0.162 | 1.0000 p = --- |
| Fumaric acid | 0.0084 p = 0.959 | −0.0149 p = 0.927 | 0.0896 p = 0.583 | 0.1622 p = 0.317 | −0.0231 p = 0.887 | −0.0727 p = 0.656 | −0.1887 p = 0.244 | 0.3119 p = 0.050 | −0.0602 p = 0.712 | −0.2532 p = 0.115 | −0.1352 p = 0.406 | −0.0848 p = 0.603 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Witkowicz, R.; Biel, W.; Skrzypek, E.; Chłopicka, J.; Gleń-Karolczyk, K.; Krupa, M.; Prochownik, E.; Galanty, A. Microorganisms and Biostimulants Impact on the Antioxidant Activity of Buckwheat (Fagopyrum esculentum Moench) Sprouts. Antioxidants 2020, 9, 584. https://doi.org/10.3390/antiox9070584
Witkowicz R, Biel W, Skrzypek E, Chłopicka J, Gleń-Karolczyk K, Krupa M, Prochownik E, Galanty A. Microorganisms and Biostimulants Impact on the Antioxidant Activity of Buckwheat (Fagopyrum esculentum Moench) Sprouts. Antioxidants. 2020; 9(7):584. https://doi.org/10.3390/antiox9070584
Chicago/Turabian StyleWitkowicz, Robert, Wioletta Biel, Edyta Skrzypek, Joanna Chłopicka, Katarzyna Gleń-Karolczyk, Mateusz Krupa, Ewelina Prochownik, and Agnieszka Galanty. 2020. "Microorganisms and Biostimulants Impact on the Antioxidant Activity of Buckwheat (Fagopyrum esculentum Moench) Sprouts" Antioxidants 9, no. 7: 584. https://doi.org/10.3390/antiox9070584