
On the Mechanism of Membrane Permeabilization by Tamoxifen and 4-Hydroxytamoxifen

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Vesicle Preparation for Membrane Permeabilization
2.3. Fluorescence Measurements
2.4. Fourier-Transform Infrared Spectroscopy
2.5. High Sensitivity Differential Scanning Calorimetry
2.6. Molecular Dynamics
3. Results
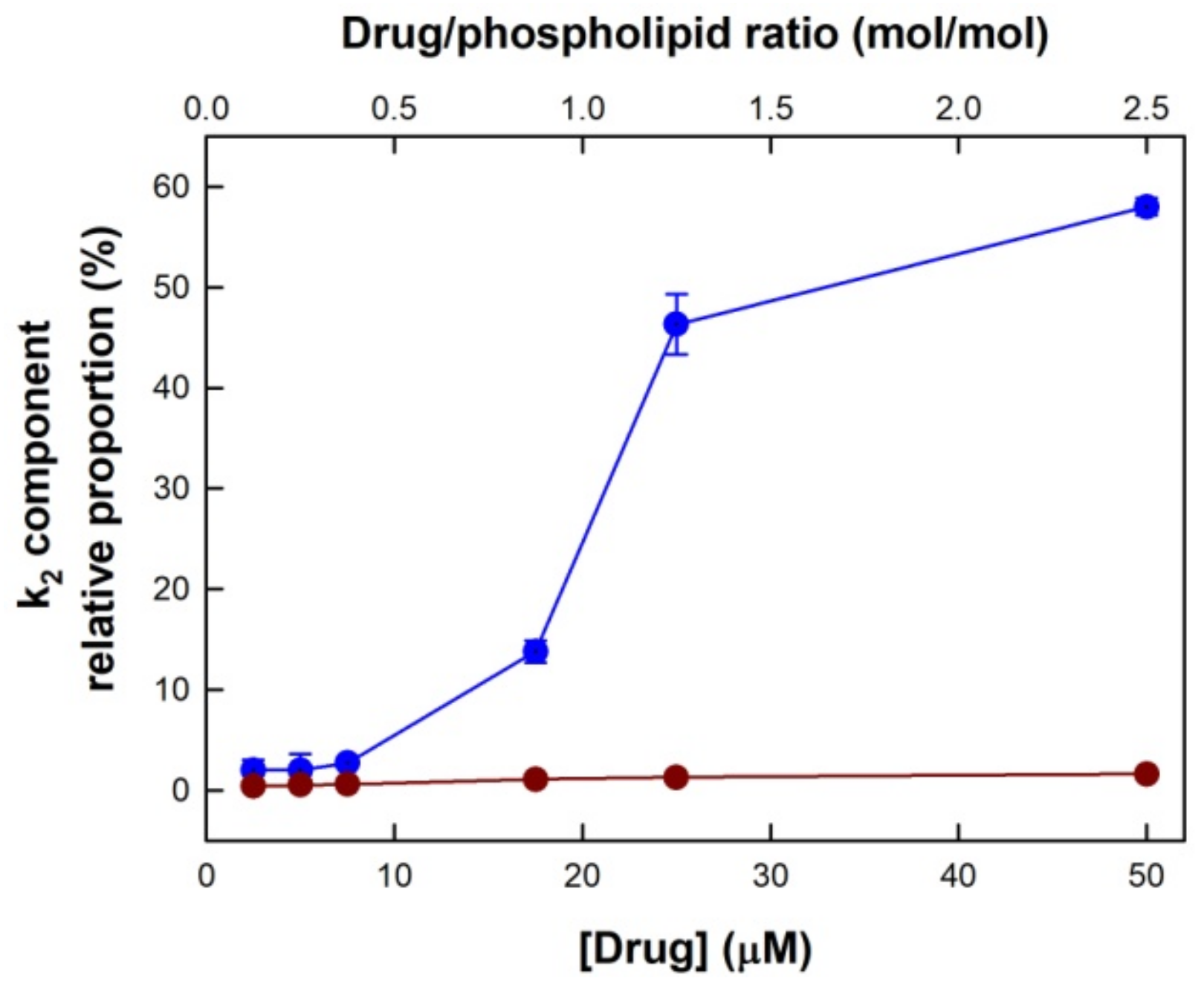
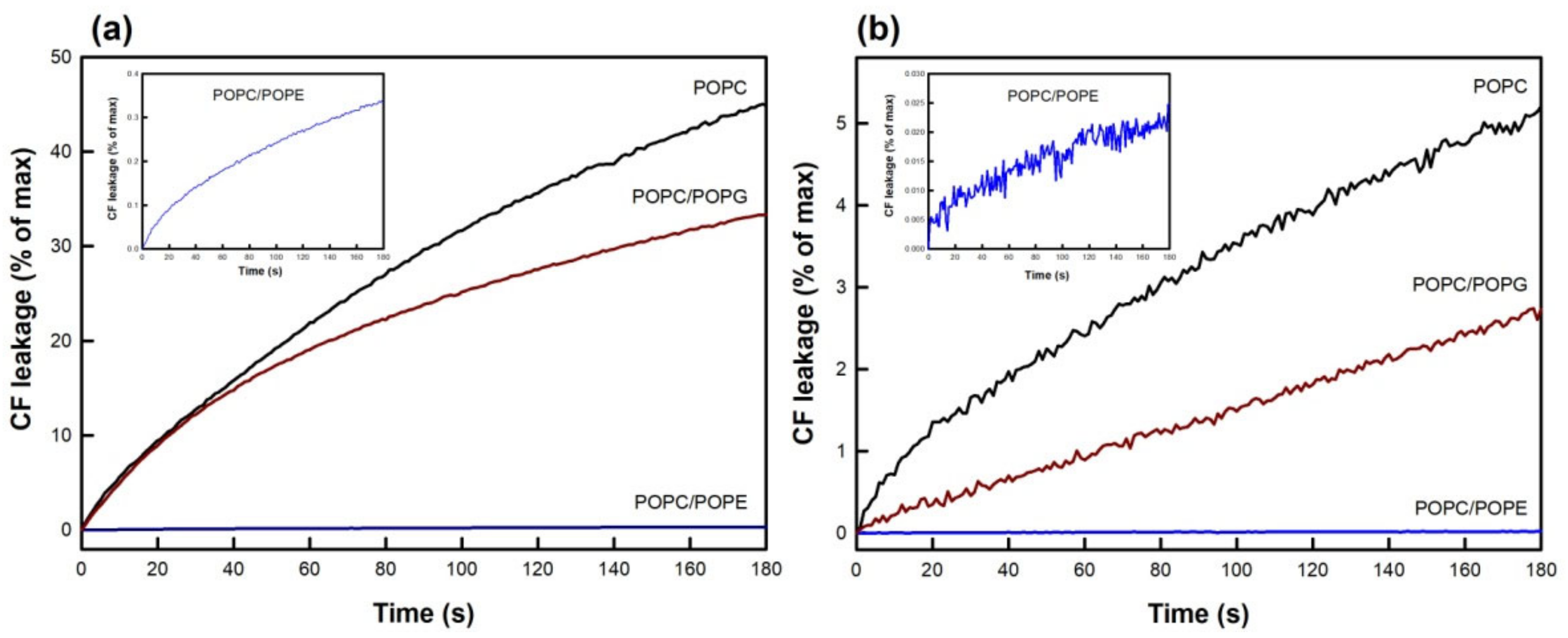
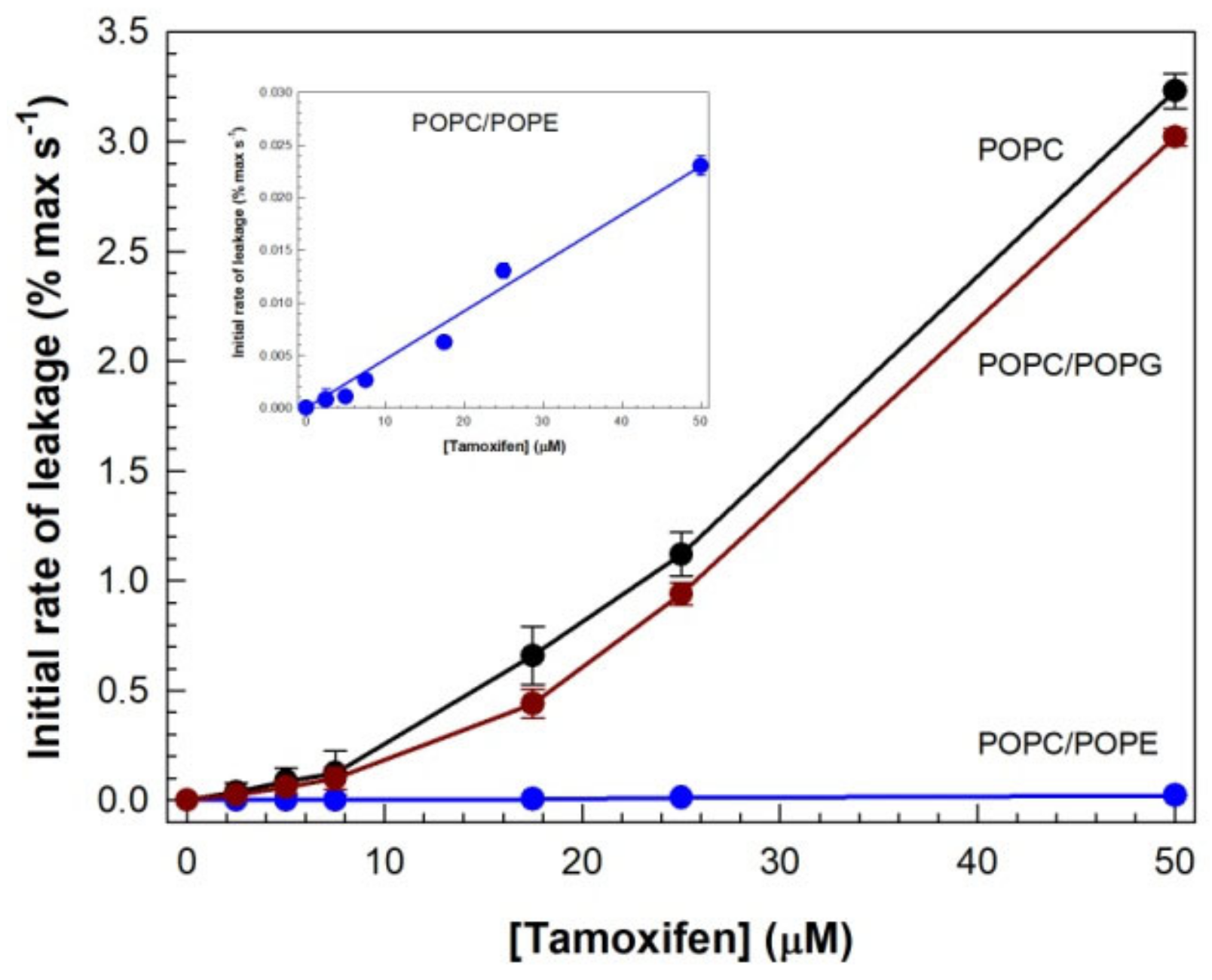
3.1. Drug-Induced Membrane Permeabilization
3.2. Fourier-Transform Infrared Spectroscopy
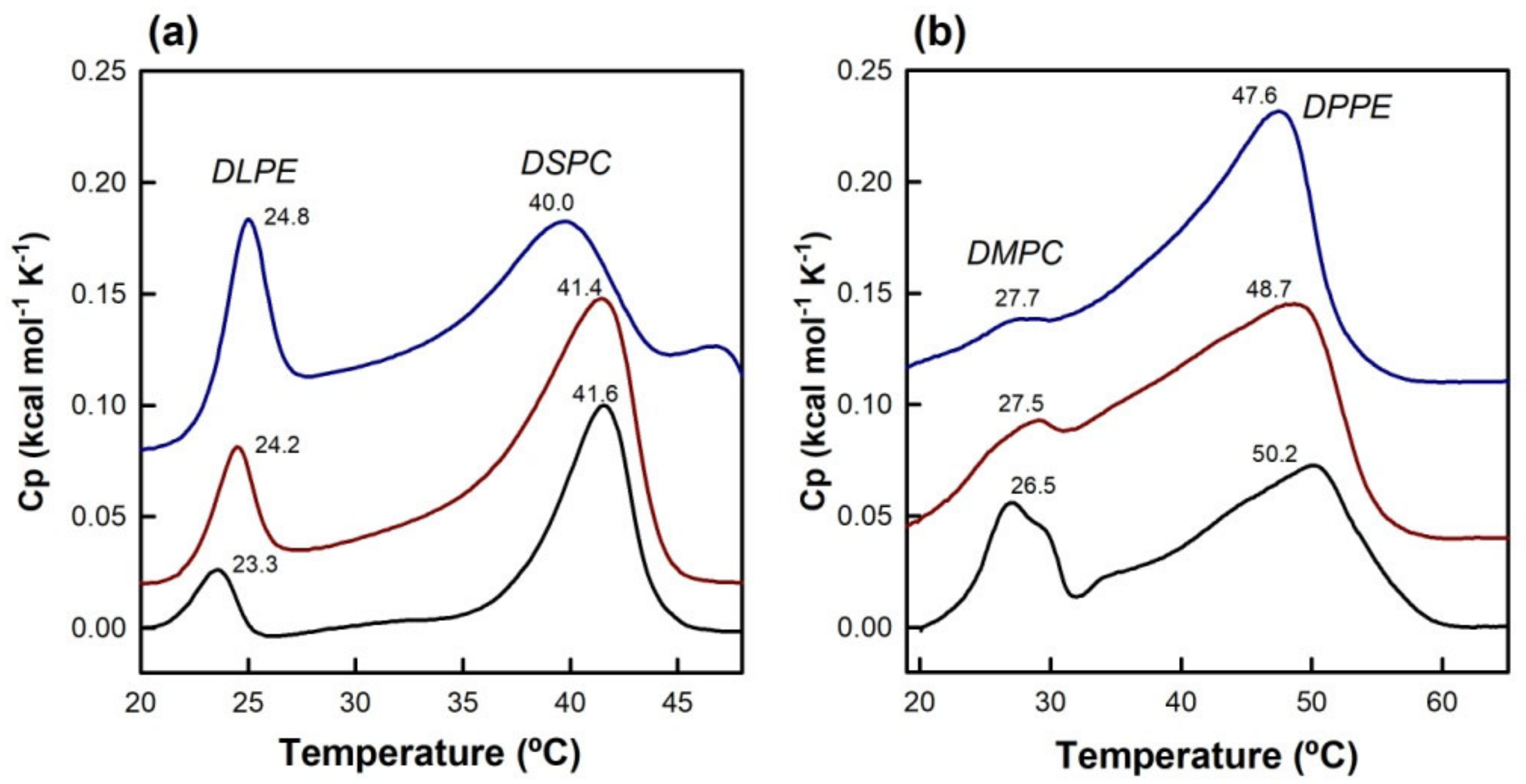
3.3. Differential Scanning Calorimetry
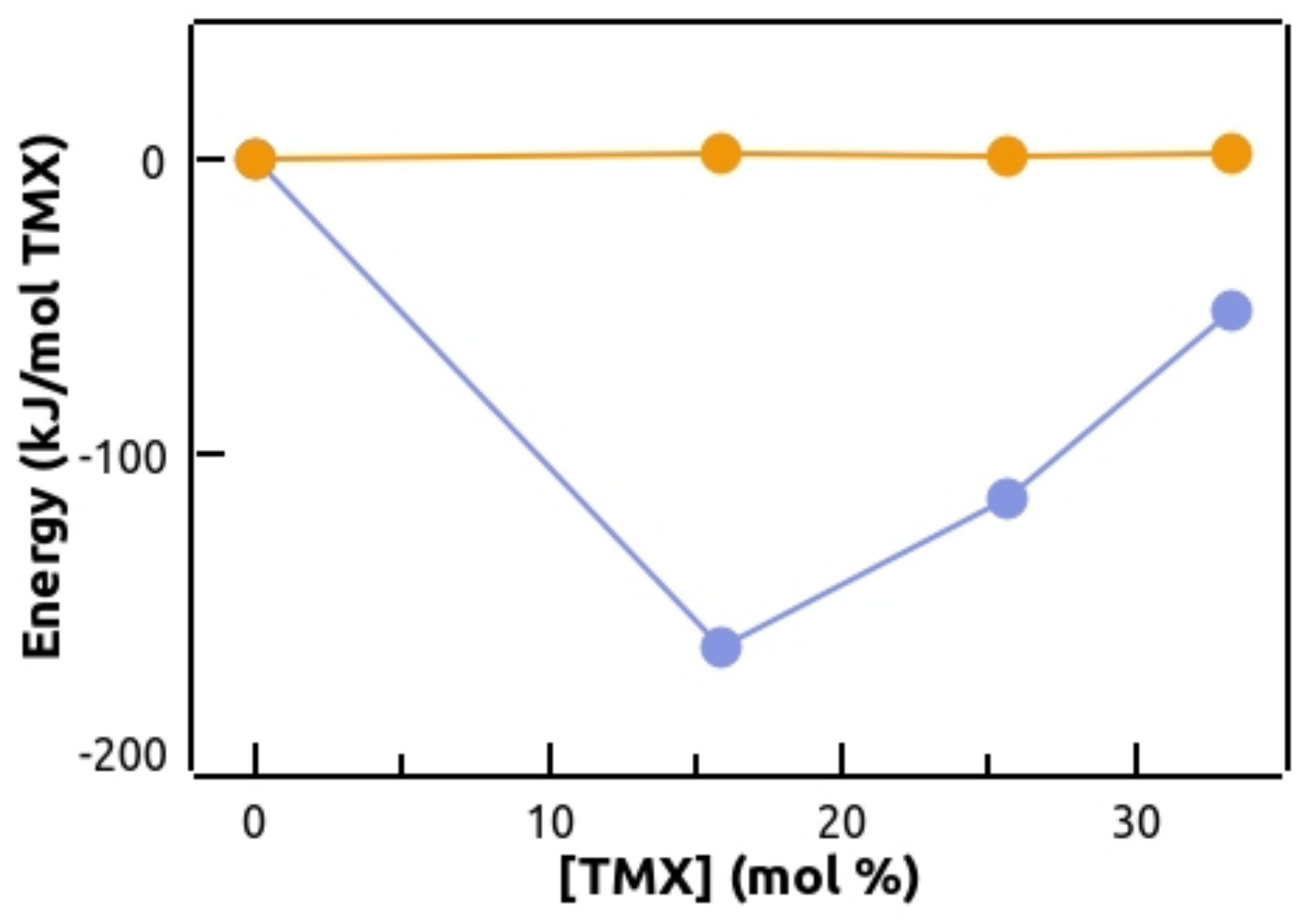
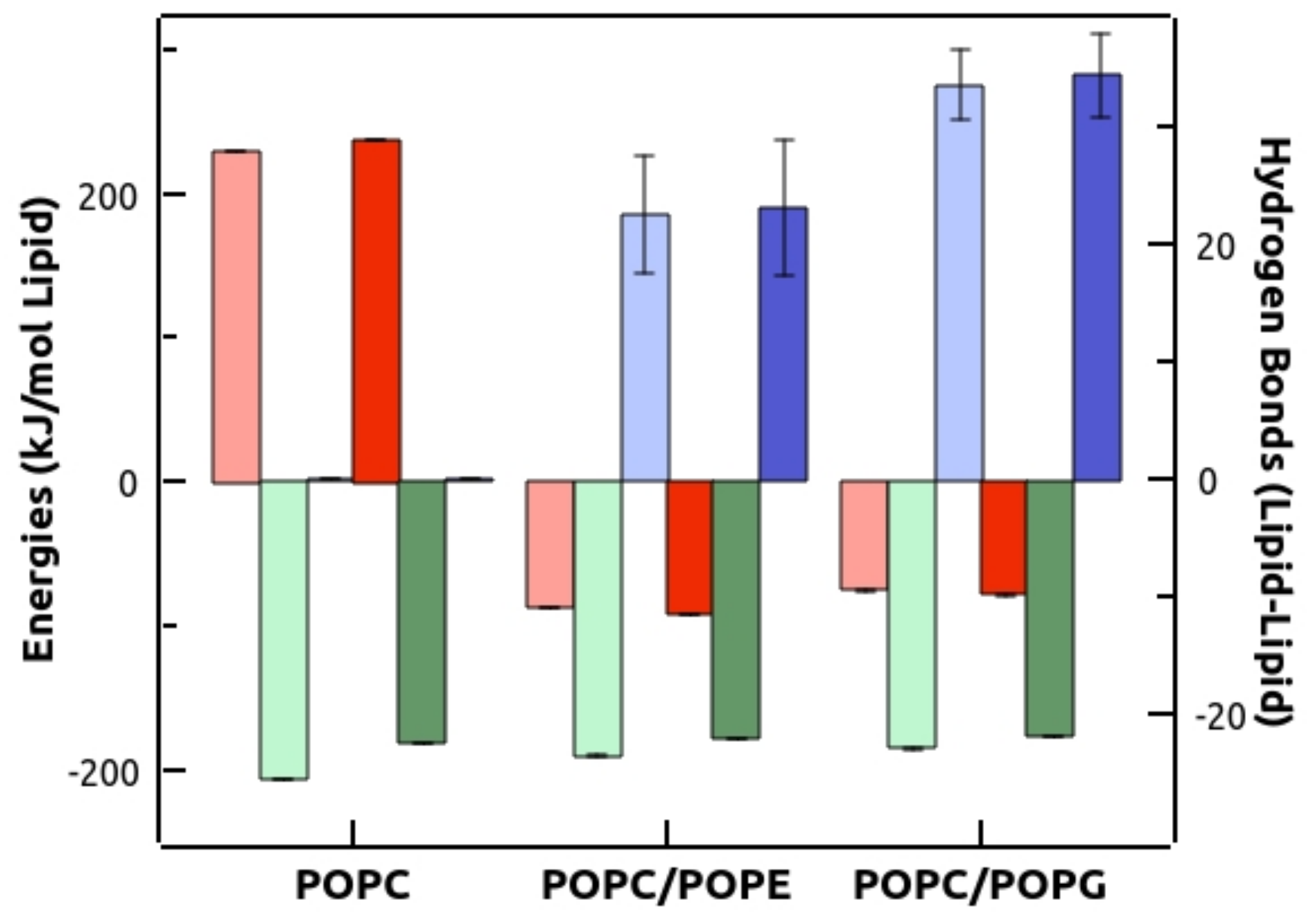
3.4. Molecular Dynamics Simulations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jordan, V.C. Tamoxifen: A most unlikely pioneering medicine. Nat. Rev. Drug Discov. 2003, 2, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Shagufta; Ahmad, I. Tamoxifen a pioneering drug: An update on the therapeutic potential of tamoxifen derivatives. Eur. J. Med. Chem. 2018, 143, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Hayes, T. Pharmacotherapy for male breast cancer. Expert Opin. Pharmacother. 2002, 3, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Hughes-Davies, L.; Caldas, C.; Wishart, G.C. Tamoxifen: The drug that came in from the cold. Br. J. Cancer 2009, 101, 875–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, M.P.; Santos, A.E.; Custódio, J.B. Mitochondria: The gateway for tamoxifen-induced liver injury. Toxicology 2014, 323, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Bilge, D.; Sahin, I.; Kazanci, N.; Severcan, F. Interactions of tamoxifen with distearoyl phosphatidylcholine multilamellar vesicles: FTIR and DSC studies. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2014, 130, 250–256. [Google Scholar] [CrossRef]
- Custódio, J.; Almeida, L.M.; Madeira, V.M. The anticancer drug tamoxifen induces changes in the physical properties of model and native membranes. Biochim. Biophys. Acta (BBA) -Biomembr. 1993, 1150, 123–129. [Google Scholar] [CrossRef]
- Dicko, A.; Morissette, M.; Ben Ameur, S.; Pézolet, M.; Di Paolo, T. Effect of estradiol and tamoxifen on brain membranes: Investigation by infrared and fluorescence spectroscopy. Brain Res. Bull. 1999, 49, 401–405. [Google Scholar] [CrossRef]
- Engelke, M.; Bojarski, P.; Bloß, R.; Diehl, H. Tamoxifen perturbs lipid bilayer order and permeability: Comparison of DSC, fluorescence anisotropy, Laurdan generalized polarization and carboxyfluorescein leakage studies. Biophys. Chem. 2001, 90, 157–173. [Google Scholar] [CrossRef]
- Khadka, N.K.; Cheng, X.; Ho, C.S.; Katsaras, J.; Pan, J. Interactions of the Anticancer Drug Tamoxifen with Lipid Membranes. Biophys. J. 2015, 108, 2492–2501. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, J.; Aranda, F.J.; Teruel, J.A.; Ortiz, A. Dissimilar action of tamoxifen and 4-hydroxytamoxifen on phosphatidylcholine model membranes. Biophys. Chem. 2021, 278, 106681. [Google Scholar] [CrossRef] [PubMed]
- Custódio, J.; Almeida, L.M.; Madeira, V.M. A reliable and rapid procedure to estimate drug partitioning in biomembranes. Biochem. Biophys. Res. Commun. 1991, 176, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Vance, J.E. Thematic Review Series: Glycerolipids. Phosphatidylserine and phosphatidylethanolamine in mammalian cells: Two metabolically related aminophospholipids. J. Lipid Res. 2008, 49, 1377–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, J.; Teruel, J.A.; Aranda, F.J.; Ortiz, A. Anticancer drugs tamoxifen and 4hydroxytamoxifen as effectors of phosphatidylethanolamine lipid polymorphism. Chem. Phys. Lipids 2022, 248, 105239. [Google Scholar] [CrossRef]
- Matsuzaki, K. Membrane Permeabilization Mechanisms. In Antimicrobial Peptides; Springer: Berlin/Heidelberg, Germany, 2019; Volume 1117, pp. 9–16. [Google Scholar] [CrossRef]
- Seddon, A.M.; Casey, D.; Law, R.V.; Gee, A.; Templer, R.H.; Ces, O. Drug interactions with lipid membranes. Chem. Soc. Rev. 2009, 38, 2509–2519. [Google Scholar] [CrossRef]
- Nicol, F.; Nir, S.; Szoka, F.C. Effect of Phospholipid Composition on an Amphipathic Peptide-Mediated Pore Formation in Bilayer Vesicles. Biophys. J. 2000, 78, 818–829. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Miao, Q.; Sun, Y.; Yang, F. tBid forms a pore in the liposome membrane. FEBS Lett. 2003, 555, 545–550. [Google Scholar] [CrossRef] [Green Version]
- Zaragoza, A.; Aranda, F.J.; Espuny, M.J.; Teruel, J.A.; Marqués, A.; Manresa, A.; Ortiz, A. Mechanism of Membrane Permeabilization by a Bacterial Trehalose Lipid Biosurfactant Produced by Rhodococcus sp. Langmuir 2009, 25, 7892–7898. [Google Scholar] [CrossRef]
- Sánchez, M.; Aranda, F.J.; Teruel, J.A.; Espuny, M.J.; Marqués, A.; Manresa, A.; Ortiz, A. Permeabilization of biological and artificial membranes by a bacterial dirhamnolipid produced by Pseudomonas aeruginosa. J. Colloid Interface Sci. 2010, 341, 240–247. [Google Scholar] [CrossRef]
- Fiedler, S.; Heerklotz, H. Vesicle Leakage Reflects the Target Selectivity of Antimicrobial Lipopeptides from Bacillus subtilis. Biophys. J. 2015, 109, 2079–2089. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.C.; Madeira, V.M.; Almeida, L.M.; Custódio, J.B. Hemolysis of human erythrocytes induced by tamoxifen is related to disruption of membrane structure. Biochim. Biophys. Acta (BBA) -Biomembr. 2000, 1464, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Khodov, I.A.; Musabirova, G.S.; Klochkov, V.V.; Karataeva, F.K.; Huster, D.; Scheidt, H.A. Structural details on the interaction of fenamates with lipid membranes. J. Mol. Liq. 2022, 367, 120502. [Google Scholar] [CrossRef]
- Pereira-Leite, C.; Figueiredo, M.; Burdach, K.; Nunes, C.; Reis, S. Unraveling the Role of Drug-Lipid Interactions in NSAIDs-Induced Cardiotoxicity. Membranes 2020, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Tomisato, W.; Tanaka, K.-I.; Katsu, T.; Kakuta, H.; Sasaki, K.; Tsutsumi, S.; Hoshino, T.; Aburaya, M.; Li, D.; Tsuchiya, T.; et al. Membrane permeabilization by non-steroidal anti-inflammatory drugs. Biochem. Biophys. Res. Commun. 2004, 323, 1032–1039. [Google Scholar] [CrossRef]
- Böttcher, C.; Van Gent, C.; Pries, C. A rapid and sensitive sub-micro phosphorus determination. Anal. Chim. Acta 1961, 24, 203–204. [Google Scholar] [CrossRef]
- Chen, R.F.; Knutson, J.R. Mechanism of fluorescence concentration quenching of carboxyfluorescein in liposomes: Energy transfer to nonfluorescent dimers. Anal. Biochem. 1988, 172, 61–77. [Google Scholar] [CrossRef]
- Nasr, G.; Greige-Gerges, H.; Elaissari, A.; Khreich, N. Liposomal membrane permeability assessment by fluorescence techniques: Main permeabilizing agents, applications and challenges. Int. J. Pharm. 2020, 580, 119198. [Google Scholar] [CrossRef]
- Ortiz, A.; Aranda, F.J.; Teruel, J.A. Interaction of Dirhamnolipid Biosurfactants with Phospholipid Membranes: A Molecular Level Study. In Biosurfactants; Springer: Berlin/Heidelberg, Germany, 2010; Volume 672, pp. 42–53. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem substance and compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Abraham, M.J.; van der Spoel, D.; Lindahl, E.; Hess, B.; Team, G.D. GROMACS User Manual, Version 5.0.7. Available online: www.gromacs.org (accessed on 26 January 2023).
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Brooks, B.R.; Brooks, C.L., III; MacKerell, A.D., Jr.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y.; et al. CHARMM-GUI Input Generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM Simulations Using the CHARMM36 Additive Force Field. J. Chem. Theory Comput. 2015, 12, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Mark, P.; Nilsson, L. Structure and dynamics of the TIP3P, SPC, and SPC/E water models at 298 K. J. Phys. Chem. A 2001, 105, 9954–9960. [Google Scholar] [CrossRef]
- Jo, S.; Lim, J.B.; Klauda, J.B.; Im, W. CHARMM-GUI Membrane Builder for Mixed Bilayers and Its Application to Yeast Membranes. Biophys. J. 2009, 97, 50–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjelkmar, P.; Larsson, P.; Cuendet, M.A.; Hess, B.; Lindahl, E. Implementation of the CHARMM Force Field in GROMACS: Analysis of Protein Stability Effects from Correction Maps, Virtual Interaction Sites, and Water Models. J. Chem. Theory Comput. 2010, 6, 459–466. [Google Scholar] [CrossRef]
- Martínez, L.; Andrade, R.; Birgin, E.G.; Martínez, J.M. PACKMOL: A package for building initial configurations for molecular dynamics simulations. J. Comput. Chem. 2009, 30, 2157–2164. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; Van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Hoover, W.G. Canonical dynamics: Equilibrium phase-space distributions. Phys. Rev. A 1985, 31, 1695–1697. [Google Scholar] [CrossRef] [Green Version]
- Parrinello, M.; Rahman, A. Polymorphic transitions in single crystals: A new molecular dynamics method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Schrödinger, L. The PyMOL Molecular Graphics System, Version 2.3. 2015. Available online: https://pymol.org/2/ (accessed on 26 January 2023).
- Van der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The critical role of phosphatidylcholine and phosphatidylethanolamine metabolism in health and disease. Biochim. Biophys. Acta (BBA) -Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef]
- Tada, K.; Goto, M.; Tamai, N.; Matsuki, H.; Kaneshina, S. Pressure effect on the bilayer phase transition of asymmetric lipids with an unsaturated acyl chain. Ann. N. Y. Acad. Sci. 2010, 1189, 77–85. [Google Scholar] [CrossRef]
- Trombetta, D.; Castelli, F.; Sarpietro, M.G.; Venuti, V.; Cristani, M.; Daniele, C.; Saija, A.; Mazzanti, G.; Bisignano, G. Mechanisms of Antibacterial Action of Three Monoterpenes. Antimicrob. Agents Chemother. 2005, 49, 2474–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, H.I.; Lee, S.K.; Kim, J.; Kim, S.W.; Yu, J.; Bae, S.Y.; Lee, J.E.; Nam, S.J.; Lee, S.-Y. Variations in plasma concentrations of tamoxifen metabolites and the effects of genetic polymorphisms on tamoxifen metabolism in Korean patients with breast cancer. Oncotarget 2017, 8, 100296–100311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, M.; Mukhopadhyay, C. Molecular Dynamics Simulations of the Interactions of Kinin Peptides with an Anionic POPG Bilayer. Langmuir 2011, 27, 3713–3722. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.N.; McElhaney, R.N. The structure and organization of phospholipid bilayers as revealed by infrared spectroscopy. Chem. Phys. Lipids 1998, 96, 9–21. [Google Scholar] [CrossRef]
- Lewis, R.N.; McElhaney, R.N.; Pohle, W.; Mantsch, H.H. Components of the carbonyl stretching band in the infrared spectra of hydrated 1,2-diacylglycerolipid bilayers: A reevaluation. Biophys. J. 1994, 67, 2367–2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, R.; McElhaney, R. Calorimetric and spectroscopic studies of the polymorphic phase behavior of a homologous series of n-saturated 1,2-diacyl phosphatidylethanolamines. Biophys. J. 1993, 64, 1081–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, A.; Aranda, F.J.; Gomez-Fernandez, J.C. A differential scanning calorimetry study of the interaction of a-tocopherol. Biochim. Biophys. Acta (BBA) -Biomembr. 1987, 898, 214–222. [Google Scholar] [CrossRef]
- Friedman, R.; Khalid, S.; Aponte-Santamaría, C.; Arutyunova, E.; Becker, M.; Boyd, K.J.; Christensen, M.; Coimbra, J.T.S.; Concilio, S.; Daday, C.; et al. Understanding Conformational Dynamics of Complex Lipid Mixtures Relevant to Biology. J. Membr. Biol. 2018, 251, 609–631. [Google Scholar] [CrossRef] [Green Version]
- Clifford, R.; Bowden, D.; Blower, E.; Kirwan, C.; Vimalachandran, D. Does tamoxifen have a therapeutic role outside of breast cancer? A systematic review of the evidence. Surg. Oncol. 2020, 33, 100–107. [Google Scholar] [CrossRef]
- Wiseman, H.; Quinn, P.; Halliwell, B. Tamoxifen and related compounds decrease membrane fluidity in liposomes. FEBS Lett. 1993, 330, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Suwalsky, M.; Hernández, P.; Villena, F.; Aguilar, F.; Sotomayor, C.P. Interaction of the Anticancer Drug Tamoxifen with the Human Erythrocyte Membrane and Molecular Models. Z. Nat. C 1998, 53, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Furse, S. Is phosphatidylglycerol essential for terrestrial life? J. Chem. Biol. 2016, 10, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veiko, A.G.; Sekowski, S.; Lapshina, E.A.; Wilczewska, A.Z.; Markiewicz, K.H.; Zamaraeva, M.; Zhao, H.-C.; Zavodnik, I.B. Flavonoids modulate liposomal membrane structure, regulate mitochondrial membrane permeability and prevent erythrocyte oxidative damage. Biochim. Biophys. Acta (BBA) -Biomembr. 2020, 1862, 183442. [Google Scholar] [CrossRef] [PubMed]
- Kicinska, A.; Jarmuszkiewicz, W. Flavonoids and Mitochondria: Activation of Cytoprotective Pathways? Molecules 2020, 25, 3060. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, V.; Uaratanawong, R.; Patel, R.R.; Patel, H.; Bao, W.; Hartney, B.; Cohen, E.; Chen, X.; Zhong, Q.; Isales, C.M.; et al. Phosphatidylglycerol Inhibits Toll-Like Receptor–Mediated Inflammation by Danger-Associated Molecular Patterns. J. Investig. Dermatol. 2019, 139, 868–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuronuma, K.; Mitsuzawa, H.; Takeda, K.; Nishitani, C.; Chan, E.D.; Kuroki, Y.; Nakamura, M.; Voelker, D.R. Anionic Pulmonary Surfactant Phospholipids Inhibit Inflammatory Responses from Alveolar Macrophages and U937 Cells by Binding the Lipopolysaccharide-interacting Proteins CD14 and MD-2. J. Biol. Chem. 2009, 284, 25488–25500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.-W.; Chao, Y.-J.; Chang, W.-H.; Chan, J.-F.; Hsu, Y.-H.H. Phosphatidylglycerol Incorporates into Cardiolipin to Improve Mitochondrial Activity and Inhibits Inflammation. Sci. Rep. 2018, 8, 4919. [Google Scholar] [CrossRef]
- Numata, M.; Kandasamy, P.; Nagashima, Y.; Posey, J.; Hartshorn, K.; Woodland, D.; Voelker, D.R. Phosphatidylglycerol Suppresses Influenza A Virus Infection. Am. J. Respir. Cell Mol. Biol. 2012, 46, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.T.-H.; Chan, K.-G.; Pusparajah, P.; Lee, W.-L.; Chuah, L.-H.; Khan, T.M.; Lee, L.-H.; Goh, B.-H. Targeting Membrane Lipid a Potential Cancer Cure? Front. Pharmacol. 2017, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Koynova, R.; Caffrey, M. Phases and phase transitions of the hydrated phosphatidylethanolamines. Chem. Phys. Lipids 1994, 69, 1–34. [Google Scholar] [CrossRef]
- Boggs, J.M. Lipid intermolecular hydrogen bonding: Influence on structural organization and membrane function. Biochim. Biophys. Acta (BBA) -Rev. Biomembr. 1987, 906, 353–404. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| [TMX] (μM) | k1 (s−1) ± SD | % ± SD | k2 (s−1) ± SD | % ± SD |
|---|---|---|---|---|
| 2.5 | 1.4 × 10−4 ± 0.62 × 10−4 | 98.0 ± 0 | 0.035 ± 0.0070 | 2.0 ± 0 |
| 5 | 1.4 × 10−4 ± 0.51 × 10−4 | 97.7 ± 0.35 | 0.035 ± 0.0084 | 2.3 ± 0.35 |
| 7.5 | 5.5 × 10−4 ± 0.71 × 10−4 | 96.8 ± 0.70 | 0.036 ± 0.0077 | 3.2 ± 0.70 |
| 17.5 | 25 × 10−4 ± 1.1 × 10−4 | 86.8 ± 0.85 | 0.028 ± 0.0028 | 13.2 ± 0.84 |
| 25 | 31 × 10−4 ± 5.6 × 10−4 | 59.9 ± 8.70 | 0.023 ± 0.0028 | 40.1 ± 8.76 |
| 50 | 115 × 10−4 ± 20.5 × 10−4 | 45.6 ± 5.10 | 0.047 ± 0.0160 | 54.4 ± 5.09 |
| [HTMX] (μM) | k1 (s−1) ± SD | % ± SD | k2 (s−1) ± SD | % ± SD |
| 2.5 | 0.89 × 10−4 ± 0.50 × 10−4 | 99.8 ± 0.22 | 0.042 ± 0.0082 | 0.2 ± 0.025 |
| 5 | 0.72 × 10−4 ± 0.68 × 10−4 | 98.5 ± 0.75 | 0.028 ± 0.0035 | 1.5 ± 0.120 |
| 7.5 | 2.4 × 10−4 ± 0.90 × 10−4 | 99.5 ± 0.84 | 0.049 ± 0.0078 | 0.5 ± 0.040 |
| 17.5 | 2.2 × 10−4 ± 0.44 × 10−4 | 98.6 ± 0.90 | 0.035 ± 0.0028 | 1.4 ± 0.008 |
| 25 | 2.7 × 10−4 ± 0.72 × 10−4 | 98.7 ± 0.88 | 0.045 ± 0.0014 | 1.3 ± 0.006 |
| 50 | 4.8 × 10−4 ± 0.62 × 10−4 | 98.9 ± 0.94 | 0.050 ± 0.0063 | 1.1 ± 0.004 |
| Tamoxifen | ||||
| Composition | k1 (s−1) ± SD | % ± SD | k2 (s−1) ± SD | % ± SD |
| POPC | 25 × 10−4 ± 1.1 × 10−4 | 86.8 ± 0.85 | 0.028 ± 0.0028 | 13.2 ± 0.84 |
| POPC/POPG | 18 × 10−4 ± 1 × 10−4 | 85.7 ± 0.98 | 0.036 ± 0.0052 | 14.3 ± 0.98 |
| POPC/POPE | 9.0 × 10−4 ± 1.4 × 10−4 | 100 ± 0 | ||
| 4−Hydroxytamoxifen | ||||
| Composition | k1 (s−1) ± SD | % ± SD | k2 (s−1) ± SD | % ± SD |
| POPC | 2.2 × 10−4 ± 0.44 × 10−4 | 98.6 ± 0.90 | 0.035 ± 0.0028 | 1.4 ± 0.008 |
| POPC/POPG | 1.9 × 10−4 ± 0.32 × 10−4 | 100 ± 0 | ||
| POPC/POPE | 2.0 × 10−4 ± 0.50 × 10−4 | 100 ± 0 | ||
| Composition | νCH2 (cm−1) | νCO (cm−1) |
|---|---|---|
| POPC | 2847.1 | 1723.2 |
| POPC/TMX (5:1.7) | 2852.3 | 1730.7 |
| POPC/POPE (5:1.7) | 2847.8 | 1723.0 |
| POPC/POPE/TMX (5:1.7:2.3) | 2852.4 | 1730.7 |
| POPC/POPG (5:1.7) | 2852.0 | 1729.2 |
| POPC/POPG/TMX (5:1.7:2.3) | 2853.0 | 1732.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortiz, J.; Teruel, J.A.; Aranda, F.J.; Ortiz, A. On the Mechanism of Membrane Permeabilization by Tamoxifen and 4-Hydroxytamoxifen. Membranes 2023, 13, 292. https://doi.org/10.3390/membranes13030292
Ortiz J, Teruel JA, Aranda FJ, Ortiz A. On the Mechanism of Membrane Permeabilization by Tamoxifen and 4-Hydroxytamoxifen. Membranes. 2023; 13(3):292. https://doi.org/10.3390/membranes13030292
Chicago/Turabian StyleOrtiz, Julia, José A. Teruel, Francisco J. Aranda, and Antonio Ortiz. 2023. "On the Mechanism of Membrane Permeabilization by Tamoxifen and 4-Hydroxytamoxifen" Membranes 13, no. 3: 292. https://doi.org/10.3390/membranes13030292