
Every Detail Matters. That Is, How the Interaction between Gα Proteins and Membrane Affects Their Function


Abstract
:1. Introduction
2. What Makes Gα Dock to the Membrane?
2.1. Palmitoylation
2.2. N-Myristoylation
2.3. Prenylation
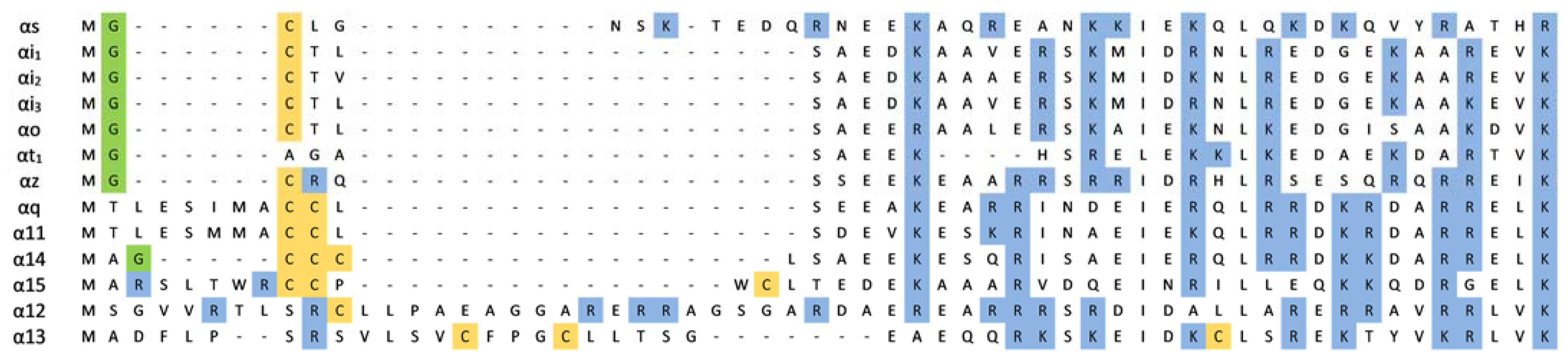
2.4. Polybasic Motifs
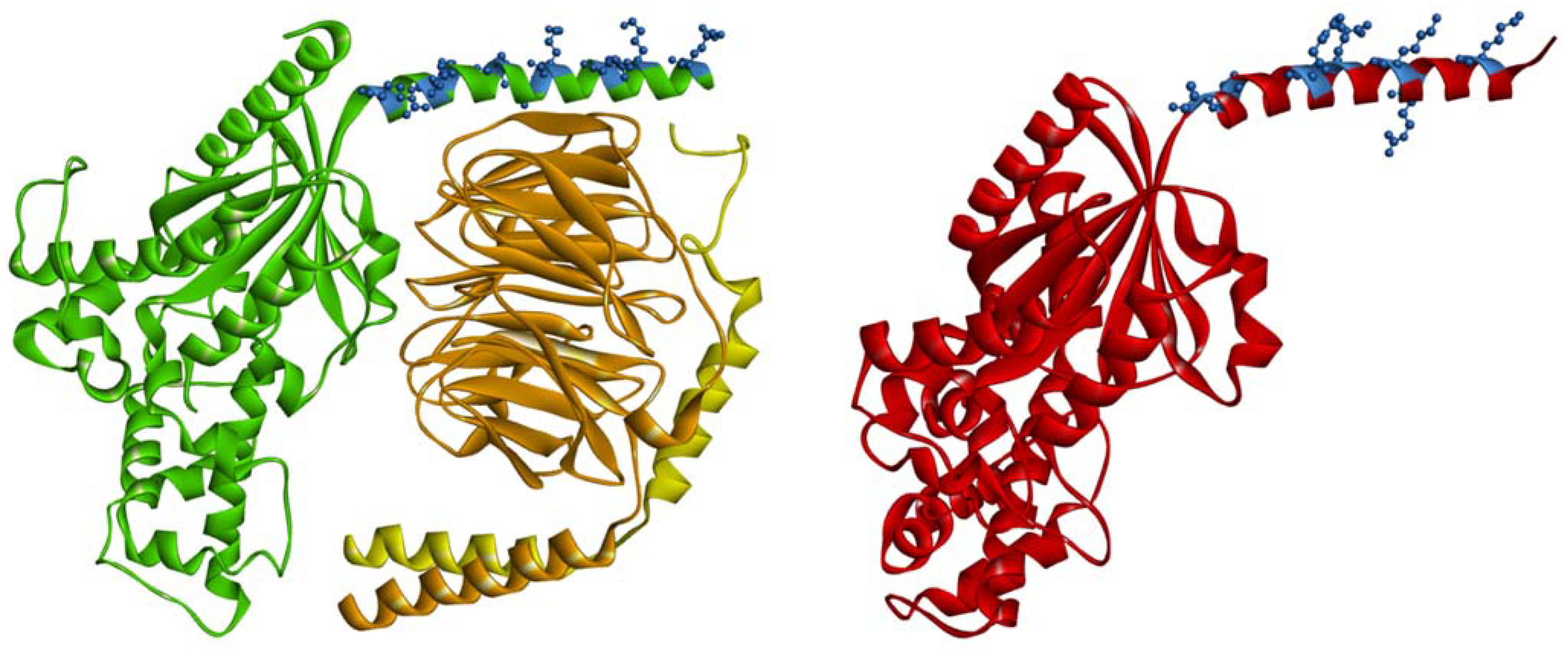
3. Looking from the Plasma Membrane Side; What Makes Gα Proteins Associate with It?
4. Does It Matter to Us? Why Is It Worth Researching?
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Neer, E.J. Heterotrimeric G Proteins: Organizers of Transmembrane Signals. Cell 1995, 80, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Milligan, G.; Kostenis, E. Heterotrimeric G-Proteins: A Short History. Br. J. Pharmacol. 2006, 147, S46–S55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilman, A.G. G Proteins: Transducers of Receptor-Generated Signals. Annu. Rev. Biochem. 1987, 56, 615–649. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Vera, T.M.; Vanhauwe, J.; Thomas, T.O.; Medkova, M.; Preininger, A.; Mazzoni, M.R.; Hamm, H.E. Insights into G Protein Structure, Function, and Regulation. Endocr. Rev. 2003, 24, 765–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takai, Y.; Sasaki, T.; Matozaki, T. Small GTP-Binding Proteins. Physiol. Rev. 2001, 81, 153–208. [Google Scholar] [CrossRef] [PubMed]
- Downes, G.B.; Gautam, N. The G Protein Subunit Gene Families. Genomics 1999, 62, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Okashah, N.; Wan, Q.; Ghosh, S.; Sandhu, M.; Inoue, A.; Vaidehi, N.; Lambert, N.A. Variable G Protein Determinants of GPCR Coupling Selectivity. Proc. Natl. Acad. Sci. USA 2019, 116, 12054–12059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldham, W.M.; Hamm, H.E. Heterotrimeric G Protein Activation by G-Protein-Coupled Receptors. Nat. Rev. Mol. Cell Biol. 2008, 9, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, J.P.; Sunahara, R.K. Mechanistic Insights into GPCR–G Protein Interactions. Curr. Opin. Struct. Biol. 2016, 41, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Lambright, D.G.; Sondek, J.; Bohm, A.; Skiba, N.P.; Hamm, H.E.; Sigler, P.B. The 2.0 Å Crystal Structure of a Heterotrimeric G Protein. Nature 1996, 379, 311–319. [Google Scholar] [CrossRef]
- Wall, M.A.; Coleman, D.E.; Lee, E.; Iaiguez-lluhi, J.A.; Posner, B.A.; Gilman, A.G.; Ft, S. The Structure of the G Protein Heterotrimer Giα1β1γ2. Cell 1995, 83, 1047–1058. [Google Scholar] [CrossRef] [Green Version]
- Syrovatkina, V.; Alegre, K.O.; Dey, R.; Huang, X. Regulation, Signaling and Physiological Functions of G-Proteins. J. Mol. Biol. 2016, 428, 3850–3868. [Google Scholar] [CrossRef] [Green Version]
- Clapham, D.E.; Neer, E.J. G Protein Βγ Subunits. Annu. Rev. Pharmacol. Sci. 1997, 37, 167–203. [Google Scholar] [CrossRef] [PubMed]
- Hillenbrand, M.; Schori, C.; Schöppe, J.; Plückthun, A. Comprehensive Analysis of Heterotrimeric G-Protein Complex Diversity and Their Interactions with GPCRs in Solution. Proc. Natl. Acad. Sci. USA 2015, 112, E1181–E1190. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.J.; Thomas, T.C.; Levinell, M.A.; Neers, E.J. Specificity of G Protein β and γ. J. Biol. Chem. 1992, 267, 13807–13810. [Google Scholar] [CrossRef]
- Wall, M.A.; Posner, B.A.; Sprang, S.R. Structural Basis of Activity and Subunit Recognition in G Protein Heterotrimers. Structure 1998, 6, 1169–1183. [Google Scholar] [CrossRef] [Green Version]
- Ford, C.E.; Skiba, N.P.; Bae, H.; Daaka, Y.; Reuveny, E.; Shekter, L.R.; Rosal, R.; Weng, G.; Yang, C.S.; Iyengar, R.; et al. Molecular Basis for Interactions of G Protein Βγ Subunits with Effectors. Science 1998, 280, 1271–1274. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.M.; Sleno, R.; Gora, S.; Zylbergold, P.; Laverdure, J.-P.; Labbe, J.-C.; Miller, G.J.; Hebert, T.E. The Expanding Roles of Gβγ Subunits in G Protein-Coupled Receptor Signaling and Drug Action. Pharmacol. Rev. 2013, 65, 545–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondar, A.; Lazar, J. Dissociated GαGTP and Gβγ Protein Subunits Are the Major Activated Form of Heterotrimeric Gi/o Proteins. J. Biol. Chem. 2014, 289, 1271–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobingier, B.T.; von Zastrow, M. When Trafficking and Signaling Mix: How Subcellular Location Shapes G Protein-Coupled Receptor Activation of Heterotrimeric G Proteins. Traffic 2019, 20, 130–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedegaertner, P.B. G Protein Trafficking. Subcell Biochem. 2012, 63, 193–223. [Google Scholar] [CrossRef] [PubMed]
- Nadolski, M.J.; Linder, M.E. Protein Lipidation. FEBS J. 2007, 274, 5202–5210. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Casey, P.J. Protein Prenylation: Unique Fats Make Their Mark on Biology. Nat. Rev. Mol. Cell Biol. 2016, 17, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhang, X.; Chen, X.; Aramsangtienchai, P.; Tong, Z.; Lin, H. Protein Lipidation: Occurrence, Mechanisms, Biological Functions, and Enabling Technologies. Chem. Rev. 2018, 118, 919–988. [Google Scholar] [CrossRef] [PubMed]
- Hiol, A.; Davey, P.C.; Osterhout, J.L.; Waheed, A.A.; Fischer, E.R.; Chen, C.K.; Milligan, G.; Druey, K.M.; Jones, T.L.Z. Palmitoylation Regulates Regulators of G-Protein Signaling (RGS) 16 Function: I. Mutation of Amino-Terminal Cysteine Residues on RGS16 Prevents Its Targeting to Lipid Rafts and Palmitoylation of an Internal Cysteine Residue. J. Biol. Chem. 2003, 278, 19301–19308. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.J.Y.; Gabriel, F.; Tandale, A.; Nietlispach, D. Structure and Dynamics of GPCRs in Lipid Membranes: Physical Principles and Experimental Approaches. Molecules 2020, 25, 4729. [Google Scholar] [CrossRef] [PubMed]
- Villar, V.A.M.; Cuevas, S.; Zheng, X.; Jose, P.A. Localization and Signaling of GPCRs in Lipid Rafts. Methods Cell Biol. 2016, 132, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Fallahi-Sichani, M.; Linderman, J.J. Lipid Raft-Mediated Regulation of G-Protein Coupled Receptor Signaling by Ligands Which Influence Receptor Dimerization: A Computational Study. PLoS ONE 2009, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lukasheva, V.; Devost, D.; Le Gouill, C.; Aoki, J. Signal Profiling of the Β1AR Reveals Coupling to Novel Signalling Pathways and Distinct Phenotypic Responses Mediated by Β1AR and Β2AR. Sci. Rep. 2020, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Polit, A.; Rysiewicz, B.; Mystek, P.; Błasiak, E.; Wasylewska, M.D. The Gαi Protein Subclass Selectivity to the Dopamine -D2 Receptor Is Also Decided by Their Location at the Cell Membrane. Cell Commun. Signal. 2020, 18, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mystek, P.; Rysiewicz, B.; Gregrowicz, J.; Dziedzicka-Wasylewska, M.; Polit, A. Gγ and Gα Identity Dictate a G-Protein Heterotrimer Plasma Membrane Targeting. Cells 2019, 8, 1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, J.P.; Paterson, H.; Marshalit, C.J. A Polybasic Domain or Palmitoylation Is Required in Addition to the CAAX Motif to Localize P21ras to the Plasma Membrane. Cell 1990, 63, 133–139. [Google Scholar] [CrossRef]
- Thaker, T.M.; Sarwar, M.; Preininger, A.M.; Hamm, H.E.; Iverson, T.M. A Transient Interaction between the Phosphate Binding Loop and Switch i Contributes to the Allosteric Network between Receptor and Nucleotide in Gαi1. J. Biol. Chem. 2014, 289, 11331–11341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyon, A.M.; Begley, J.A.; Manett, T.D.; Tesmer, J.J.G. Molecular Mechanisms of Phospholipase C Β3 Autoinhibition. Structure 2014, 22, 1844–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mou, T.C.; Gille, A.; Suryanarayana, S.; Richter, M.; Seifert, R.; Sprang, S.R. Broad Specificity of Mammalian Adenylyl Cyclase for Interaction with 2′,3′-Substituted Purine- and Pyrimidine Nucleotide Inhibitors. Mol. Pharmacol. 2006, 70, 878–886. [Google Scholar] [CrossRef]
- Kimple, A.J.; Soundararajan, M.; Hutsell, S.Q.; Roos, A.K.; Urban, D.J.; Setola, V.; Temple, B.R.S.; Roth, B.L.; Knapp, S.; Willard, F.S.; et al. Structural Determinants of G-Protein α Subunit Selectivity by Regulator of G-Protein Signaling 2 (RGS2). J. Biol. Chem. 2009, 284, 19402–19411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crouthamel, M.; Abankwa, D.; Zhang, L.; Dilizio, C.; Manning, D.R.; Hancock, J.F.; Wedegaertner, P.B. An N-Terminal Polybasic Motif of Gαq Is Required for Signaling and Influences Membrane Nanodomain Distribution. Mol. Pharmacol. 2010, 78, 767–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosloff, M.; Elia, N.; Selinger, Z. Structural Homology Discloses a Bifunctional Structural Motif at the N-Termini of Gα Proteins. Biochemistry 2002, 41, 14518–14523. [Google Scholar] [CrossRef] [PubMed]
- Simossis, V.A.; Kleinjung, J.; Heringa, J. Homology-Extended Sequence Alignment. Nucleic Acids Res. 2005, 33, 816–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mixon, M.B.; Lee, E.; Coleman, D.E.; Berghuis, A.M.; Gilman, A.G.; Sprang, S.R. Tertiary and Quaternary Structural Changes in Gial Induced by GTP Hydrolysis. Science 1995, 270, 954–960. [Google Scholar] [CrossRef]
- Van Eps, N.; Altenbach, C.; Caro, L.N.; Latorraca, N.R.; Hollingsworth, S.A.; Dror, R.O.; Ernst, O.P.; Hubbell, W.L. Gi- and Gs-Coupled GPCRs Show Different Modes of G-Protein Binding. PNAS 2018, 115, 2383–2388. [Google Scholar] [CrossRef] [Green Version]
- Smotrys, J.E.; Linder, M.E. Palmitoylation of Intracellular Signalling Proteins: Regulation and Function. Annu. Rev. Biochem. 2004, 73, 559–587. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.E.; Deschenes, R.J. Palmitoylation: Policing Protein Stability and Traffic. Nat. Rev. Mol. Cell Biol. 2007, 8, 74–84. [Google Scholar] [CrossRef]
- Salaun, C.; Greaves, J.; Chamberlain, L.H. The Intracellular Dynamic of Protein Palmitoylation. J. Cell Biol. 2010, 191, 1229–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plain, F.; Congreve, S.D.; Yee, R.S.Z.; Kennedy, J.; Howie, J.; Kuo, C.W.; Fraser, N.J.; Fuller, W. An Amphipathic α-Helix Directs Palmitoylation of the Large Intracellular Loop of the Sodium/Calcium Exchanger. J. Biol. Chem. 2017, 292, 10745–10752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Politis, E.G.; Roth, A.F.; Davis, N.G. Transmembrane Topology of the Protein Palmitoyl Transferase Akr1. J. Biol. Chem. 2005, 280, 10156–10163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobin, A.B.; Wheatley, M. G-Protein-Coupled Receptor Phosphorylation and Palmitoylation. Methods Mol. Biol. 2004, 259, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.E.; Middleton, P.; Hepler, J.R.; Taussig, R.; Gilman, A.G.; Mumby, S.M. Lipid Modifications of G Proteins: α Subunits Are Palmitoylated. Proc. Natl. Acad. Sci. USA 1993, 90, 3675–3679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedegaertner, P.B.; Chu, D.H.; Wilson, P.T.; Levis, M.J.; Bourne, H.R. Palmitoylation Is Required for Signaling Functions and Membrane Attachment of G(q)α and G(s)α. J. Biol. Chem. 1993, 268, 25001–25008. [Google Scholar] [CrossRef]
- Nini, L.; Waheed, A.A.; Panicker, L.M.; Czapiga, M.; Zhang, J.H.; Simonds, W.F. R7-Binding Protein Targets the G Protein Β5/R7-Regulator of G Protein Signaling Complex to Lipid Rafts in Neuronal Cells and Brain. BMC Biochem. 2007, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charych, E.I.; Jiang, L.; Lo, F.; Sullivan, K.; Brandon, N.J. Interplay of Palmitoylation and Phosphorylation in the Trafficking and Localization of Phosphodiesterase 10A: Implications for the Treatment of Schizophrenia. J. Neurosci. Neuro 2010, 30, 9027–9037. [Google Scholar] [CrossRef] [Green Version]
- Stoffel, R.H.; Randall, R.R.; Premont, R.T.; Lefkowitz, R.J.; Inglese, J. Palmitoylation of G Protein-Coupled Receptor Kinase, GRK6. Lipid Modification Diversity in the GRK Family. J. Biol. Chem. 1994, 269, 27791–27794. [Google Scholar] [CrossRef]
- Premont, R.T.; Inglese, J.; Lefkowitz, R.J. Protein Kinases That Phosphorylate Activated G Protein-coupled Receptors. FASEB J. 1995, 9, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Marsden, A.N.; Akimzhanov, A.M.; Boehning, D. The Palmitoyl Acyltransferase DHHC5 Mediates Beta-Adrenergic Signaling in the Heart by Targeting Gα Proteins and G Protein-Coupled Receptor Kinase 2. bioRxiv 2018, 371211, 1–16. [Google Scholar] [CrossRef]
- Qanbar, R.; Bouvier, M. Role of Palmitoylation/Depalmitoylation Reactions in G-Protein-Coupled Receptor Function. Pharmacol. Ther. 2003, 97, 1–33. [Google Scholar] [CrossRef]
- Escribá, P.V.; Wedegaertner, P.B.; Goñi, F.M.; Vögler, O. Lipid–Protein Interactions in GPCR-Associated Signaling. Biochim. Et Biophys. Acta Biomembr. 2007, 1768, 836–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delint-Ramirez, I.; Willoughby, D.; Hammond, G.R.V.; Ayling, L.J.; Cooper, D.M.F. Palmitoylation Targets AKAP79 Protein to Lipid Rafts and Promotes Its Regulation of Calcium-Sensitive Adenylyl Cyclase Type 8. J. Biol. Chem. 2015, 290, 32962–32975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levental, I.; Grzybek, M.; Simons, K. Greasing Their Way: Lipid Modifications Determine Protein Association with Membrane Rafts. Biochemistry 2010, 49, 6305–6316. [Google Scholar] [CrossRef]
- Zhang, T.; Luo, Q.; Yang, L.; Jiang, H.; Yang, H. Characterizing the Interactions of Two Lipid Modifications with Lipid Rafts: Farnesyl Anchors vs. Palmitoyl Anchors. Eur. Biophys. J. 2017, 47, 19–30. [Google Scholar] [CrossRef]
- Huang, K.; El-Husseini, A. Modulation of Neuronal Protein Trafficking and Function by Palmitoylation. Curr. Opin. Neurobiol. 2005, 15, 527–535. [Google Scholar] [CrossRef]
- Ponimaskin, E.; Harteneck, C.; Schultz, G.; Schmidt, M.F.G. A Cysteine-11 to Serine Mutant of Gα12 Impairs Activation through the Thrombin Receptor. FEBS Lett. 1998, 429, 370–374. [Google Scholar] [CrossRef] [Green Version]
- Ponimaskin, E.; Behn, H.; Adarichev, V.; Voyno-Yasenetskaya, T.A.; Offermanns, S.; Schmidt, M.F.G. Acylation of Gα13 Is Important for Its Interaction with Thrombin Receptor, Transforming Activity and Actin Stress Fiber Formation. FEBS Lett. 2000, 478, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Tsutsumi, R.; Fukata, Y.; Noritake, J.; Iwanaga, T.; Perez, F.; Fukata, M. Identification of G Protein α Subunit-Palmitoylating Enzyme. Mol. Cell. Biol. 2009, 29, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleuss, C.; Krause, E. Gαs Is Palmitoylated at the N-Terminal Glycine. EMBO J. 2003, 22, 826–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juhnn, Y.S.; Jones, T.L.Z.; Spiegel, A.M. Amino- and Carboxy-Terminal Deletion Mutants of Gsα Are Localized to the Particulate Fraction of Transfected COS Cells. J. Cell Biol. 1992, 119, 523–530. [Google Scholar] [CrossRef]
- Mumby, S.M.; Kleuss, C.; Gilman, A.G. Receptor Regulation of G-Protein Palmitoylation. Proc. Natl. Acad. Sci. USA 1994, 91, 2800–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedegaertner, P.B.; Bourne, H.R. Activation and Depalmitoylation of Gsα. Cell 1994, 77, 1063–1070. [Google Scholar] [CrossRef]
- Resh, M.D. Fatty Acylation of Proteins: New Insights into Membrane Targeting of Myristoylated and Palmitoylated Proteins. BBA Mol. Cell Res. 1999, 1451, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wensel, T.G. N-Myristoylation of the Rod Outer Segment G Protein, Transducin, in Cultured Retinas. J. Biol. Chem. 1992, 267, 23197–23201. [Google Scholar] [CrossRef]
- Farazi, T.A.; Waksman, G.; Gordon, J.I. The Biology and Enzymology of Protein N-Myristoylation. J. Biol. Chem. 2001, 276, 39501–39505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer-Stroh, S.; Eisenhaber, B.; Eisenhaber, F. N-Terminal N-Myristoylation of Proteins: Refinement of the Sequence Motif and Its Taxon-Specific Differences. J. Mol. Biol. 2002, 317, 523–540. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y.; et al. CHARMM-GUI Input Generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM Simulations Using the CHARMM36 Additive Force Field. J. Chem. Theory Comput. 2016, 12, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.L.Z.; Simonds, W.F.; Merendino, J.J., Jr.; Brann, M.R.; Spiegel, A.M. Myristoylation of an Inhibitory GTP-Binding Protein α Subunit Is Essential for Its Membrane Attachment. Proc. Natl. Acad. Sci. USA 1990, 87, 568–572. [Google Scholar] [CrossRef] [Green Version]
- Degtyarev, M.Y.; Spiegel, A.M.; Jones, T.L.Z. Palmitoylation of a G Protein α(i) Subunit Requires Membrane Localization Not Myristoylation. J. Biol. Chem. 1994, 269, 30898–30903. [Google Scholar] [CrossRef]
- Morales, J.; Fishburn, C.S.; Wilson, P.T.; Bourne, H.R. Plasma Membrane Localization of Gα(z) Requires Two Signals. Mol. Biol. Cell 1998, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.T.; Bourne, H.R. Fatty Acylation of α(z). Effects of Palmitoylation and Myristoylation on α(z) Signaling. J. Biol. Chem. 1995, 270, 9667–9675. [Google Scholar] [CrossRef] [Green Version]
- Gallego, C.; Gupta, S.K.; Winitz, S.I.M.; Eisfelder, B.J.; Johnson, G.L. Myristoylation of the Gαi2 Polypeptide, a G Protein α Subunit, Is Required for Its Signaling and Transformation Functions. Proc. Natl. Acad. Sci. USA 1992, 89, 9695–9699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taussig, R.; Lniguez-LIuhi, J.A.; Gilman, A.G. Inhibition of Adenylyl Cyclase by Giα. Science 1993, 261, 218–221. [Google Scholar] [CrossRef]
- Preininger, A.M.; Van Eps, N.; Nan-Jun, Y.; Medkova, M.; Hubbell, W.L.; Hamm, H.E. The Myristoylated Amino Terminus of Gαi1 Plays a Critical Role in the Structure and Function of Gαi1 Subunits in Solution. Biochemistry 2003, 42, 7931–7941. [Google Scholar] [CrossRef]
- van Keulen, S.C.; Rothlisberger, U. Effect of N-Terminal Myristoylation on the Active Conformation of Gαi1-GTP. Biochemistry 2017, 56, 271–280. [Google Scholar] [CrossRef] [Green Version]
- van Keulen, S.C.; Rothlisberger, U. Exploring the Inhibition Mechanism of Adenylyl Cyclase Type 5 by N-Terminal Myristoylated Gαi1. PLoS Comput. Biol. 2017, 13, 1–18. [Google Scholar] [CrossRef]
- Zhang, F.L.; Casey, P.J. Protein Prenylation: Molecular Mechanisms and Functional Consequences. Annu. Rev. Biochem. 1996, 65, 241–269. [Google Scholar] [CrossRef]
- Higgins, J.B.; Casey, P.J. The Role of Prenylation in G-Protein Assembly and Function. Cell. Signal. 1996, 8, 433–437. [Google Scholar] [CrossRef]
- Rocks, O.; Peyker, A.; Kahms, M.; Verveer, P.J.; Koerner, C.; Lumbierres, M.; Kuhlmann, J.; Waldmann, H.; Wittinghofer, A.; Bastiaens, P.I.H. An Acylation Cycle Regulates Localization and Activity of Palmitoylated Ras Isoforms. Science 2005, 307, 1746–1752. [Google Scholar] [CrossRef]
- Shahinian, S.; Silvius, J.R. Doubly-Lipid-Modified Protein Sequence Motifs Exhibit Long-Lived Anchorage to Lipid Bilayer Membranes. Biochemistry 1995, 34, 3813–3822. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.B.; Casey, P.J. In Vitro Processing of Recombinant G Protein γ Subunits. Requirements for Assembly of an Active Βγ Complex. J. Biol. Chem. 1994, 269, 9067–9073. [Google Scholar] [CrossRef]
- Wedegaertner, P.B.; Wilson, P.T.; Bourne, H.R. Lipid Modifications of Trimeric G Proteins. J. Biol. Chem. 1995, 270, 503–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melkonian, K.A.; Ostermeyer, A.G.; Chen, J.Z.; Roth, M.G.; Brown, D.A. Role of Lipid Modifications in Targeting Proteins to Detergent-Resistant Membrane Rafts. J. Biol. Chem. 1999, 274, 3910–3917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weise, K.; Triola, G.; Brunsveld, L.; Waldmann, H.; Winter, R. Influence of the Lipidation Motif on the Partitioning and Association of N-Ras in Model Membrane Subdomains. J. Am. Chem. Soc. 2009, 131, 49352–49359. [Google Scholar] [CrossRef]
- Crouthamel, M.; Thiyagarajan, M.M.; Evanko, D.S.; Wedegaertner, P.B. N-Terminal Polybasic Motifs Are Required for Plasma Membrane Localization of Gαs and Gαq. Cell. Signal. 2008, 20, 1900–1910. [Google Scholar] [CrossRef] [Green Version]
- Pedone, K.H.; Hepler, J.R. The Importance of N-Terminal Polycysteine and Polybasic Sequences for G14α and G16α Palmitoylation, Plasma Membrane Localization, and Signaling Function. J. Biol. Chem. 2007, 282, 25199–25212. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.L. The Polybasic Region of Ras and Rho Family Small GTPases: A Regulator of Protein Interactions and Membrane Association and a Site of Nuclear Localization Signal Sequences. Cell. Signal. 2003, 15, 1071–1080. [Google Scholar] [CrossRef]
- Do Heo, W.; Inoue, T.; Park, W.S.; Kim, M.L.; Park, B.O.; Wandless, T.J.; Meyer, T. PI(3,4,5)P3 and PI(4,5)P2 Lipids Target Proteins with Polybasic Clusters to the Plasma Membrane. Science 2006, 314, 1458–1462. [Google Scholar] [CrossRef] [Green Version]
- Prakash, P.; Hancock, J.F.; Gorfe, A.A. Three Distinct Regions of CRaf Kinase Domain Interact with Membrane. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A. Membrane Recognition by Phospholipid-Binding Domains. Nat. Rev. Mol. Cell Biol. 2008, 9, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Shi, X.; Guo, X.; Li, H.; Xu, C. Ionic Protein-Lipid Interaction at the Plasma Membrane: What Can the Charge Do? Trends Biochem. Sci. 2014, 39, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Marrari, Y.; Crouthamel, M.; Irannejad, R.; Wedegaertner, P.B. Assembly and Trafficking of Heterotrimeric G Proteins. Biochemistry 2007, 46, 7665–7677. [Google Scholar] [CrossRef] [Green Version]
- Lounsburys, K.M.; Caseyot, P.J.; Brass, L.F.; Manning, D.R. Phosphorylation of Gz in Human Platelets. J. Biol. Chem. 1991, 266, 22051–22056. [Google Scholar] [CrossRef]
- Kosloff, M.; Alexov, E.; Arshavsky, V.Y.; Honig, B. Electrostatic and Lipid Anchor Contributions to the Interaction of Transducin with Membranes. J. Biol. Chem. 2008, 283, 31197–31207. [Google Scholar] [CrossRef] [Green Version]
- Borders, C.L.; Broadwater, J.A.; Bekeny, P.A.; Salmon, J.E.; Lee, A.N.N.S.; Eldridge, A.M.; Pett, V.B. A Structural Role for Arginine in Proteins: Multiple Hydrogen Bonds to Backbone Carbonyl Oxygens. Protein Sci. 1994, 3, 541–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donald, J.E.; Kulp, D.W.; Degrado, W.F. Salt Bridges: Geometrically Specific, Designable Interactions. Proteins 2011, 79, 898–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hristova, K.; William, C. Wimley A Look at Arginine in Membranes. J. Membr. Biol. 2011, 23, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, P.; Gorfe, A.A. Membrane Orientation Dynamics of Lipid-Modified Small GTPases. Small GTPases 2017, 8, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Crocker, E.; McLaughlin, S.; Smith, S.O. Binding of Peptides with Basic and Aromatic Residues to Bilayer Membranes. Phenylalanine in the Myristoylated Alanine-Rich C Kinase Substrate Effector Domain Penetrates into the Hydrophobic Core of the Bilayer. J. Biol. Chem. 2003, 278, 21459–21466. [Google Scholar] [CrossRef] [Green Version]
- Krahn, M.P. Phospholipids of the Plasma Membrane – Regulators or Consequence of Cell Polarity? Front. Cell Dev. Biol. 2020, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Di Paolo, G.; De Camilli, P. Phosphoinositides in Cell Regulation and Membrane Dynamics. Nature 2006, 443, 651–657. [Google Scholar] [CrossRef]
- Leventis, P.A.; Grinstein, S. The Distribution and Function of Phosphatidylserine in Cellular Membranes. Annu. Rev. Biophys. 2010, 39, 407–427. [Google Scholar] [CrossRef] [PubMed]
- Okeley, N.M.; Gelb, M.H. A Designed Probe for Acidic Phospholipids Reveals the Unique Enriched Anionic Character of the Cytosolic Face of the Mammalian Plasma Membrane. J. Biol. Chem. 2004, 279, 21833–21840. [Google Scholar] [CrossRef] [Green Version]
- Tate, M.W.; Eikenberry, E.F.; Turner, D.C.; Shyamsunder, E.; Gruner, S.M. Nonbilayer Phases of Membrane Lipids. Chem. Phys. Lipids 1991, 57, 147–164. [Google Scholar] [CrossRef]
- Koldsø, H.; Shorthouse, D.; Helie, J.; Sansom, M.S.P. Lipid Clustering Correlates with Membrane Curvature as Revealed by Molecular Simulations of Complex Lipid Bilayers. PLoS Comput. Biol. 2014, 10, e1003911. [Google Scholar] [CrossRef] [PubMed]
- Cebecauer, M.; Amaro, M.; Jurkiewicz, P.; Sarmento, M.J.; Săachl, R.; Cwiklik, L.; Hof, M. Membrane Lipid Nanodomains. Chem. Rev. 2018, 118, 11259–11297. [Google Scholar] [CrossRef] [PubMed]
- Zegarlińska, J.; Piaścik, M.; Sikorski, A.F.; Czogalla, A. Phosphatidic Acid - A Simple Phospholipid with Multiple Faces. Acta Biochim. Pol. 2018, 65, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Zhukovsky, M.A.; Filograna, A.; Luini, A.; Corda, D.; Valente, C. Phosphatidic Acid in Membrane Rearrangements. FEBS Lett. 2019, 593, 2428–2451. [Google Scholar] [CrossRef] [Green Version]
- Tanguy, E.; Kassas, N.; Vitale, N. Protein–Phospholipid Interaction Motifs: A Focus on Phosphatidic Acid. Biomol. NMR Assign. 2018, 8, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kooijman, E.E.; Tieleman, D.P.; Testerink, C.; Munnik, T.; Rijkers, D.T.S.; Burger, K.N.J.; De Kruijff, B. An Electrostatic/Hydrogen Bond Switch as the Basis for the Specific Interaction of Phosphatidic Acid with Proteins. J. Biol. Chem. 2007, 282, 11356–11364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiselev, V.Y.; Marenduzzo, D.; Goryachev, A.B. Lateral Dynamics of Proteins with Polybasic Domain on Anionic Membranes: A Dynamic Monte-Carlo Study. Biophys. J. 2011, 100, 1261–1270. [Google Scholar] [CrossRef] [Green Version]
- Golebiewska, U.; Gambhir, A.; Hangyas-Mihalyne, G.; Zaitseva, I.; Radler, J.; Mclaughlin, S. Membrane-Bound Basic Peptides Sequester Multivalent (PIP2), but Not Monovalent (PS), Acidic Lipids. Biophys. J. 2006, 91, 588–599. [Google Scholar] [CrossRef] [Green Version]
- Gambhir, A.; Hangyas-Mihalyne, G.; Zaitseva, I.; Cafiso, D.S.; Wang, J.; Murray, D.; Pentyala, S.N.; Smith, S.O.; Mclaughlin, S. Electrostatic Sequestration of PIP2 on Phospholipid Membranes by Basic/Aromatic Regions of Proteins. Biophys. J. 2004, 86, 2188–2207. [Google Scholar] [CrossRef] [Green Version]
- Balla, T. Phosphoinositides: Tiny Lipids With Giant Impact on Cell Regulation. Physiol. Rev. 2013, 93, 1019–1137. [Google Scholar] [CrossRef]
- Yeung, T.; Terebiznik, M.; Yu, L.; Silvius, J.R.; Abidi, W.M.; Philips, M.; Levine, T.; Kapus, A.; Grinstein, S. Receptor Activation Alters Inner Surface Potential During Phagocytosis. Science 2006, 313, 347–351. [Google Scholar] [CrossRef]
- Lee, A.G. How Lipids Affect the Activities of Integral Membrane Proteins. Biochim. Et Biophys. Acta Biomembr. 2004, 1666, 62–87. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Prakash, P.; Liang, H.; Cho, K.; Gorfe, A.A.; Hancock, J.F. Lipid-Sorting Specificity Encoded in K-Ras Article Lipid-Sorting Specificity Encoded in K-Ras Membrane Anchor Regulates Signal Output. Cell 2017, 168, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Jastrzebska, B.; Debinski, A.; Filipek, S.; Palczewski, K. Role of Membrane Integrity on G Protein-Coupled Receptors: Rhodopsin Stability and Function. Prog. Lipid Res. 2011, 50, 267–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aveldaño de Caldironi, M.; Giusto, N.; Bazàn, N. Polyunsaturated Fatty Acids of the Retina. Prog. Lipid Res. 1981, 20, 49–57. [Google Scholar] [CrossRef]
- Hessel, E.; Heck, M.; Mu, P.; Herrmann, A.; Peter, K. Signal Transduction in the Visual Cascade Involves Specific Lipid-Protein Interactions. J. Biol. Chem. 2003, 278, 22853–22860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, D.; Mclaughlin, S.; Honig, B. The Role of Electrostatic Interactions in the Regulation of the Membrane Association of G Protein Βγ Heterodimers. J. Biol. Chem. 2001, 276, 45153–45159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Melia, T.J.; He, F.; Yuan, C.; McGough, A.; Schmid, M.F.; Wensel, T.G. How a G Protein Binds a Membrane. J. Biol. Chem. 2004, 279, 33937–33945. [Google Scholar] [CrossRef] [Green Version]
- Jastrzebska, B.; Goc, A.; Golczak, M.; Palczewski, K. Phospholipids Are Needed for the Proper Formation, Stability, and Function of the Photoactivated Rhodopsin-Transducin Complex. Biochemistry 2009, 48, 5159–5170. [Google Scholar] [CrossRef] [Green Version]
- Elliott, M.H.; Fliesler, S.J.; Ghalayini, A.J. Articles Cholesterol-Dependent Association of Caveolin-1 with the Transducin R Subunit in Bovine Photoreceptor Rod Outer Segments: Disruption by Cyclodextrin and Guanosine 5′-O-(3-Thiotriphosphate). Biochemistry 2003, 42, 7892–7903. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, F.; Saito, N.; Tanimoto, Y.; Okada, K.; Morigaki, K.; Seno, K.; Maekawa, S. Raftophilic Rhodopsin-Clusters Offer Stochastic Platforms for G Protein Signalling in Retinal Discs. Commun. Biol. 2019, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hessel, E.; Mu, P.; Herrmann, A.; Hofmann, K.P. Light-Induced Reorganization of Phospholipids in Rod Disc Membranes. J. Biol. Chem. 2001, 276, 2538–2543. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, R.; López, D.J.; Casas, J.; Lladó, V.; Higuera, M.; Nagy, T.; Barceló, M.; Busquets, X.; Escribá, P. V G Protein-Membrane Interactions I: Gαi1 Myristoyl and Palmitoyl Modifications in Protein-Lipid Interactions and Its Implications in Membrane Microdomain Localization. BBA Mol. Cell Biol. Lipids 2015, 1851, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Casas, J.; Ibarguren, M.; Álvarez, R.; Terés, S.; Lladó, V.; Piotto, S.P.; Concilio, S.; Busquets, X.; López, D.J.; Escribá, P.V. G Protein-Membrane Interactions II: Effect of G Protein-Linked Lipids on Membrane Structure and G Protein-Membrane Interactions. Biochim. Et Biophys. Acta Biomembr. 2017, 1859, 1526–1535. [Google Scholar] [CrossRef]
- Vögler, O.; Casas, J.; Capó, D.; Nagy, T.; Borchert, G.; Martorell, G.; Escribá, P.V. The Gβγ Dimer Drives the Interaction of Heterotrimeric Gi Proteins with Nonlamellar Membrane Structures. J. Biol. Chem. 2004, 279, 36540–36545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mystek, P.; Dutka, P.; Tworzydło, M.; Dziedzicka-Wasylewska, M.; Polit, A. The Role of Cholesterol and Sphingolipids in the Dopamine D1 Receptor and G Proteins Distribution in the Plasma Membrane. Biochim. Et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2016, 1861, 1775–1786. [Google Scholar] [CrossRef]
- Liang, Y.; Khoshouei, M.; Radjainia, M.; Zhang, Y.; Glukhova, A.; Tarrasch, J.; Thal, D.M.; Furness, S.G.B.; Christopoulos, G.; Coudrat, T.; et al. Phase-Plate Cryo-EM Structure of a Class B GPCR–G-Protein Complex. Nature 2017, 546, 118–136. [Google Scholar] [CrossRef]
- Kumar, K.K.; Shalev-benami, M.; Robertson, M.J.; Malhotra, S.V.; Kobilka, B.K.; Kumar, K.K.; Shalev-benami, M.; Robertson, M.J.; Hu, H.; Banister, S.D. Structure of a Signaling Cannabinoid Receptor 1-G Protein Complex. Cell 2019, 176, 448–458.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, H.E.; Zhang, Y.; Hu, H.; Suomivuori, C.; Marie, F.; Kadji, N.; Aoki, J.; Kumar, K.K.; Fonseca, R.; Hilger, D.; et al. Conformational Transitions of a Neurotensin Receptor 1–Gi1 Complex. Nature 2019, 572, 80–101. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.; Pamula, F.; Nehmé, R.; Mühle, J.; Weinert, T.; Flock, T.; Nogly, P.; Edwards, P.C.; Carpenter, B.; Gruhl, T.; et al. Crystal Structure of Rhodopsin in Complex with a Mini-Go Sheds Light on the Principles of G Protein Selectivity. Science 2018, 4, 1–9. [Google Scholar]
- García-Nafría, J.; Lee, Y.; Bai, X.; Carpenter, B.; Tate, C.G. Cryo-EM Structure of the Adenosine A2A Receptor Coupled to an Engineered Heterotrimeric G Protein. eLife 2018, 7, e35946. [Google Scholar] [CrossRef]
- Strohman, M.J.; Maeda, S.; Hilger, D.; Masureel, M.; Du, Y.; Kobilka, B.K. Local Membrane Charge Regulates Β2 Adrenergic Receptor Coupling to Gi. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Yen, H.; Hoi, K.K.; Liko, I.; Hedger, G.; Horrell, M.R.; Wu, D.; Heine, P.; Warne, T.; Lee, Y.; Carpenter, B.; et al. PIP2 Stabilises Active States of GPCRs and Enhances the Selectivity of G-Protein Coupling. Nature 2018, 559, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Gui, M.; Wang, Z.-F.; Gorgulla, C.; Yu, J.J.; Wu, H.; Sun, Z.; Klenk, C.; Merklinger, L.; Morstein, L.; et al. Cryo-EM Structure of an Activated GPCR-G Protein Complex in Lipid Nanodiscs. Nat. Struct. Mol. Biol. 2021, 28, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, S.; Ghirlando, R.; White, J.F.; Gvozdenovic-Jeremic, J.; Northup, J.K.; Grisshammer, R. Modulation of the Interaction between Neurotensin Receptor NTS1 and Gq Protein by Lipid. J. Mol. Biol. 2012, 417, 95–111. [Google Scholar] [CrossRef] [Green Version]
- Heberle, F.A.; Feigenson, G.W. Phase Separation in Lipid Membranes. Cold Spring Harb. Perspect. Biol. 2011, 3, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kiessling, V.; Crane, J.M.; Tamm, L.K. Transbilayer Effects of Raft-Like Lipid Domains in Asymmetric Planar Bilayers Measured by Single Molecule Tracking. Biophys. J. 2006, 91, 3313–3326. [Google Scholar] [CrossRef] [Green Version]
- Goñi, F.M. “Rafts”: A Nickname for Putative Transient Nanodomains. Chem. Phys. Lipids 2019, 218, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Pike, L.J. Rafts Defined: A Report on the Keystone Symposium on Lipid Rafts and Cell Function. J. Lipid Res. 2006, 47, 1597–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Bogaart, G.; Meyenberg, K.; Risselada, H.J.; Amin, H.; Willig, K.I.; Hubrich, B.E.; Dier, M.; Hell, S.W.; Grubmuller, H.; Diederichsen, U.; et al. Membrane Protein Sequestering by Ionic Protein–Lipid Interactions. Nature 2011, 479, 4–7. [Google Scholar] [CrossRef]
- Murray, D.H.; Tamm, L.K. Clustering of Syntaxin-1A in Model Membranes Is Modulated by Phosphatidylinositol. Biochemistry 2009, 48, 4617–4625. [Google Scholar] [CrossRef] [Green Version]
- Mouritsen, O.G.; Bagatolli, L.A. Lipid Domains in Model Membranes: A Brief Historical Perspective. Essays Biochem. 2015, 1–19. [Google Scholar] [CrossRef]
- Devaux, P.F. Dynamic Lipid Asymmetry in Cell Membranes. Biochemistry 1991, 30, 1163–1173. [Google Scholar] [CrossRef]
- Lorent, J.H.; Levental, K.R.; Ganesan, L.; Rivera-Longsworth, G.; Sezgin, E.; Doktorova, M.D.; Lyman, E.; Levental, I. Plasma Membranes Are Asymmetric in Lipid Unsaturation, Packing and Protein Shape. Nat. Chem. Biol. 2020, 16, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Kusumi, A.; Fujiwara, T.K.; Chadda, R.; Xie, M.; Tsunoyama, T.A.; Kalay, Z.; Kasai, R.S.; Suzuki, K.G.N. Dynamic Organizing Principles of the Plasma Membrane That Regulate Signal Transduction: Commemorating the Fortieth Anniversary of Singer and Nicolson’s Fluid-Mosaic Model. Annu. Rev. Cell Dev. Biol. 2012, 28, 215–250. [Google Scholar] [CrossRef] [PubMed]
- Honigmann, A.; Sadeghi, S.; Keller, J.; Hell, S.W.; Eggeling, C.; Vink, R. A Lipid Bound Actin Meshwork Organizes Liquid Phase Separation in Model Membranes. eLife 2014, 3, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moffett, S.; Brown, D.A.; Linder, M.E. Lipid-Dependent Targeting of G Proteins into Rafts. J. Biol. Chem. 2000, 275, 2191–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, P.; Schnitzer, J.E. Segregation of Heterotrimeric G Proteins in Cell Surface Microdomains: Gq Binds Caveolin to Concentrate in Caveolae, Whereas Gi and Gs Target Lipid Rafts by Default. Mol. Biol. Cell 2001, 12, 685–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mystek, P.; Tworzydło, M.; Dziedzicka-Wasylewska, M.; Polit, A. New Insights into the Model of Dopamine D1 Receptor and G-Proteins Interactions. BBA Mol. Cell Res. 2015, 1853, 594–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zidovetzki, R.; Levitan, I. Use of Cyclodextrins to Manipulate Plasma Membrane Cholesterol Content: Evidence, Misconceptions and Control Strategies. BBA Biomembr. 2007, 1768, 1311–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabouridis, P.S.; Janzen, J.; Magee, A.L.; Ley, S.C. Cholesterol Depletion Disrupts Lipid Rafts and Modulates the Activity of Multiple Signaling Pathways in T Lymphocytes. Eur. J. Immunol. 2000, 30, 954–963. [Google Scholar] [CrossRef]
- Lim, W.K.; Myung, C.-S.; Garrison, J.C.; Neubig, R.R. Receptor-G Protein γ Specificity: Γ11 Shows Unique Potency for A1 Adenosine And 5-HT1A Receptors. Biochemistry 2001, 40, 10532–10541. [Google Scholar] [CrossRef] [PubMed]
- Senarath, K.; Payton, J.L.; Kankanamge, D.; Siripurapu, P.; Tennakoon, M.; Karunarathne, A. Gγ Identity Dictates Efficacy of Gβγ Signaling and Macrophage Migration. J. Biol. Chem. 2018, 293, 2974–2989. [Google Scholar] [CrossRef] [Green Version]
- Kisselev, O.; Pronin, A.; Ermolaeva, M.; Gautam, N. Receptor-G Protein Coupling Is Established by a Potential Conformational Switch in the Βγ Complex. Proc. Natl. Acad. Sci. USA 1995, 92, 9102–9106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, L.A.; Schey, K.L.; Cleator, J.H.; Wilcox, M.D.; Dingus, J.; Hildebrandt, J.D. Identification of a Region in G Protein γ Subunits Conserved across Species but Hypervariable among Subunit Isoforms. Protein Sci. 2001, 10, 2548–2555. [Google Scholar] [CrossRef]
- Wen, P.; Mahinthichaichan, P.; Trebesch, N.; Jiang, T.; Zhao, Z.; Shinn, E.; Wang, Y.; Shekhar, M.; Kapoor, K.; Chan, C.K.; et al. Microscopic View of Lipids and Their Diverse Biological Functions. Curr. Opin. Struct. Biol. 2018, 51, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Vereb, G.; Szollosi, J.; Matko, J.; Nagy, P.; Farkas, T.; Vıgh, L.; Matyus, L.; Waldmann, T.A.; Damjanovich, S. Dynamic, yet Structured: The Cell Membrane Three Decades after the Singer–Nicolson Model. Proc. Natl. Acad. Sci. USA 2003, 100, 8053–8058. [Google Scholar] [CrossRef] [Green Version]
- Kaya, A.İ.; Uğur, O.; Altuntaş, O.; Sayar, K.; Onaran, H.O. Long and Short Distance Movements of β(2)-Adrenoceptor in Cell Membrane Assessed by Photoconvertible Fluorescent Protein Dendra2-β(2)-Adrenoceptor Fusion. BBA Mol. Cell Res. 2011, 1813, 1511–1524. [Google Scholar] [CrossRef] [Green Version]
- Senese, N.B.; Rasenick, M.M.; Traynor, J.R. The Role of G-Proteins and G-Protein Regulating Proteins in Depressive Disorders. Front. Pharmacol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Donati, R.J.; Dwivedi, Y.; Roberts, R.C.; Conley, R.R.; Pandey, G.N.; Rasenick, M.M. Postmortem Brain Tissue of Depressed Suicides Reveals Increased Gsα Localization in Lipid Raft Domains Where It Is Less Likely to Activate Adenylyl Cyclase. J. Neurosci. 2008, 28, 3042–3050. [Google Scholar] [CrossRef] [Green Version]
- Donati, R.J.; Rasenick, M.M. Chronic Antidepressant Treatment Prevents Accumulation of Gsα in Cholesterol-Rich, Cytoskeletal-Associated, Plasma Membrane Domains (Lipid Rafts). Neuropsychopharmacology 2005, 30, 1238–1245. [Google Scholar] [CrossRef] [Green Version]
- Hicks, D.A.; Nalivaeva, N.N.; Turner, A.J. Lipid Rafts and Alzheimer’s Disease: Protein-Lipid Interactions and Perturbation of Signaling. Front. Physiol. 2012, 3, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukiw, W.J. Alzheimer’s Disease (AD) as a Disorder of the Plasma Membrane. Front. Physiol. 2013, 4, 15–17. [Google Scholar] [CrossRef] [Green Version]
- Fabiani, C.; Antollini, S.S.; Guido, M.E.; Eckert, G.P.; Stephan, A. Alzheimer’s Disease as a Membrane Disorder: Spatial Cross-Talk Among Beta-Amyloid Peptides, Nicotinic Acetylcholine Receptors and Lipid Rafts. Front. Cell. Neurosci. 2019, 13, 1–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simons, K.; Ehehalt, R. Cholesterol, Lipid Rafts, and Disease. J. Clin. Investig. 2002, 110, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Di Scala, C.; Troadec, J.; Lelièvre, C.; Garmy, N.; Fantini, J.; Chahinian, H. Mechanism of Cholesterol-assisted Oligomeric Channel Formation by a Short Alzheimer Β-amyloid Peptide. J. Neurochem. 2014, 128, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Thathiah, A.; Strooper, B. De The Role of G Protein-Coupled Receptors in the Pathology of Alzheimer’s Disease. Nat. Rev. Neurosci. 2011, 12, 73–87. [Google Scholar] [CrossRef]
- Zhao, J.; Deng, Y.; Jiang, Z.; Qing, H. G Protein-Coupled Receptors (GPCRs) in Alzheimer’s Disease: A Focus on BACE1 Related GPCRs. Front. Aging Neurosci. 2016, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, E.; Kim, I.; Akther, M.; Choi, D. Importance of GPCR-Mediated Microglial Activation in Alzheimer’s Disease. Front. Cell. Neurosci. 2018, 12, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Sushma; Mondal, A.C. Role of GPCR Signaling and Calcium Dysregulation in Alzheimer’s Disease. Mol. Cell. Neurosci. 2019, 101, 1–11. [Google Scholar] [CrossRef]
- Escribá, P.V.; Sánchez-dominguez, J.M.; Alemany, R.; Perona, J.S.; Ruiz-gutie, V. Alteration of Lipids, G Proteins, and PKC in Cell Membranes of Elderly Hypertensives. Hypertension 2003, 41, 176–182. [Google Scholar] [CrossRef] [Green Version]
- Perona, J.S.; Vo, O.; Sa, J.M.; Escriba, P.V.; Ruiz-Gutierrez, V. Consumption of Virgin Olive Oil Influences Membrane Lipid Composition and Regulates Intracellular Signaling in Elderly Adults With Type 2 Diabetes Mellitus. J. Gerantology Biol. Sci. 2007, 62, 256–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 0, 1–41. [Google Scholar] [CrossRef]
- Szlasa, W.; Zendran, I.; Zalesińska, A.; Tarek, M.; Kulbacka, J. Lipid Composition of the Cancer Cell Membrane. J. Bioenerg. Biomembr. 2020, 52, 321–342. [Google Scholar] [CrossRef] [PubMed]
- Zalba, S.; Hagen, T.L.M. Cell Membrane Modulation as Adjuvant in Cancer Therapy. Cancer Treat. Rev. 2017, 52, 48–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escriba, P.V.; Sastre, M.; Garcia-Sevilla, J.A. Disruption of Cellular Signaling Pathways by Daunomycin through Destabilization of Nonlamellar Membrane Structures. Proc. Natl. Acad. Sci. USA 1995, 92, 7595–7599. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Family | Member | Distribution | Major Signaling Role |
|---|---|---|---|
| Gαs | Gαs(S) Gαs(L) | ubiquitous | adenylyl cyclase (+) |
| Gαs(XL) | neuroendocrine cells | ||
| Gαolf | olfactory sensory neurons, striatum | ||
| Gαi/o | Gαi1, Gαi2, Gαi3 | ubiquitous | adenylyl cyclase (−) |
| Gαo1, Gαo2 | neurons, heart, neuroendocrine cells | ||
| Gαz | rod and cone cells of the eye | ||
| Gαt1, Gαt2 | taste cells | phosphodiesterase (+) | |
| Gαg | neurons and platelets, adrenal chromaffin cells, neurosecretory cells | ||
| Gαq/11 | Gαq | ubiquitous | phospholipase C (+) |
| Gα11 | ubiquitous | ||
| Gα14 | kidney, lung, liver, spleen | ||
| Gα15 | hematopoietic cells | ||
| Gα16 | hematopoietic cells | ||
| Gα12/13 | Gα12 | ubiquitous | Rho GTPases (+) |
| Gα13 | ubiquitous |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polit, A.; Mystek, P.; Błasiak, E. Every Detail Matters. That Is, How the Interaction between Gα Proteins and Membrane Affects Their Function. Membranes 2021, 11, 222. https://doi.org/10.3390/membranes11030222
Polit A, Mystek P, Błasiak E. Every Detail Matters. That Is, How the Interaction between Gα Proteins and Membrane Affects Their Function. Membranes. 2021; 11(3):222. https://doi.org/10.3390/membranes11030222
Chicago/Turabian StylePolit, Agnieszka, Paweł Mystek, and Ewa Błasiak. 2021. "Every Detail Matters. That Is, How the Interaction between Gα Proteins and Membrane Affects Their Function" Membranes 11, no. 3: 222. https://doi.org/10.3390/membranes11030222
APA StylePolit, A., Mystek, P., & Błasiak, E. (2021). Every Detail Matters. That Is, How the Interaction between Gα Proteins and Membrane Affects Their Function. Membranes, 11(3), 222. https://doi.org/10.3390/membranes11030222