1. Introduction
The standardized
Kt/
V, which indicates the ratio of blood purification amount by dialysis to the amount of body fluid for the patient, is widely used today as an index to evaluate dialysis adequacy [
1,
2]. The standardized
Kt/
V consists of three important dialysis parameters, namely,
K: clearance (mL/min),
t: dialysis time (min), and
V: total volume of body fluid (m
3) for the patient. This standardized
Kt/
V was proposed by Gotch and Sargent [
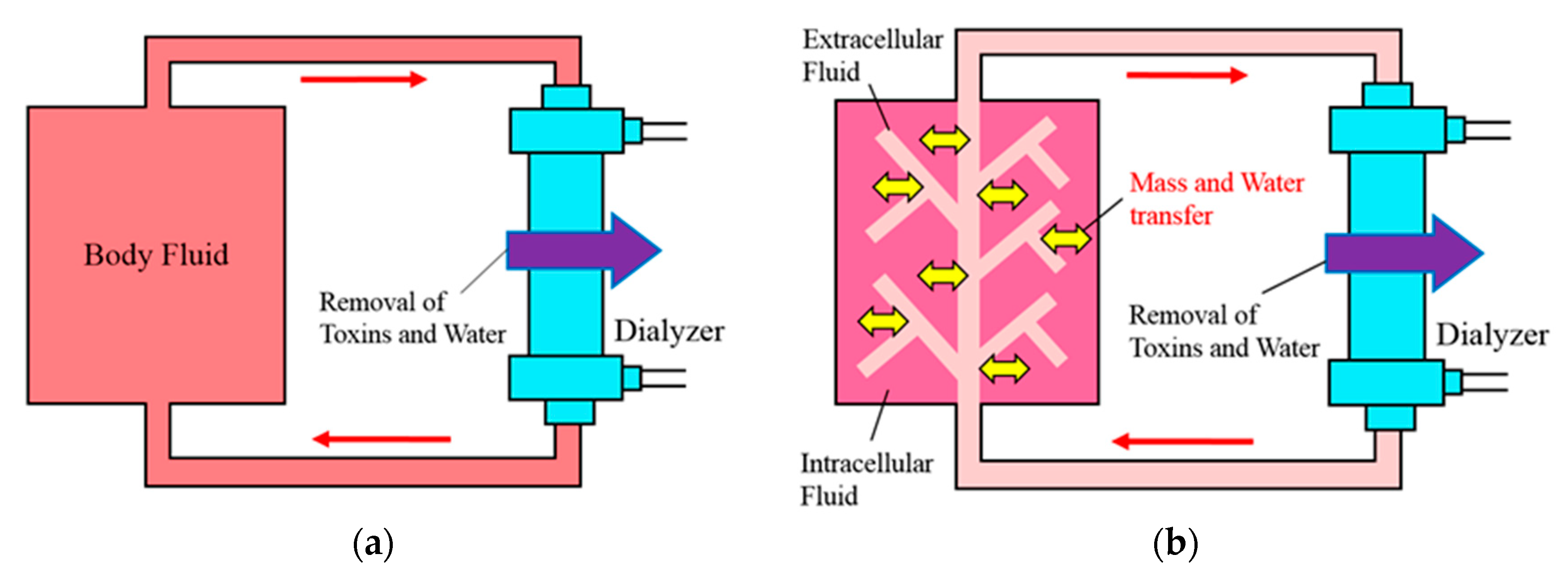
1] for predicting the blood toxin concentration during dialysis. They assumed that a whole body is a contaminated pool (single compartment) and a loop for blood purification is connected during dialysis treatment (see
Figure 1a). Their model was simple and acceptable to doctors who treated with dialysis at the time, however, the model did not consider the effect of the volume decrease by ultrafiltration and toxin production in the body. Subsequently, several improved models have been proposed, so that the Daugirdas formula is recommended by the K/DOQI guidelines; on the other hand, the Shinzato formula is recommended by the Japanese Society for Dialysis Therapy [
3,
4,
5]. However, their models are also proposed based on single-compartment assumption.
Pedrini et al. [
6] raised the question of the validity of the models derived by the single-compartment assumption. Subsequently, some models taking into account the non-uniformity of metabolic substances in the body have been proposed [
7,
8]. There is the two-compartment model [
8] in them, which was proposed based on an assumption that the body consists of intracellular and extracellular fluids (see
Figure 1b). In this model, a loop for blood purification is connected at an extracellular fluid, and mass transfer takes place between intracellular and extracellular fluids. This model was a more realistic model because blood purification in dialysis is performed via blood. On the other hand, this model was more complicated than the single-compartment model. Some expression formulas [
8,
9,
10,
11,
12,
13,
14] have been proposed as two-compartment models so far, and none were mathematically modeled from mass transfer in the body. Furthermore, in almost these studies, numerical simulation was used to evaluate the blood toxin concentration. Similarly, there is peritoneal dialysis as a treatment method for purifying blood. In peritoneal dialysis, the mass transfer takes place between multiple compartments such as blood, peritoneum, and dialysate. Several multi-compartment models have been proposed so far [
15,
16,
17,
18], which have been evaluated both numerically and analytically. Generically, numerical simulation requires considerable calculation time and is less accurate than an analytical solution. In contrast, an analytical solution is an algebraic expression, so that we can obtain the blood toxin concentration in any time period and any conditions when substituting dialysis conditions. Furthermore, the analytical solution can be implemented into the dialysis device, and the doctor may perform hemodialysis by predicting the blood toxin concentration.
In the present study, we attempted to derive the mathematical model that predicts blood toxin concentrations during and after dialysis, which has both accuracy and practicality. First, the new two-compartment model was mathematically derived by adapting volume-averaging theory to the mass transfer around peripheral tissues. Volume-averaging theory is a kind of multi-scale theory that derives a macroscopic governing equation from a microscopic governing equation by coupling heat or mass transport phenomena in numerous small elements existing in a large space [
19,
20,
21,
22,
23]. Subsequently, analytical solutions for a two-compartment model were derived as a practical model for predicting blood toxin concentrations during and after dialysis. The post dialysis analytical solution for predicting the post dialysis rebound phenomenon was derived from the same procedure. The validity of the proposed solutions was examined by comparing with clinical data and numerical solutions. Furthermore, the proposed analytical solutions were compared with Gotch formula [
1] and Shinzato formula [
4], which are the single-compartment models.
2. Theory and Analytical Solutions
We shall consider the mass transport phenomena around peripheral tissues, as shown in
Figure 2. According to Pedrini et al. [
6], the toxin concentration differs mainly inside and outside the cell, because mass transfer resistance at the cell membrane is higher than that of the capillary wall. Therefore, in the present study, we focus on the toxin concentrations in extracellular and intracellular fluid phases and consider the mass transfer between theses phases. Note that active transport via ion channels is not considered in the present study.
In volume-averaging theory, the intrinsic volume average of a certain variable
ϕ is defined as:
where
Vphase is the volume occupied by intracellular and extracellular fluids. When applying volume averaging theory, considering the control volume scale [
22], the control volume scale
V in
Figure 2 should be much greater than the microscopic characteristic length (cell length), but at the same time, should be much smaller than the macroscopic characteristic length (human length). Furthermore, a variable is decomposed into its intrinsic average and the spatial deviation therefrom:
In volume-averaging theory, the following spatial average relationships are used (for details, see [
19,
20,
21,
22,
23]):
where
εphase = Vphase/V is the local volume fraction of each space, and
Aint is the interfacial area between each phase within the control volume. In the present study,
ni is defined as a unit vector pointing outward from the extracellular fluid to the intracellular fluid.
The microscopic species mass transfer equations for the individual phases can be written as follows:
where
D is the diffusion coefficient, and the species concentration
c can be treated as an independent component in metabolic substances, such as urea nitrogen or creatinine. Furthermore,
s indicates the toxin production rate per unit volume, which is assumed as a constant value and it would be zero extracellular fluid phases. The
xj indicates Cartesian coordinate, which has
x,
y and
z. The index
j is a subscript of Einstein notation, which represents from 1 to 3.
The macroscopic governing equations for the extracellular and intracellular phases can be expressed using the above spatial average relationships:
where the subscripts
ex and
in indicate extracellular fluid and intracellular fluid phases, respectively. Note that the velocity in the intracellular fluid vanished because the integral of the velocity vector in the cell (i.e., closed space) is zero. In contrast, the last term on the right-hand side, which expresses the mass transport with flow through cell membranes (i.e., plasma refilling [
24]), cannot be eliminated because of the product of the unit vector. In this analysis, the assumption that the perfusion by plasma refilling via the cell membrane has a uniform velocity vector in the normal direction of the cell membrane was applied. Based on a previous paper [
25], the perfusion rate for plasma refilling per unit volume
νpl (s
−1) is defined as:
and the third terms in Equations (5) and (6) can be modeled as:
On the other hand, the second term on the right-hand side in Equations (5) and (6), which describes the diffusive mass transport between extracellular and intracellular fluid phases, may be modeled via Newton’s cooling law as:
where
asf is the specific surface area, and
h is the overall mass transfer coefficient between the extracellular fluid and intracellular fluid phases. Furthermore, the second and third bracketed terms on the right-hand sides of Equations (5) and (6), which describe the effects of tortuosity and mechanical dispersion, respectively, are negligibly small around peripheral tissues. Thus, the macroscopic species mass transfer equations for extracellular fluid and intracellular fluid phases can be expressed as:
We shall derive the two-compartment models from Equations (10) and (11). Commonly, the measured toxin concentration from a vein during and after dialysis is considered to be the average concentration, rather than the concentration in an individual organ. Therefore, the above macroscopic mass transfer (Equations (10) and (11)) around capillaries is integrated over extracellular and intracellular fluid volumes
Vex and
Vin, respectively, with the assumption that the volume of the solute is small enough that it does not affect each volume
Vex and
Vin. The lumped parameter models of Equations (12) and (13) can be led as follows:
where
C(
t) is a function of time
t and indicates the average concentration over the intrinsic volume of each phase in the body. Therefore, the terms on the left-hand side in Equations (12) and (13) indicate the amount of toxin change per unit of time in the extracellular and intracellular fluid volumes, respectively. On the other hand, the first terms on the right-hand side in Equations (12) and (13) indicate the mass transport that occurs between the extracellular and intracellular fluid phases, where
A is the total surface area between the extracellular and intracellular fluid phases. Furthermore,
h is the overall mass transfer coefficient. The second terms on the right-hand side in Equations (12) and (13) indicate the amount of toxin transported with plasma refilling. Here, the perfusion rate of plasma refilling
ωpl (mL/s) was assumed to be a positive value because plasma mainly flows out from the intracellular fluid phase to the extracellular fluid phase during dialysis. Moreover, the third term in Equation (13) indicates the amount of toxin produced in the body,
S is the value of
s integrated over the intracellular fluid volume
Vin. Therefore, Equations (12) and (13) are a general two-compartment model obtained from mass transport phenomena in the body.
We shall derive analytical solution of Equations (12) and (13). In Equation (12),
B.C. indicates the mass increase–decrease rate associated with the boundary condition. In the present study, firstly, we shall attempt to analyze the time development toxin concentration in the body during dialysis. The boundary condition during dialysis is given using the clearance
K:
This equation indicates the toxin removal rate per unit of time in a dialyzer, which is taken into account in the toxin removal rates by diffusion and ultrafiltration in the dialyzer [
26,
27].
As shown in Equations (12) and (13), each volume is also a function of time because the intracellular and extracellular volumes change during dialysis. The volumes of the extracellular and intracellular fluids are changed due to ultrafiltration and plasma refilling over the dialysis treatment time as follows:
where
ωf indicates the ultrafiltration flow rate in the dialyzer, for which it is assumed that the ultrafiltration flow rate
ωf is always greater than the perfusion rate of plasma refilling
ωpl. This is because, in dialysis, the plasma feeding occurs due to the decrease in blood volume with ultrafiltration. By substituting Equations (15) and (16) into Equations (12) and (13) and then removing
V from the time derivatives according to the differential formula rule, the following mass balance equations can be obtained:
In the present study, we attempt to derive analytical solutions of the mass balance equations, i.e., Equations (17) and (18). Based on the papers [
10,
11,
12,
13,
28], the extracellular fluid volume and intracellular fluid volume are assumed to maintain a constant ratio:
The term
ωpl can be estimated from Equations (15), (16) and (19) as follows:
From Equation (20), ωpl is approximately constant under dialysis treatment conditions. Therefore, in the present study, ωpl was treated as a constant.
During dialysis (0 ≦
t ≦
T), the extracellular fluid volume and the intracellular fluid volume change over time. Therefore, for obtaining the general solutions of Equations (17) and (18), we essentially use Formulas (19) and (20). In the present study, we introduce a variable transformation. A new parameter,
t*, that satisfies the following relationship is introduced:
Thus, from Equations (15) and (21), the following relationship is obtained:
By integrating this equation under the initial conditions (at
t = 0:
t* = 0), a relational expression between
t and
t* can be obtained as follows:
On the other hand, the concentrations for the extracellular and intracellular fluid phases, which are functions
f of
t*, are defined as:
Differentiating
and
with respect to
t*, the following relations can be obtained:
In other words, the differential values of
and
correspond to the left-hand sides of Equations (17) and (18), respectively. The right-hand sides of Equations (17) and (18) are similarly rewritten in terms of
and
, as follows:
Thus, the number of governing equations, i.e., Equations (28) and (29), and the number of unknowns became the same by introducing the defining equations, i.e., Equations (21), (24) and (25), which means that we can solve the analytical solutions. Therefore, in the present study, the analytical solutions are derived from Equations (28) and (29).
From simultaneous Equations (19) and (20), the second-order ordinary differential equation can be obtained as follows:
where
are constants. Equation (30) can be solved, and the general solution of
is given as follows:
where
and
and
are integration constants. In Equation (30), the analytical solution associated with the multiple root is omitted to enhance the understanding of the content for the reader, since it was confirmed that there is probably no multiple root within the range in dialysis treatment by an exhaustive investigation. Furthermore, by substituting Equation (32) into Equation (29), the general solution of
is obtained as follows:
By exploiting Equation (23) through Equation (25), the analytical solutions for the toxin concentrations in the extracellular and intracellular fluids during dialysis are derived as follows.
Analytical solution during dialysis:
where the integration constants
and
can be determined by the initial conditions of the extracellular and intracellular fluid phases. In the present study, we adopted the following initial conditions:
Thus,
and
were estimated as follows:
Next, we shall derive analytical solution during the process in which intracellular and extracellular toxin concentrations re-equilibrate after dialysis (i.e., rebound process). After dialysis (
t ≧
T), the blood purification in the dialyzer was completed, so
K = 0 mL/min and
= 0 mL/min. Accordingly,
is considered to be 0 mL/min. If the patient is not eating or drinking, then the extracellular and intracellular fluid volumes after dialysis are constant. Thus, the general solutions of Equations (17) and (18) can be obtained without introducing variable conversion. From Equations (17) and (18), the second-order ordinary differential equation for evaluating the rebound process can be obtained as follows:
Thus, the general solution of
is given as follows:
where
where
and
are integration constants, and by substituting Equation (41) into Equation (18), the general solution of
is given as follows:
where integration constants
and
can be estimated based on the initial conditions for the extracellular and intracellular fluid phases. In the present study, the toxin concentrations in the extracellular and intracellular fluids at the end of dialysis
and
were adopted as the initial conditions, which were calculated from the analytical solutions, i.e., Equations (35) and (36):
As a result of this initial condition,
and
were estimated as follows:
Combining the analytical solutions during dialysis (i.e., Equations (35) and (36) with Equations (38) and (39)) and the analytical solutions after dialysis (i.e., Equations (41) and (43) with Equations (45) and (46)), it is possible to predict the time development of the toxin concentration in the body during and after dialysis.








{kind=link}
{kind=link}
{kind=link}
{kind=link}