
The Ionic Selectivity of Lysenin Channels in Open and Sub-Conducting States


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
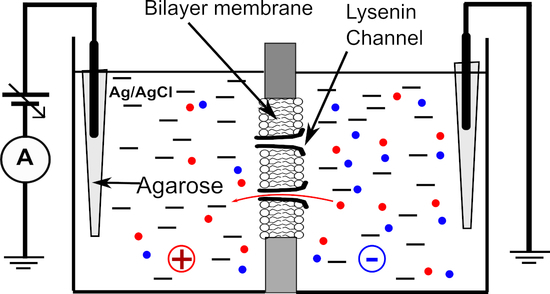
2.2.1. BLM Production, Characterization, and Channel Insertion
2.2.2. Voltage Measurements
2.2.3. Investigations on Sub-Conducting Channels
3. Results
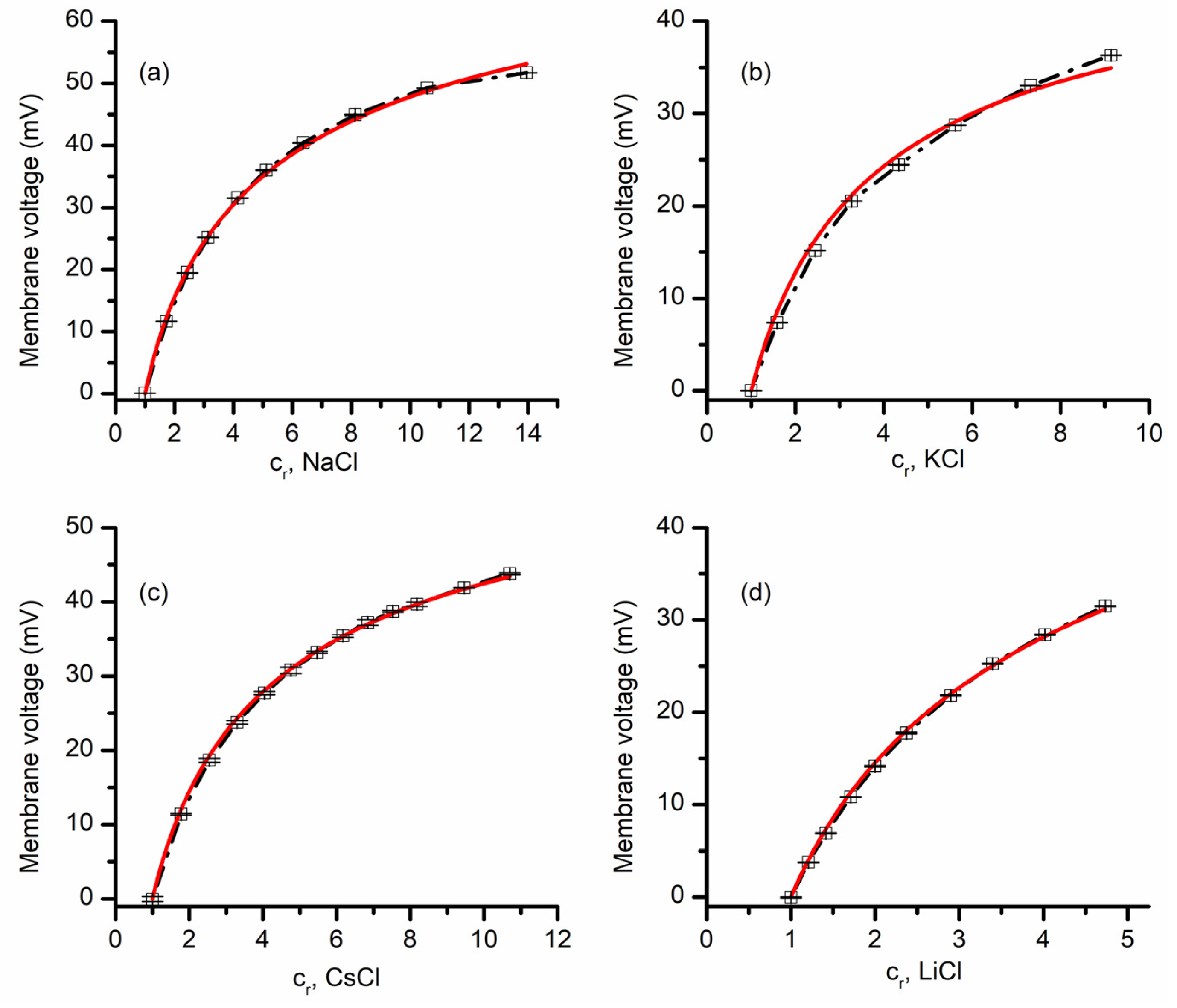
3.1. Lysenin Channels in the Open State Are Cation-Selective
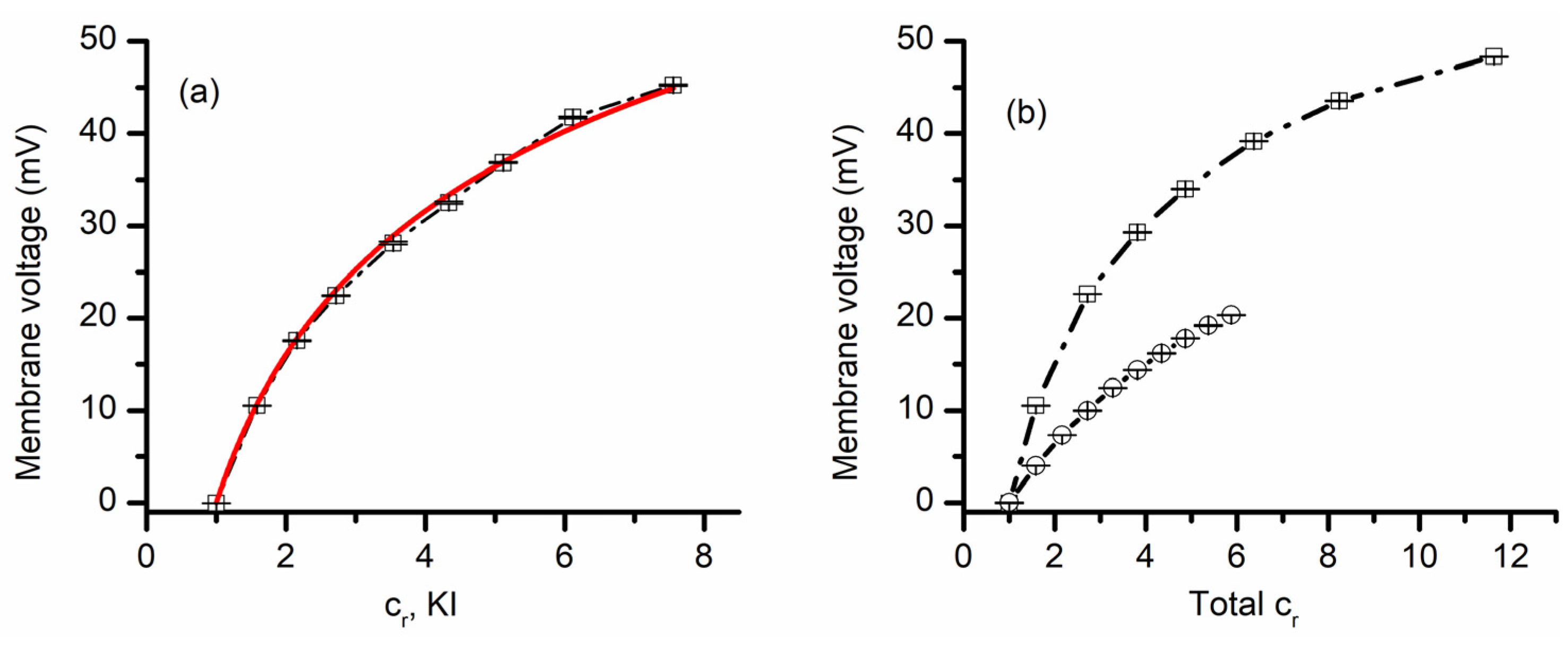
3.2. Lysenin Channels in an Open State Present Different Permeability for Anions
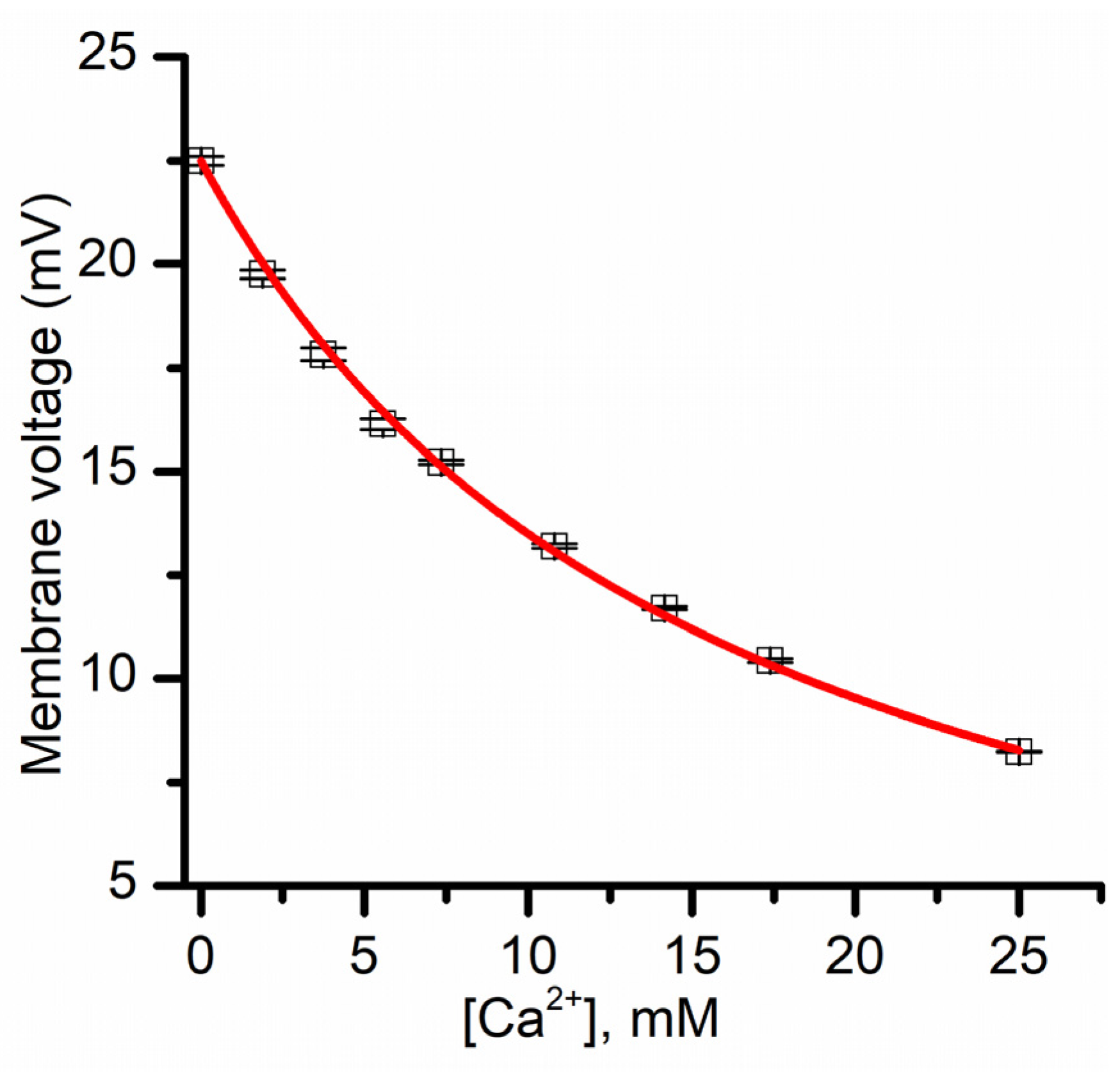
3.3. Investigations on the Selectivity of Lysenin Channels in Sub-Conducting States
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hille, B. Ion. Channels of Excitable Membranes, 3rd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001. [Google Scholar]
- Bezanilla, F. Ion channels: From conductance to structure. Neuron 2008, 60, 456–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho-de-Souza, J.L.; Saponaro, A.; Bassetto, C.A.Z.; Rauh, O.; Schroeder, I.; Franciolini, F.; Catacuzzeno, L.; Bezanilla, F.; Thiel, G.; Moroni, A. Experimental challenges in ion channel research: Uncovering basic principles of permeation and gating in potassium channels. Adv. Phys.-X 2022, 7, 1978317. [Google Scholar] [CrossRef]
- De Lera Ruiz, M.; Kraus, R.L. Voltage-gated sodium channels: Structure, function, pharmacology, and clinical indications. J. Med. Chem. 2015, 58, 7093–7118. [Google Scholar] [CrossRef]
- Dudev, T.; Lim, C. Ion selectivity strategies of sodium channel selectivity filters. Acc. Chem. Res. 2014, 47, 3580–3587. [Google Scholar] [CrossRef] [PubMed]
- Roux, B. Ion channels and ion selectivity. Essays Biochem. 2017, 61, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Cymes, G.D.; Grosman, C. Identifying the elusive link between amino acid sequence and charge selectivity in pentameric ligand-gated ion channels. Proc. Nat. Acad. Sci. USA 2016, 113, E7106. [Google Scholar] [CrossRef] [Green Version]
- Westhoff, M.; Eldstrom, J.; Murray, C.I.; Thompson, E.; Fedida, D. IKs ion-channel pore conductance can result from individual voltage sensor movements. Proc. Nat. Acad. Sci. USA 2019, 116, 7879–7888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul Kadir, L.; Stacey, M.; Barrett-Jolley, R. Emerging roles of the membrane potential: Action beyond the action potential. Front. Physiol. 2018, 9, 1661. [Google Scholar] [CrossRef]
- Peraro, M.D.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Bainbridge, G.; Gokce, I.; Lakey, J.H. Voltage gating is a fundamental feature of porin and toxin β-barrel membrane channels. FEBS Lett. 1998, 431, 305–308. [Google Scholar] [CrossRef] [Green Version]
- Fologea, D.; Krueger, E.; Al Faori, R.; Lee, R.; Mazur, Y.I.; Henry, R.; Arnold, M.; Salamo, G.J. Multivalent ions control the transport through lysenin channels. Biophys. Chem. 2010, 152, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Ide, T.; Aoki, T.; Takeuchi, Y.; Yanagida, T. Lysenin forms a voltage-dependent channel in artificial lipid bilayer membranes. Biochem. Biophys. Res. Commun. 2006, 346, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Bogard, A.; Abatchev, G.; Hutchinson, Z.; Ward, J.; Finn, P.W.; McKinney, F.; Fologea, D. Lysenin Channels as sensors for ions and molecules. Sensors 2020, 20, 6099. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Hirano, M.; Takeuchi, Y.; Kobayashi, T.; Yanagida, T.; Ide, T. Single channel properties of lysenin measured in artificial lipid bilayers and their applications to biomolecule detection. Proc. Jpn. Acad. Ser. B 2010, 86, 920–925. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.-Q.; Dalla Serra, M.; Vincent, J.B.; Vigh, G.; Cheley, S.; Braha, O.; Bayley, H. Reversal of charge selectivity in transmembrane protein pores by using noncovalent molecular adapters. Proc. Nat. Acad. Sci. USA 2000, 97, 3959–3964. [Google Scholar] [CrossRef] [Green Version]
- Kwiatkowska, K.; Hordejuk, R.; Szymczyk, P.; Kulma, M.; Abdel-Shakor, A.-B.; Plucienniczak, A.; Dolowy, K.; Szewczyk, A.; Sobota, A. Lysenin-His, a sphingomyelin-recognizing toxin, requires tryptophan 20 for cation-selective channel assembly but not for membrane binding. Mol. Membr. Biol. 2007, 24, 121–134. [Google Scholar] [CrossRef]
- Nestorovich, E.M.; Karginov, V.A.; Bezrukov, S.M. Polymer partitioning and ion selectivity suggest asymmetrical shape for the membrane pore formed by epsilon toxin. Biophys. J. 2010, 99, 782–789. [Google Scholar] [CrossRef] [Green Version]
- Ostolaza, H.; Gonzalez-Bullon, D.; Uribe, K.B.; Martin, C.; Amuategi, J.; Fernandez-Martinez, X. Membrane permeabilization by pore-forming RTX toxins: What kind of lesions do these toxins form? Toxins 2019, 11, 354. [Google Scholar] [CrossRef] [Green Version]
- Shogomori, H.; Kobayashi, T. Lysenin: A sphingomyelin specific pore-forming toxin. Biochim. Biophys. Acta Gen. Subj. 2008, 1780, 612–618. [Google Scholar] [CrossRef]
- Yamaji-Hasegawa, A.; Makino, A.; Baba, T.; Senoh, Y.; Kimura-Suda, H.; Sato, S.B.; Terada, N.; Ohno, S.; Kiyokawa, E.; Umeda, M.; et al. Oligomerization and pore formation of a sphingomyelin-specific toxin, lysenin. J. Biol. Chem. 2003, 278, 22762–22770. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, N.; Yamada, T.; Greimel, P.; Uchihashi, T.; Ando, T.; Kobayashi, T. Real-time visualization of assembling of a sphingomyelin-specific toxin on planar lipid membranes. Biophys. J. 2013, 105, 1397–1405. [Google Scholar] [CrossRef] [Green Version]
- Shakor, A.-B.A.; Czurylo, E.A.; Sobota, A. Lysenin, a unique sphingomyelin-binding protein. FEBS Lett. 2003, 542, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bruhn, H.; Winkelmann, J.; Andersen, C.; Andra, J.; Leippe, M. Dissection of the mechanisms of cytolytic and antibacterial activity of lysenin, a defense protein of the annelid Eisenia fetida. Dev. Comp. Immunol. 2006, 30, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Kobayashi, T. Assemblies of pore-forming toxins visualized by atomic force microscopy. Biochim. Biophys. Acta—Biomembr. 2016, 1858, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Yamaji-Hasegawa, A.; Hullin-Matsuda, F.; Kobayashi, T. Molecular mechanisms of action of sphingomyelin-specific pore-forming toxin, lysenin. Semin. Cell Dev. Biol. 2018, 73, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Najbauer, E.E.; Becker, S.; Giller, K.; Zweckstetter, M.; Lange, A.; Steinem, C.; de Groot, B.L.; Griesinger, C.; Andreas, L.B. Structure, gating and interactions of the voltage-dependent anion channel. Eur. Biophys. J. 2021, 50, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Sander, P.; Gudermann, T.; Schredelseker, J. A calcium guard in the outer membrane: Is VDAC a regulated gatekeeper of mitochondrial calcium uptake? Int. J. Mol. Sci 2021, 22, 946. [Google Scholar] [CrossRef] [PubMed]
- Briones, R.; Weichbrodt, C.; Paltrinieri, L.; Mey, I.; Villinger, S.; Giller, K.; Lange, A.; Zweckstetter, M.; Griesinger, C.; Becker, S.; et al. Voltage dependence of conformational dynamics and subconducting states of VDAC-1. Biophys. J. 2016, 111, 1223–1234. [Google Scholar] [CrossRef] [Green Version]
- Gincel, D.; Silberberg, S.D.; Shoshan-Barmatz, V. Modulation of the Voltage-Dependent Anion Channel (VDAC) by glutamate. J. Bioenerg. Biomembr. 2000, 32, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.D.; Nomura, T.; Ziegler, C.S.; Campbell, A.K.; Wann, K.T.; Martinac, B. Selectivity mechanism of the mechanosensitive channel MscS revealed by probing channel subconducting states. Nat. Commun. 2013, 4, 2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flegler, V.J.; Rasmussen, T.; Böttcher, B. More than just closed and open: Unraveling a mechanosensor. Trends Biochem. Sci. 2021, 46, 623–625. [Google Scholar] [CrossRef]
- Shapovalov, G.; Lester, H.A. Gating transitions in bacterial ion channels measured at 3 microns resolution. J. Gen. Physiol. 2004, 124, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Jo, A.; Hoi, H.; Zhou, H.; Gupta, M.; Montemagno, C.D. Single-molecule study of full-length NaChBac by planar lipid bilayer recording. PLoS ONE 2017, 12, e0188861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, M.L.; VanDongen, A.M.J. K Channel subconductance levels result from heteromeric pore conformations. J. Gen. Physiol. 2005, 126, 87–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.W.L.; Enkvetchakul, D.; Nichols, C.G. KirBac1.1: It’s an inward rectifying potassium channel. J. Gen. Physiol. 2009, 133, 295–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warner, L.R.; Mass, O.; Donnelly, L.N.; Grantham, B.R.; Oxford, J.T. Growing and handling of bacterial cultures within a shared core facility for integrated structural biology program. In Growing and Handling of Bacterial Cultures; Mishra, M., Ed.; IntechOpen Limited: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Duong-Ly, K.C.; Gabelli, S.B.; Lorsch, J.R. Affinity purification of a recombinant protein expressed as a fusion with the Maltose-Binding Protein (MBP) tag. In Methods in Enzymology; Lorsch, J.R., Ed.; Academic Press: Amsterdam, The Netherlands, 2015; Volume 559, pp. 17–26. [Google Scholar] [CrossRef] [Green Version]
- Jenny, R.J.; Mann, K.G.; Lundblad, R.L. A critical review of the methods for cleavage of fusion proteins with thrombin and factor Xa. Protein Expr. Purif. 2003, 31, 1–11. [Google Scholar] [CrossRef]
- Lebendiker, M.; Danieli, T. Purification of proteins fused to maltose-binding protein. In Protein Chromatography: Methods and Protocols; Walls., D., Loughran, S.T., Eds.; Humana Press: New York, NY, USA, 2017; pp. 257–273. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bogard, A.; Finn, P.W.; McKinney, F.; Flacau, I.M.; Smith, A.R.; Whiting, R.; Fologea, D. The Ionic Selectivity of Lysenin Channels in Open and Sub-Conducting States. Membranes 2021, 11, 897. https://doi.org/10.3390/membranes11110897
Bogard A, Finn PW, McKinney F, Flacau IM, Smith AR, Whiting R, Fologea D. The Ionic Selectivity of Lysenin Channels in Open and Sub-Conducting States. Membranes. 2021; 11(11):897. https://doi.org/10.3390/membranes11110897
Chicago/Turabian StyleBogard, Andrew, Pangaea W. Finn, Fulton McKinney, Ilinca M. Flacau, Aviana R. Smith, Rosey Whiting, and Daniel Fologea. 2021. "The Ionic Selectivity of Lysenin Channels in Open and Sub-Conducting States" Membranes 11, no. 11: 897. https://doi.org/10.3390/membranes11110897
APA StyleBogard, A., Finn, P. W., McKinney, F., Flacau, I. M., Smith, A. R., Whiting, R., & Fologea, D. (2021). The Ionic Selectivity of Lysenin Channels in Open and Sub-Conducting States. Membranes, 11(11), 897. https://doi.org/10.3390/membranes11110897