
DNA-Based Molecular Engineering of the Cell Membrane


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
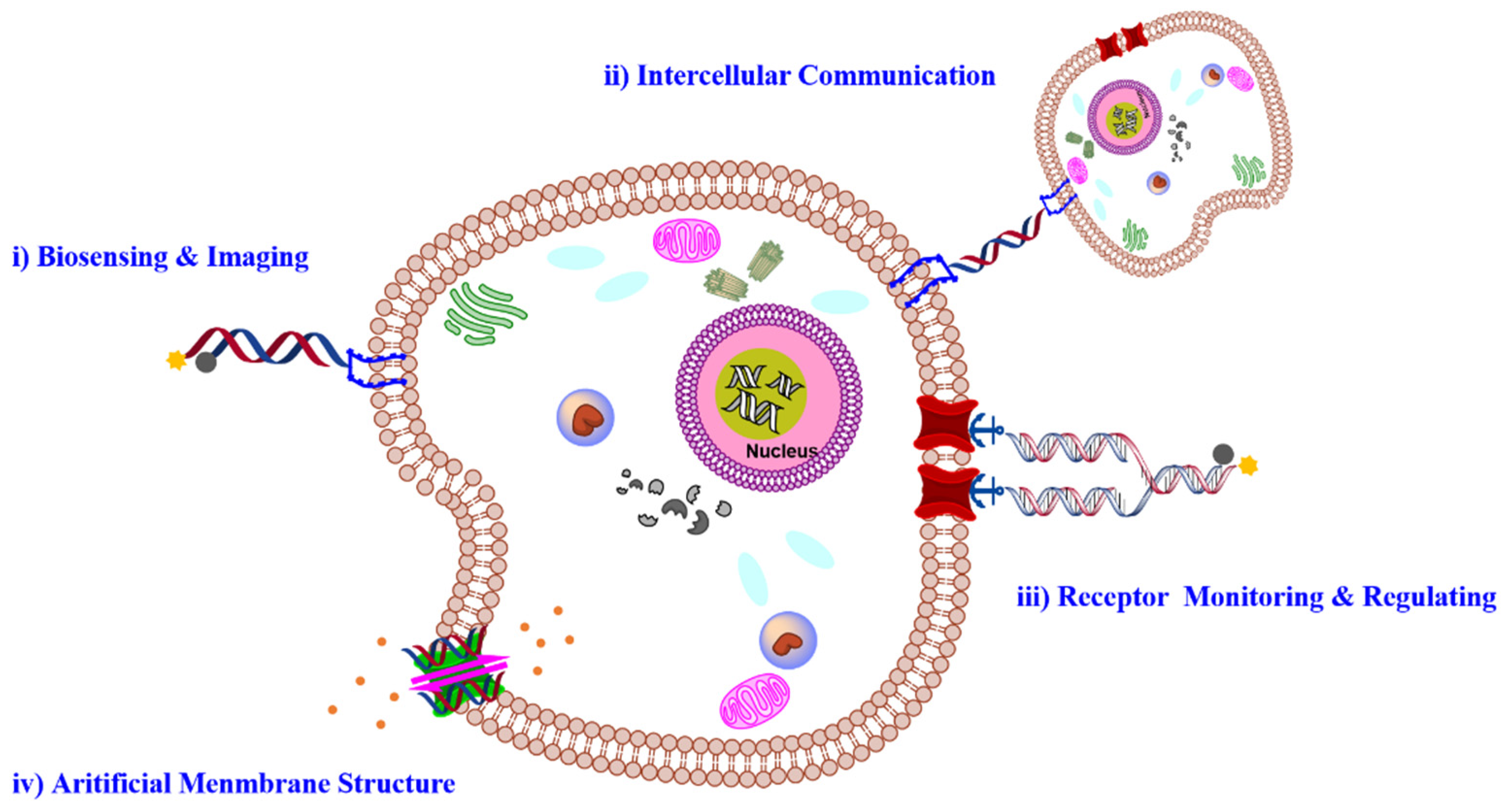
2. Membrane Functionalization Strategies
3. DNA-Based Molecular Engineering on Cell Membrane
3.1. FNAs for Cell Surface Sensing and Imaging
3.2. DNA-Based Intercellular Communication
3.3. DNA-Based Receptor Monitoring and Regulating
3.4. DNA-Based Biomimetic Membrane Constructs
4. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Munro, S. Lipid rafts: Elusive or illusive? Cell 2003, 115, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Grecco, H.E.; Schmick, M.; Bastiaens, P.I. Signaling from the living plasma membrane. Cell 2011, 144, 897–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blume-Jensen, P.; Hunter, T. Oncogenic kinase signalling. Nature 2001, 411, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Schlessinger, J. Cell signaling by receptor tyrosine kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [Green Version]
- Fischbach, M.A.; Bluestone, J.A.; Lim, W.A. Cell-based therapeutics: The next pillar of medicine. Sci. Transl. Med 2013, 5, 179ps177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, D.; Pulsipher, A.; Luo, W.; Mak, H.; Yousaf, M.N. Engineering Cell Surfaces via Liposome Fusion. Bioconj. Chem. 2011, 22, 2423–2433. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Wang, H.-H.; Xie, S.; Wang, M.; Nie, Z. Engineering Cell-Surface Receptors with DNA Nanotechnology for Cell Manipulation. ChemBioChem 2020, 21, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Miao, Y.; Han, D. DNA Nanotechnology on Live Cell Membranes. Chem. Res. Chin. Univ. 2020, 36, 203–210. [Google Scholar] [CrossRef]
- Feng, L.; Li, J.; Sun, J.; Wang, L.; Fan, C.; Shen, J. Recent Advances of DNA Nanostructure-Based Cell Membrane Engineering. Adv. Healthc. Mater. 2021, 10, 2001718. [Google Scholar] [CrossRef] [PubMed]
- Schoenit, A.; Cavalcanti-Adam, E.A.; Göpfrich, K. Functionalization of Cellular Membranes with DNA Nanotechnology. Trends Biotechnol. 2021, 9, 1208–1220. [Google Scholar] [CrossRef]
- Micura, R.; Höbartner, C. Fundamental studies of functional nucleic acids: Aptamers, riboswitches, ribozymes and DNAzymes. Chem. Soc. Rev. 2020, 49, 7331–7353. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cao, Z.; Lu, Y. Functional Nucleic Acid Sensors. Chem. Rev. 2009, 109, 1948–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Liu, X.; Willner, I. DNA switches: From principles to applications. Angew. Chem. Int. Ed. 2015, 54, 1098–1129. [Google Scholar] [CrossRef]
- Kruger, K.; Grabowski, P.J.; Zaug, A.J.; Sands, J.; Gottschling, D.E.; Cech, T.R. Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA intervening sequence of tetrahymena. Cell 1982, 31, 147–157. [Google Scholar] [CrossRef]
- Breaker, R.R.; Joyce, G.F. A DNA enzyme that cleaves RNA. Chem. Biol. 1994, 1, 223–229. [Google Scholar] [CrossRef]
- Liu, J.; Lu, Y. A DNAzyme Catalytic Beacon Sensor for Paramagnetic Cu2+ Ions in Aqueous Solution with High Sensitivity and Selectivity. J. Am. Chem. Soc. 2007, 129, 9838–9839. [Google Scholar] [CrossRef]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Dunn, M.R.; Jimenez, R.M.; Chaput, J.C. Analysis of aptamer discovery and technology. Nat. Rev. Chem. 2017, 1, 0076. [Google Scholar] [CrossRef]
- Vázquez-González, M.; Willner, I. Aptamer-Functionalized Hybrid Nanostructures for Sensing, Drug Delivery, Catalysis and Mechanical Applications. Int. J. Mol. Sci. 2021, 22, 1803. [Google Scholar] [CrossRef]
- Seeman, N.C.; Sleiman, H.F. DNA nanotechnology. Nat. Rev. Mater. 2017, 3, 17068. [Google Scholar] [CrossRef]
- Shih, W.M.; Quispe, J.D.; Joyce, G.F. A 1.7-kilobase single-stranded DNA that folds into a nanoscale octahedron. Nature 2004, 427, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Pal, S.; Nangreave, J.; Deng, Z.; Liu, Y.; Yan, H. DNA origami with complex curvatures in three-dimensional space. Science 2011, 332, 342–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, W.-C.; Willner, I. Synthesis and Applications of Stimuli-Responsive DNA-Based Nano- and Micro-Sized Capsules. Adv. Funct. Mater 2017, 27, 1702732. [Google Scholar] [CrossRef]
- Seeman, N.C. Nucleic acid junctions and lattices. J. Theor. Biol. 1982, 99, 237–247. [Google Scholar] [CrossRef]
- Chen, J.H.; Seeman, N.C. Synthesis from DNA of a molecule with the connectivity of a cube. Nature 1991, 350, 631–633. [Google Scholar] [CrossRef]
- Rothemund, P.W.K. Folding DNA to create nanoscale shapes and patterns. Nature 2006, 440, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Ke, Y.; Ong, L.L.; Shih, W.M.; Yin, P. Three-dimensional structures self-assembled from DNA bricks. Science 2012, 338, 1177–1183. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Lv, H.; Li, Q.; Li, J.; Zhang, X.; Shi, J.; Wang, L.; Fan, C. Implementing digital computing with DNA-based switching circuits. Nat. Commun. 2020, 11, 121. [Google Scholar] [CrossRef] [Green Version]
- Xiaoqing, L.; Chun-Hua, L.; Itamar, W. Switchable Reconfiguration of Nucleic Acid Nanostructures by Stimuli-Responsive DNA Machines. Acc. Chem. Res. 2014, 47, 1673–1680. [Google Scholar]
- Liang, Y.; Shan, W.; Zhixin, Z.; Itamar, W. Nucleic Acid Based Constitutional Dynamic Networks: From Basic Principles to Applications. J. Am. Chem. Soc. 2020, 142, 52. [Google Scholar]
- Ramezani, H.; Dietz, H. Building machines with DNA molecules. Nat. Rev. Genet. 2020, 21, 5–26. [Google Scholar] [CrossRef] [PubMed]
- Jianbang, W.; Liang, Y.; Ziyuan, L.; Junji, Z.; He, T.; Itamar, W. Active generation of nanoholes in DNA origami scaffolds for programmed catalysis in nanocavities. Nat. Commun. 2019, 10, 4963. [Google Scholar]
- Langecker, M.; Arnaut, V.; List, J.; Simmel, F.C. DNA Nanostructures Interacting with Lipid Bilayer Membranes. Acc. Chem. Res. 2014, 47, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Tian, Q.; Bagheri, Y.; You, M. Lipid–oligonucleotide conjugates for simple and efficient cell membrane engineering and bioanalysis. Curr. Opin. Biomed. Engin. 2020, 13, 76–83. [Google Scholar] [CrossRef]
- Weber, R.J.; Liang, S.I.; Selden, N.S.; Desai, T.A.; Gartner, Z.J. Efficient Targeting of Fatty-Acid Modified Oligonucleotides to Live Cell Membranes through Stepwise Assembly. Biomacromolecules 2014, 15, 4621–4626. [Google Scholar] [CrossRef] [Green Version]
- Borisenko, G.G.; Zaitseva, M.A.; Chuvilin, A.N.; Pozmogova, G.E. DNA modification of live cell surface. Nucleic Acids Res. 2009, 37, e28. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhu, Z.; Kang, H.; Wu, Y.; Sefan, K.; Tan, W. DNA-based micelles: Synthesis, micellar properties and size-dependent cell permeability. Chemistry 2010, 16, 3791–3797. [Google Scholar] [CrossRef]
- Wang, R.; Wang, C.; Cao, Y.; Zhu, Z.; Yang, C.; Chen, J.; Qing, F.-L.; Tan, W. Trifluoromethylated nucleic acid analogues capable of self-assembly through hydrophobic interactions. Chem. Sci. 2014, 5, 4076–4081. [Google Scholar] [CrossRef]
- Jin, C.; Fu, T.; Wang, R.; Liu, H.; Zou, J.; Zhao, Z.; Ye, M.; Zhang, X.; Tan, W. Fluorinated molecular beacons as functional DNA nanomolecules for cellular imaging. Chem. Sci. 2017, 8, 7082–7086. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, R.; Xie, S.; Wang, R.; Jin, C.; Du, Y.; Fu, T.; Li, J.; Tan, J.; Zhang, L.; Tan, W. Artificial Sandwich Base for Monitoring Single-Nucleobase Changes and Charge-Transfer Rates in DNA. Anal. Chem. 2019, 91, 2074–2078. [Google Scholar] [CrossRef] [PubMed]
- Jie, T.; Hao, L.; Xiaoxiao, H.; Razack, A.; Sitao, X.; Lili, Z.; Mengmeng, Z.; Qiang, L.; Yazhou, L.; Zhijun, S.; et al. Size-Tunable Assemblies Based on Ferrocene-Containing DNA Polymers for Spatially Uniform Penetration. Chem 2019, 5, 1775–1792. [Google Scholar]
- Li, J.; Xun, K.; Pei, K.; Liu, X.; Peng, X.; Du, Y.; Qiu, L.; Tan, W. Cell-Membrane-Anchored DNA Nanoplatform for Programming Cellular Interactions. J. Am. Chem. Soc. 2019, 141, 18013–18020. [Google Scholar] [CrossRef]
- Jin, C.; He, J.; Zou, J.; Xuan, W.; Fu, T.; Wang, R.; Tan, W. Phosphorylated lipid-conjugated oligonucleotide selectively anchors on cell membranes with high alkaline phosphatase expression. Nat. Commun. 2019, 10, 2704. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.; Liu, J.; Guo, L.; Yao, G.; Li, Q.; Wang, L.; Li, J.; Fan, C. Programming Cell–Cell Communications with Engineered Cell Origami Clusters. J. Am. Chem. Soc. 2020, 142, 8800–8808. [Google Scholar] [CrossRef]
- Zhao, W.; Schafer, S.; Choi, J.; Yamanaka, Y.J.; Lombardi, M.L.; Bose, S.; Carlson, A.L.; Phillips, J.A.; Teo, W.; Droujinine, I.A.; et al. Cell-surface sensors for real-time probing of cellular environments. Nat. Nanotechnol. 2011, 6, 524–531. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.; Zhao, N.; Lai, J.; Coyne, J.; Gaddes, E.R.; Wang, Y. Polyvalent Display of Biomolecules on Live Cells. Angew. Chem. Int. Ed. Engl. 2018, 57, 6800–6804. [Google Scholar] [CrossRef]
- Chandra, R.A.; Douglas, E.S.; Mathies, R.A.; Bertozzi, C.R.; Francis, M.B. Programmable Cell Adhesion Encoded by DNA Hybridization. Angew. Chem. Int. Ed. 2006, 45, 896–901. [Google Scholar] [CrossRef]
- Nieves, D.J.; Hilzenrat, G.; Tran, J.; Yang, Z.; MacRae, H.H.; Baker, M.A.B.; Gooding, J.J.; Gaus, K. tagPAINT: Covalent labelling of genetically encoded protein tags for DNA-PAINT imaging. Roy. Soc. Open Sci. 2019, 6, 191268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Li, G. Aptamers against cell surface receptors: Selection, modification and application. Curr. Med. Chem. 2011, 18, 4107–4116. [Google Scholar] [CrossRef] [PubMed]
- Delcanale, P.; Porciani, D.; Pujals, S.; Jurkevich, A.; Chetrusca, A.; Tawiah, K.D.; Burke, D.H.; Albertazzi, L. Aptamers with Tunable Affinity Enable Single-Molecule Tracking and Localization of Membrane Receptors on Living Cancer Cells. Angew. Chem. Int. Ed. 2020, 59, 18546–18555. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Liu, H.; Zhao, Z.; Altman, M.B.; Lopez-Colon, D.; Yang, C.J.; Chang, L.J.; Liu, C.; Tan, W. DNA Aptamer-Mediated Cell Targeting. Angew. Chem. Int. Ed. 2013, 125, 1512–1516. [Google Scholar] [CrossRef] [Green Version]
- Peng, R.; Zheng, X.; Lyu, Y.; Xu, L.; Zhang, X.; Ke, G.; Liu, Q.; You, C.; Huan, S.; Tan, W. Engineering a 3D DNA-Logic Gate Nanomachine for Bispecific Recognition and Computing on Target Cell Surfaces. J. Am. Chem. Soc. 2018, 140, 9793–9796. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, X.; Cui, C.; Pan, X.; Li, X.; Yazd, H.S.; Wu, Q.; Qiu, L.; Li, J.; Tan, W. Aptamer Displacement Reaction from Live-Cell Surfaces and Its Applications. J. Am. Chem. Soc. 2019, 141, 17174–17179. [Google Scholar] [CrossRef]
- Liu, L.; Dou, C.-X.; Liu, J.-W.; Wang, X.-N.; Ying, Z.-M.; Jiang, J.-H. Cell Surface-Anchored DNA Nanomachine for Dynamically Tunable Sensing and Imaging of Extracellular pH. Anal. Chem. 2018, 90, 11198–11202. [Google Scholar] [CrossRef]
- Zeng, S.; Liu, D.; Li, C.; Yu, F.; Fan, L.; Lei, C.; Huang, Y.; Nie, Z.; Yao, S. Cell-Surface-Anchored Ratiometric DNA Tweezer for Real-Time Monitoring of Extracellular and Apoplastic pH. Anal. Chem. 2018, 90, 13459–13466. [Google Scholar] [CrossRef]
- Qiu, L.; Zhang, T.; Jiang, J.; Wu, C.; Zhu, G.; You, M.; Chen, X.; Zhang, L.; Cui, C.; Yu, R.; et al. Cell Membrane-Anchored Biosensors for Real-Time Monitoring of the Cellular Microenvironment. J. Am. Chem. Soc. 2014, 136, 13090–13093. [Google Scholar] [CrossRef] [Green Version]
- Xiong, M.; Zhu, H.; Rong, Q.; Yang, C.; Qiu, L.; Zhang, X.-B.; Tan, W. A membrane-anchored fluorescent probe for detecting K+ in the cell microenvironment. Chem. Commun. 2016, 52, 4679–4682. [Google Scholar] [CrossRef] [Green Version]
- Tokunaga, T.; Namiki, S.; Yamada, K.; Imaishi, T.; Nonaka, H.; Hirose, K.; Sando, S. Cell Surface-Anchored Fluorescent Aptamer Sensor Enables Imaging of Chemical Transmitter Dynamics. J. Am. Chem. Soc. 2012, 134, 9561–9564. [Google Scholar] [CrossRef]
- Feng, G.; Luo, X.; Lu, X.; Xie, S.; Deng, L.; Kang, W.; He, F.; Zhang, J.; Lei, C.; Lin, B. Engineering of nucleic acids and synthetic cofactors as holo sensors for probing signaling molecules in the cellular membrane microenvironment. Angew. Chem. Int. Ed. 2019, 131, 6662–6666. [Google Scholar] [CrossRef]
- You, M.; Lyu, Y.; Han, D.; Qiu, L.; Liu, Q.; Chen, T.; Sam Wu, C.; Peng, L.; Zhang, L.; Bao, G.; et al. DNA probes for monitoring dynamic and transient molecular encounters on live cell membranes. Nat. Nanotechnol. 2017, 12, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liang, H.; Sun, J.; Liu, Y.; Li, J.; Li, J.; Li, J.; Yang, H. Bispecific Aptamer Induced Artificial Protein-Pairing: A Strategy for Selective Inhibition of Receptor Function. J. Am. Chem. Soc. 2019, 141, 12673–12681. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; O’Brien, C.; Mudiyanselage, A.P.K.K.K.; Li, N.; Bagheri, Y.; Wu, R.; Sun, Y.; You, M. Visualizing Intercellular Tensile Forces by DNA-Based Membrane Molecular Probes. J. Am. Chem. Soc. 2017, 139, 18182–18185. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xie, X.; Ma, Z.; Li, Q.; Liu, L.; Hu, X.; Liu, C.; Li, B.; Wang, H.; Chen, N.; et al. Programming Niche Accessibility and In Vitro Stemness with Intercellular DNA Reactions. Adv. Mater. 2018, 30, 1804861. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cai, X.; Wang, L.; Li, J.; Li, Q.; Zuo, X.; Shi, J.; Huang, Q.; Fan, C. DNA orientation-specific adhesion and patterning of living mammalian cells on self-assembled DNA monolayers. Chem. Sci. 2016, 7, 2722–2727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todhunter, M.E.; Jee, N.Y.; Hughes, A.J.; Coyle, M.C.; Cerchiari, A.; Farlow, J.; Garbe, J.C.; LaBarge, M.A.; Desai, T.A.; Gartner, Z.J. Programmed synthesis of three-dimensional tissues. Nat. Methods 2015, 12, 975–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sefah, K.; Shangguan, D.; Xiong, X.; O’Donoghue, M.B.; Tan, W. Development of DNA aptamers using Cell-SELEX. Nat. Protoc. 2010, 5, 1169–1185. [Google Scholar] [CrossRef] [PubMed]
- Shangguan, D.; Li, Y.; Tang, Z.; Cao, Z.C.; Chen, H.W.; Mallikaratchy, P.; Sefah, K.; Yang, C.J.; Tan, W. Aptamers evolved from live cells as effective molecular probes for cancer study. Proc. Natl. Acad. Sci. USA 2006, 103, 11838–11843. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Deng, R.; Sun, Y.; Zhang, L.; Li, J. Reversible control of cell membrane receptor function using DNA nano-spring multivalent ligands. Chem. Sci. 2017, 8, 7098–7105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.; Donovan, M.J.; Jiang, J. Aptamers from Cell-Based Selection for Bioanalytical Applications. Chem. Rev. 2013, 113, 2842–2862. [Google Scholar] [CrossRef] [Green Version]
- You, M.; Chen, Y.; Peng, L.; Han, D.; Yin, B.; Ye, B.; Tan, W. Engineering DNA aptamers for novel analytical and biomedical applications. Chem. Sci. 2011, 2, 1003–1010. [Google Scholar] [CrossRef]
- Schueder, F.; Lara-Gutiérrez, J.; Haas, D.; Beckwith, K.S.; Yin, P.; Ellenberg, J.; Jungmann, R. Super-Resolution Spatial Proximity Detection with Proximity-PAINT. Angew. Chem. Int. Ed. 2021, 60, 716–720. [Google Scholar] [CrossRef]
- Rudchenko, M.; Taylor, S.; Pallavi, P.; Dechkovskaia, A.; Khan, S.; Butler, V.P., Jr.; Rudchenko, S.; Stojanovic, M.N. Autonomous molecular cascades for evaluation of cell surfaces. Nat. Nanotechnol. 2013, 8, 580–586. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, M.; Shi, T.; Yang, S.; Zhang, J.; Wang, H.H.; Nie, Z. A DNA-Mediated Chemically Induced Dimerization (D-CID) Nanodevice for Nongenetic Receptor Engineering To Control Cell Behavior. Angew. Chem. Int. Ed. 2018, 130, 10383–10387. [Google Scholar] [CrossRef]
- Shaw, A.; Lundin, V.; Petrova, E.; Fördős, F.; Benson, E.; Al-Amin, A.; Herland, A.; Blokzijl, A.; Högberg, B.; Teixeira, A.I. Spatial control of membrane receptor function using ligand nanocalipers. Nat. Methods 2014, 11, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.R.; Seifert, A.; Fertig, N.; Howorka, S. A biomimetic DNA-based channel for the ligand-controlled transport of charged molecular cargo across a biological membrane. Nat. Nanotechnol. 2016, 11, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Diederichs, T.; Pugh, G.; Dorey, A.; Xing, Y.; Burns, J.R.; Hung Nguyen, Q.; Tornow, M.; Tampé, R.; Howorka, S. Synthetic protein-conductive membrane nanopores built with DNA. Nat. Commun. 2019, 10, 5018. [Google Scholar] [CrossRef]
- Peng, R.; Xu, L.; Wang, H.; Lyu, Y.; Wang, D.; Bi, C.; Cui, C.; Fan, C.; Liu, Q.; Zhang, X.; et al. DNA-based artificial molecular signaling system that mimics basic elements of reception and response. Nat. Commun. 2020, 11, 978. [Google Scholar] [CrossRef] [Green Version]
- Göpfrich, K.; Li, C.-Y.; Mames, I.; Bhamidimarri, S.P.; Ricci, M.; Yoo, J.; Mames, A.; Ohmann, A.; Winterhalter, M.; Stulz, E.; et al. Ion Channels Made from a Single Membrane-Spanning DNA Duplex. Nano Lett. 2016, 16, 4665–4669. [Google Scholar] [CrossRef]
- Ohmann, A.; Li, C.-Y.; Maffeo, C.; Al Nahas, K.; Baumann, K.N.; Göpfrich, K.; Yoo, J.; Keyser, U.F.; Aksimentiev, A. A synthetic enzyme built from DNA flips 107 lipids per second in biological membranes. Nat. Commun. 2018, 9, 2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Wang, T.; Sun, Y.; Li, C.; Peng, T.; Qiu, L. DNA-Based Molecular Engineering of the Cell Membrane. Membranes 2022, 12, 111. https://doi.org/10.3390/membranes12020111
Li X, Wang T, Sun Y, Li C, Peng T, Qiu L. DNA-Based Molecular Engineering of the Cell Membrane. Membranes. 2022; 12(2):111. https://doi.org/10.3390/membranes12020111
Chicago/Turabian StyleLi, Xiaodong, Tiantian Wang, Yue Sun, Chang Li, Tianhuan Peng, and Liping Qiu. 2022. "DNA-Based Molecular Engineering of the Cell Membrane" Membranes 12, no. 2: 111. https://doi.org/10.3390/membranes12020111