

Pharmacological Characterization of a Recombinant Mitochondrial ROMK2 Potassium Channel Expressed in Bacteria and Reconstituted in Planar Lipid Bilayers



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
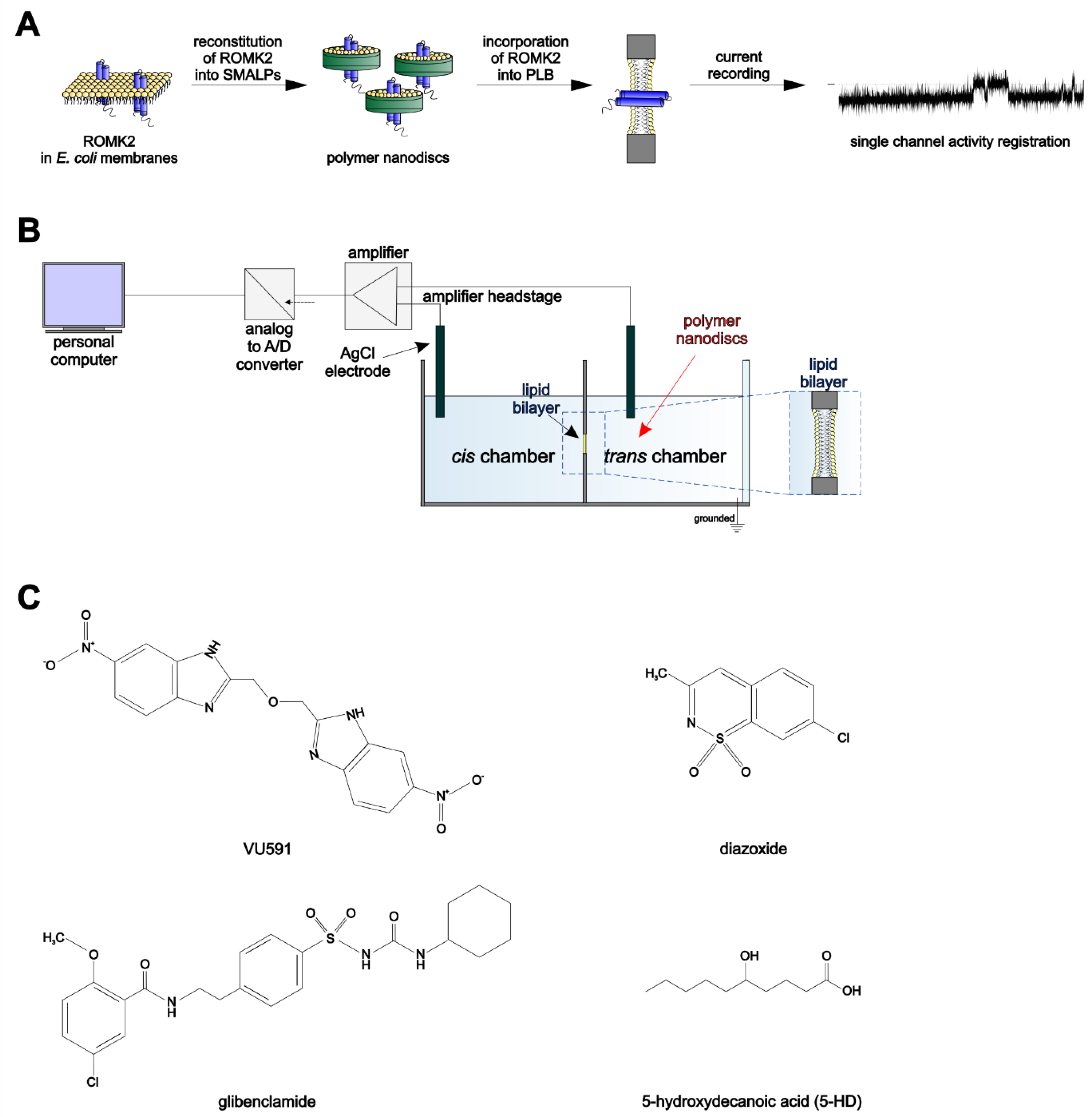
2.2. Preparation of Polymer Nanodiscs Containing ROMK2 Protein
2.3. Planar Lipid Bilayer (PLB) Measurements
2.4. Data Analysis
3. Results
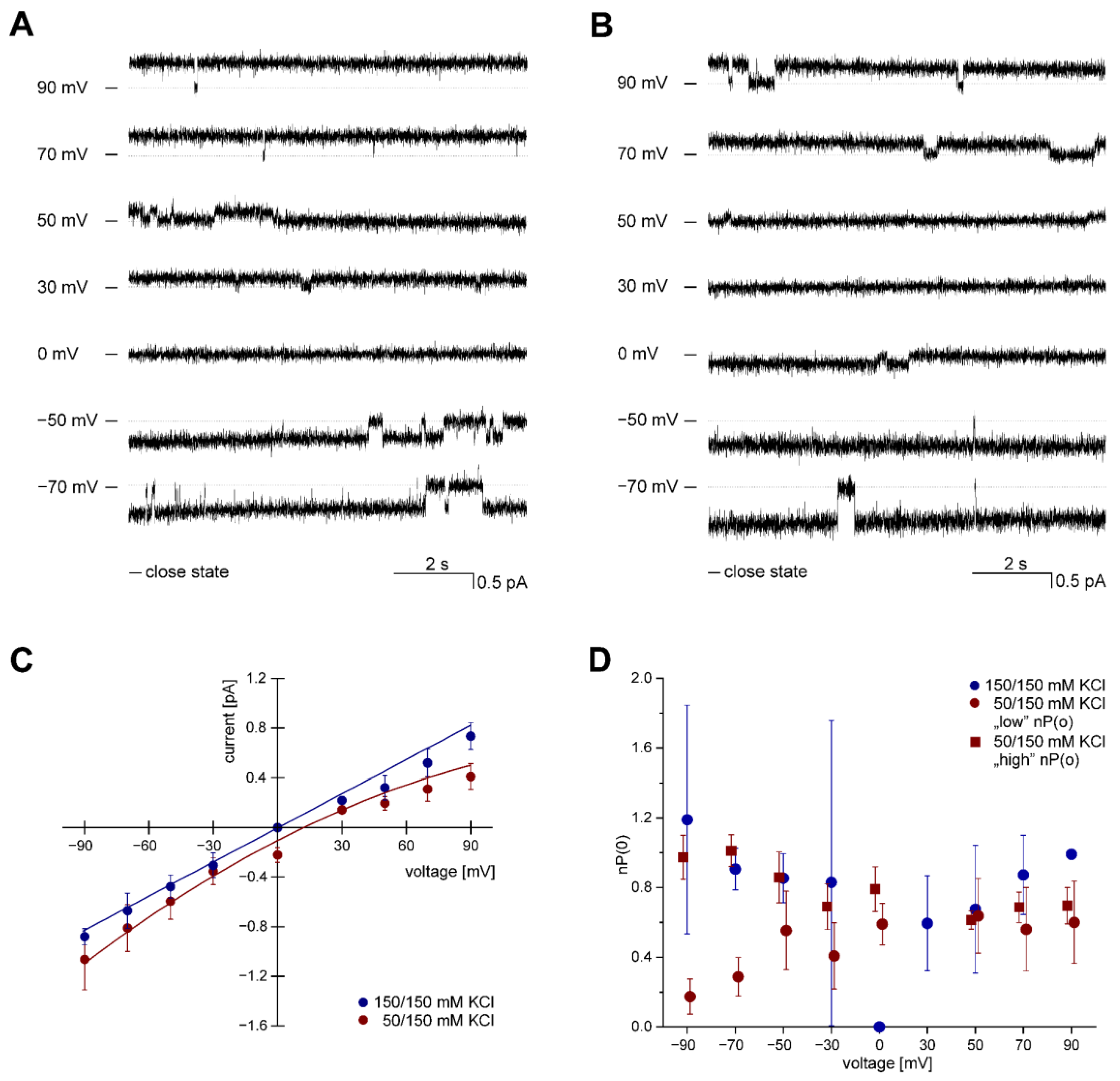
3.1. Identification of ROMK2 Single-Channel Activity
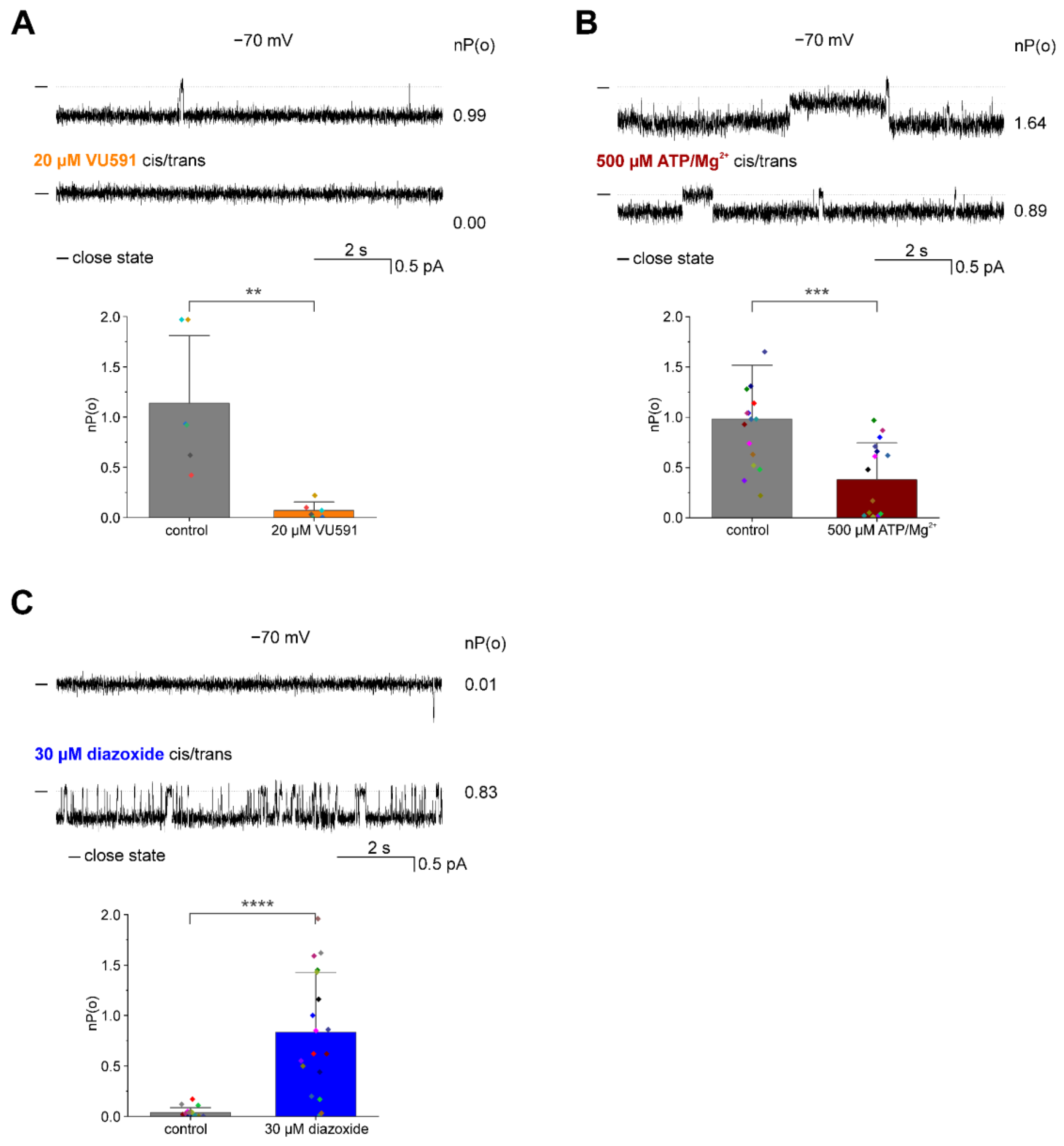
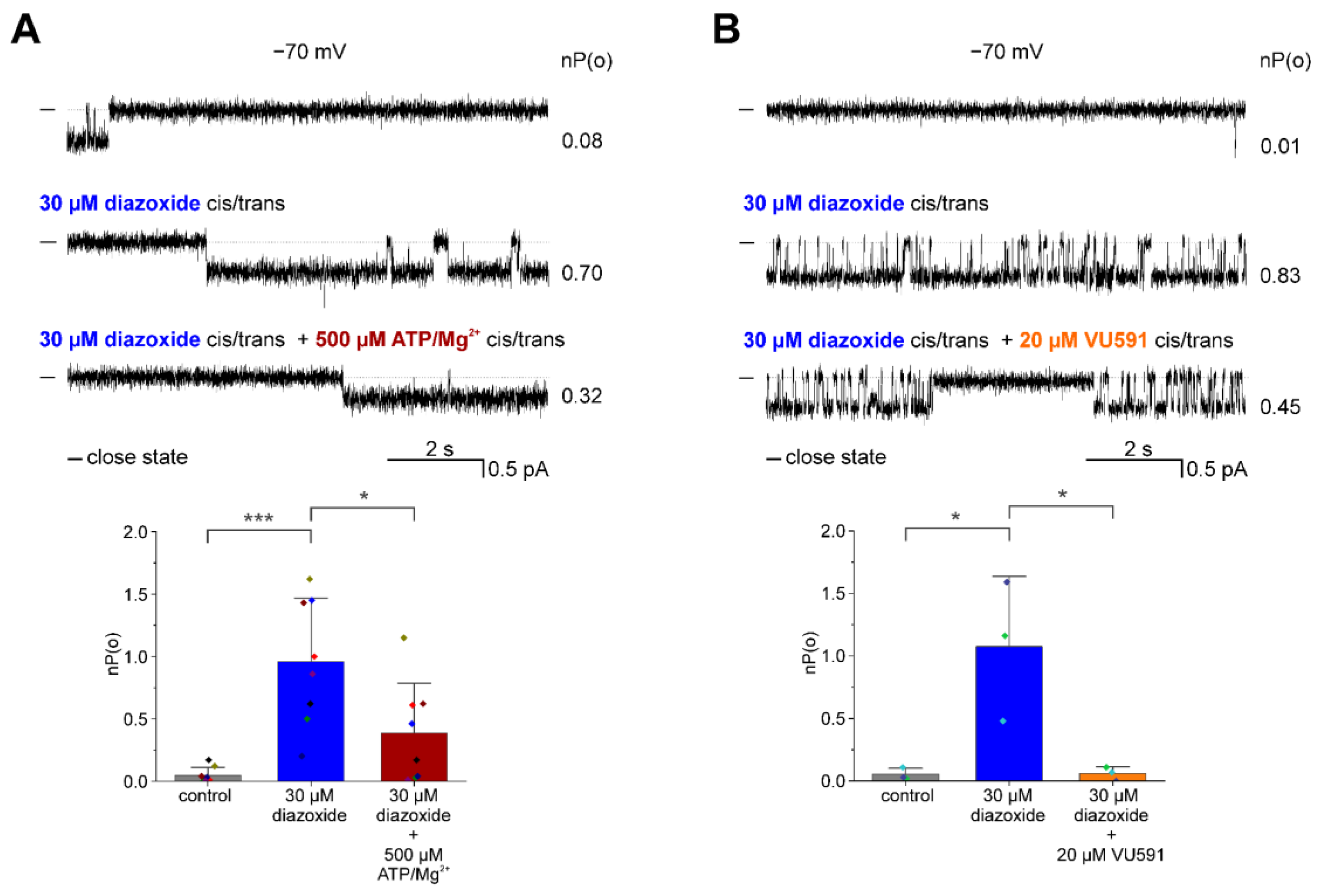
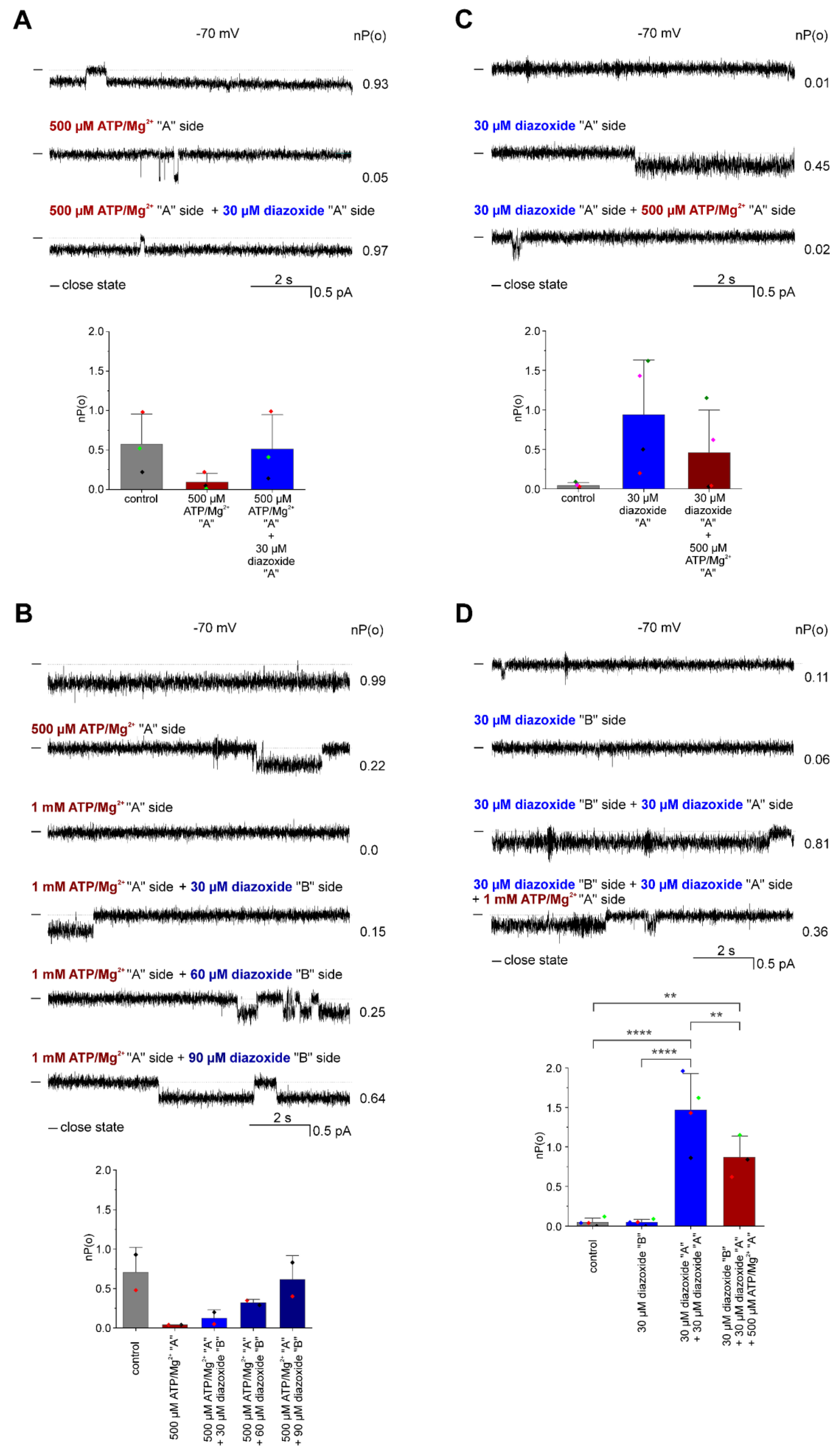
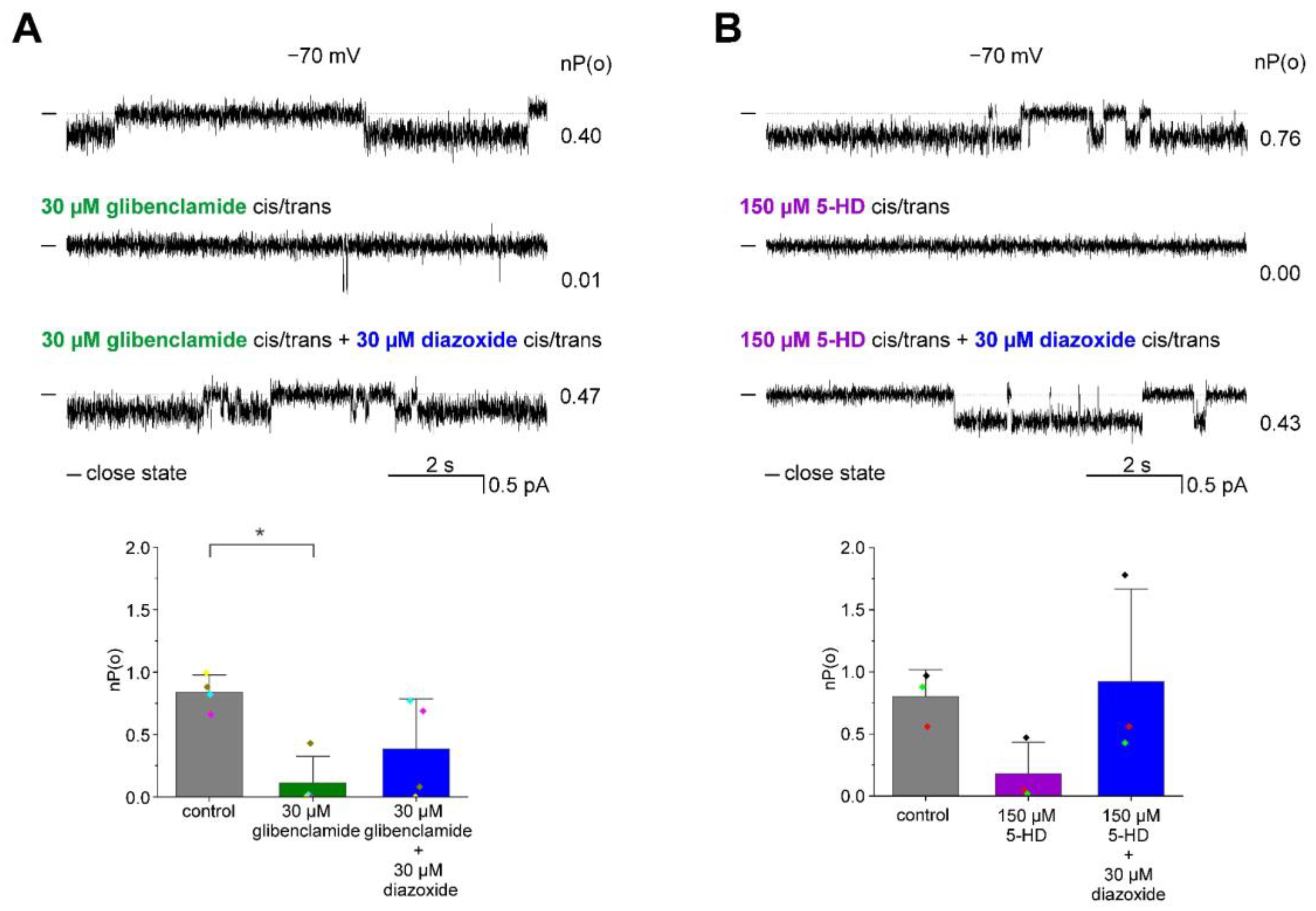
3.2. Pharmacological Modulation of the ROMK2 Channel Activity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inoue, I.; Nagase, H.; Kishi, K.; Higuti, T. ATP-sensitive K+ channel in the mitochondrial inner membrane. Nature 1991, 352, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Paucek, P.; Mironova, G.; Mahdi, F.; Beavis, A.; Woldegiorgis, G.; Garlid, K. Reconstitution and partial purification of the glibenclamide-sensitive, ATP-dependent K+ channel from rat liver and beef heart mitochondria. J. Biol. Chem. 1992, 267, 26062–26069. [Google Scholar] [CrossRef] [PubMed]
- Garlid, K.D.; Paucek, P.; Yarov-Yarovoy, V.; Sun, X.; Schindler, P.A. The Mitochondrial KATP Channel as a Receptor for Potassium Channel Openers. J. Biol. Chem. 1996, 271, 8796–8799. [Google Scholar] [CrossRef] [Green Version]
- Dȩbska, G.; May, R.; Kicinska, A.; Szewczyk, A.; Elger, C.E.; Kunz, W. Potassium channel openers depolarize hippocampal mitochondria. Brain Res. 2001, 892, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Bajgar, R.; Seetharaman, S.; Kowaltowski, A.J.; Garlid, K.D.; Paucek, P. Identification and Properties of a Novel Intracellular (Mitochondrial) ATP-sensitive Potassium Channel in Brain. J. Biol. Chem. 2001, 276, 33369–33374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choma, K.; Bednarczyk, P.; Koszela-Piotrowska, I.; Kulawiak, B.; Kudin, A.; Kunz, W.S.; Dołowy, K.; Szewczyk, A. Single channel studies of the ATP-regulated potassium channel in brain mitochondria. J. Bioenerg. Biomembr. 2009, 41, 323–334. [Google Scholar] [CrossRef]
- Cancherini, D.V.; Trabuco, L.G.; Rebouças, N.A.; Kowaltowski, A.J. ATP-sensitive K+ channels in renal mitochondria. Am. J. Physiol. Renal Physiol. 2003, 285, F1291–F1296. [Google Scholar] [CrossRef]
- Debska, G.; Kicinska, A.; Skalska, J.; Szewczyk, A.; May, R.; Elger, C.E.; Kunz, W.S. Opening of potassium channels modulates mitochondrial function in rat skeletal muscle. Biochim. Biophys. Acta 2002, 1556, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Dahlem, Y.A.; Horn, T.F.; Buntinas, L.; Gonoi, T.; Wolf, G.; Siemen, D. The human mitochondrial KATP channel is modulated by calcium and nitric oxide: A patch-clamp approach. Biochim. Biophys. Acta 2004, 1656, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Bednarczyk, P.; Kicinska, A.; Laskowski, M.; Kulawiak, B.; Kampa, R.; Walewska, A.; Krajewska, M.; Jarmuszkiewicz, W.; Szewczyk, A. Evidence for a mitochondrial ATP-regulated potassium channel in human dermal fibroblasts. Biochim. Biophys. Acta 2018, 1859, 309–318. [Google Scholar] [CrossRef]
- Laskowski, M.; Augustynek, B.; Bednarczyk, P.; Żochowska, M.; Kalisz, J.; O’Rourke, B.; Szewczyk, A.; Kulawiak, B. Single-Channel Properties of the ROMK-Pore-Forming Subunit of the Mitochondrial ATP-Sensitive Potassium Channel. Int. J. Mol. Sci. 2019, 20, 5323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.X.; Chen, Y.-F.; Campbell, W.B.; Zou, A.-P.; Gross, G.J.; Li, P.-L. Characteristics and Superoxide-Induced Activation of Reconstituted Myocardial Mitochondrial ATP-Sensitive Potassium Channels. Circ. Res. 2001, 89, 1177–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakae, Y.; Kwok, W.-M.; Bosnjak, Z.J.; Jiang, M.T. Isoflurane activates rat mitochondrial ATP-sensitive K+ channels reconstituted in lipid bilayers. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H1865–H1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paggio, A.; Checchetto, V.; Campo, A.; Menabò, R.; Di Marco, G.; Di Lisa, F.; Szabo, I.; Rizzuto, R.; De Stefani, D. Identification of an ATP-sensitive potassium channel in mitochondria. Nature 2019, 572, 609–613. [Google Scholar] [CrossRef]
- Miki, T.; Suzuki, M.; Shibasaki, T.; Uemura, H.; Sato, T.; Yamaguchi, K.; Koseki, H.; Iwanaga, T.; Nakaya, H.; Seino, S. Mouse model of Prinzmetal angina by disruption of the inward rectifier Kir6.1. Nat. Med. 2002, 8, 466–472. [Google Scholar] [CrossRef]
- Wojtovich, A.P.; Urciuoli, W.R.; Chatterjee, S.; Fisher, A.B.; Nehrke, K.; Brookes, P.S. Kir6.2 is not the mitochondrial KATP channel but is required for cardioprotection by ischemic preconditioning. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H1439–H1445. [Google Scholar] [CrossRef] [Green Version]
- Ardehali, H.; Chen, Z.; Ko, Y.; Mejía-Alvarez, R.; Marbán, E. Multiprotein complex containing succinate dehydrogenase confers mitochondrial ATP-sensitive K + channel activity. Proc. Natl. Acad. Sci. USA 2004, 101, 11880–11885. [Google Scholar] [CrossRef] [Green Version]
- Foster, D.B.; Ho, A.S.; Rucker, J.; Garlid, A.O.; Chen, L.; Sidor, A.; Garlid, K.D.; O’Rourke, B. Mitochondrial ROMK Channel Is a Molecular Component of MitoK(ATP). Circ. Res. 2012, 111, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Juhaszova, M.; Kobrinsky, E.; Zorov, D.B.; Nuss, H.B.; Yaniv, Y.; Fishbein, K.W.; de Cabo, R.; Montoliu, L.; Gabelli, S.B.; Aon, M.A.; et al. ATP Synthase K(+)- and H(+)-Fluxes Drive ATP Synthesis and Enable Mitochondrial K(+)-“Uniporter” Function: I. Characterization of Ion Fluxes. Function 2021, 3, zqab065. [Google Scholar] [CrossRef]
- Juhaszova, M.; Kobrinsky, E.; Zorov, D.B.; Nuss, H.B.; Yaniv, Y.; Fishbein, K.W.; de Cabo, R.; Montoliu, L.; Gabelli, S.B.; Aon, M.A.; et al. ATP Synthase K(+)- and H(+)-fluxes Drive ATP Synthesis and Enable Mitochondrial K(+)-“Uniporter” Function: II. Ion and ATP Synthase Flux Regulation. Function 2022, 3, zqac001. [Google Scholar] [CrossRef]
- Krajewska, M.; Koprowski, P. Solubilization, purification, and functional reconstitution of human ROMK potassium channel in copolymer styrene-maleic acid (SMA) nanodiscs. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183555. [Google Scholar] [CrossRef]
- Stefanowska, A.; Koprowski, P.; Bednarczyk, P.; Szewczyk, A.; Krysinski, P. Electrochemical studies of the mitochondrial ROMK2 potassium channel activity reconstituted into the free-standing and tethered bilayer lipid membranes. Bioelectrochemistry 2023, 151, 108372. [Google Scholar] [CrossRef] [PubMed]
- Kulawiak, B.; Bednarczyk, P. Reconstitution of brain mitochondria inner membrane into planar lipid bilayer. Acta Neurobiol. Exp. 2005, 65, 271–276. [Google Scholar]
- Skalska, J.; Bednarczyk, P.; Piwońska, M.; Kulawiak, B.; Wilczynski, G.; Dołowy, K.; Kunz, W.S.; Kudin, A.P.; Szewczyk, A. Calcium Ions Regulate K+ Uptake into Brain Mitochondria: The Evidence for a Novel Potassium Channel. Int. J. Mol. Sci. 2009, 10, 1104–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhave, G.; Chauder, B.A.; Liu, W.; Dawson, E.S.; Kadakia, R.; Nguyen, T.T.; Lewis, L.M.; Meiler, J.; Weaver, C.D.; Satlin, L.M.; et al. Development of a Selective Small-Molecule Inhibitor of Kir1.1, the Renal Outer Medullary Potassium Channel. Mol. Pharmacol. 2010, 79, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swale, D.R.; Sheehan, J.H.; Banerjee, S.; Husni, A.S.; Nguyen, T.T.; Meiler, J.; Denton, J.S. Computational and Functional Analyses of a Small-Molecule Binding Site in ROMK. Biophys. J. 2015, 108, 1094–1103. [Google Scholar] [CrossRef] [Green Version]
- Kicinska, A.; Swida, A.; Bednarczyk, P.; Koszela-Piotrowska, I.; Choma, K.; Dolowy, K.; Szewczyk, A.; Jarmuszkiewicz, W. ATP-sensitive Potassium Channel in Mitochondria of the Eukaryotic Microorganism Acanthamoeba castellanii. J. Biol. Chem. 2007, 282, 17433–17441. [Google Scholar] [CrossRef] [Green Version]
- Garlid, K.D.; Paucek, P.; Yarov-Yarovoy, V.; Murray, H.N.; Darbenzio, R.B.; D’Alonzo, A.J.; Lodge, N.J.; Smith, M.A.; Grover, G.J. Cardioprotective Effect of Diazoxide and Its Interaction With Mitochondrial ATP-Sensitive K+ Channels. Circ. Res. 1997, 81, 1072–1082. [Google Scholar] [CrossRef]
- McNicholas, C.M.; Yang, Y.; Giebisch, G.; Hebert, S.C. Molecular site for nucleotide binding on an ATP-sensitive renal K+ channel (ROMK2). Am. J. Physiol. 1996, 271, F275–F285. [Google Scholar] [CrossRef]
- MacGregor, G.G.; Dong, K.; Vanoye, C.G.; Tang, L.; Giebisch, G.; Hebert, S.C. Nucleotides and phospholipids compete for binding to the C terminus of KATP channels. Proc. Natl. Acad. Sci. USA 2002, 99, 2726–2731. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Leng, Q.; Egan, M.E.; Caplan, M.J.; Boulpaep, E.L.; Giebisch, G.H.; Hebert, S.C. CFTR is required for PKA-regulated ATP sensitivity of Kir1.1 potassium channels in mouse kidney. J. Clin. Investig. 2006, 116, 797–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konstas, A.-A.; Dabrowski, M.; Korbmacher, C.; Tucker, S.J. Intrinsic sensitivity of Kir1.1 (ROMK) to glibenclamide in the absence of SUR2B. Implications for the identity of the renal ATP-regulated secretory K+ channel. J. Biol. Chem. 2002, 277, 21346–21351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Sato, T.; Seharaseyon, J.; Szewczyk, A.; O’Rourke, B.; Marban, E. Mitochondrial ATP-Dependent Potassium Channels: Viable Candidate Effectors of Ischemic Preconditioninga. Ann. N. Y. Acad. Sci. 1999, 874, 27–37. [Google Scholar] [CrossRef]
- Light, P.E.; Kanji, H.D.; Fox, J.E.M.; French, R.J. Distinct myoprotective roles of cardiac sarcolemmal and mitochondrial K ATP channels during metabolic inhibition and recovery. FASEB J. 2001, 15, 2586–2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustynek, B.; Kunz, W.S.; Szewczyk, A. Guide to the Pharmacology of Mitochondrial Potassium Channels. Handb. Exp. Pharmacol. 2016, 240, 103–127. [Google Scholar]
- Welling, P.A.; Ho, K. A comprehensive guide to the ROMK potassium channel: Form and function in health and disease. Am. J. Physiol. Renal Physiol. 2009, 297, F849–F863. [Google Scholar] [CrossRef] [Green Version]
- Nichols, C.G.; Ho, K.; Hébert, S. Mg(2+)-dependent inward rectification of ROMK1 potassium channels expressed in Xenopus oocytes. J. Physiol. 1994, 476, 399–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, L.G.; Choe, H.; Frindt, G. Is the secretory K channel in the rat CCT ROMK? Am. J. Physiol. 1997, 273, F404–F410. [Google Scholar] [CrossRef]
- Ho, K.; Nichols, C.G.; Lederer, W.J.; Lytton, J.; Vassilev, P.M.; Kanazirska, M.V.; Hebert, S.C. Cloning and expression of an inwardly rectifying ATP-regulated potassium channel. Nature 1993, 363, 31–38. [Google Scholar] [CrossRef]
- Liou, H.-H.; Zhou, S.-S.; Huang, C.-L. Regulation of ROMK1 channel by protein kinase A via a phosphatidylinositol 4,5-bisphosphate-dependent mechanism. Proc. Natl. Acad. Sci. USA 1999, 96, 5820–5825. [Google Scholar] [CrossRef] [Green Version]
- Heginbotham, L.; Kolmakova-Partensky, L.; Miller, C. Functional Reconstitution of a Prokaryotic K+ Channel. J. Gen. Physiol. 1998, 111, 741–749. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.W.; D’Avanzo, N.; Doyle, D.A.; Nichols, C.G. Dual-Mode Phospholipid Regulation of Human Inward Rectifying Potassium Channels. Biophys. J. 2011, 100, 620–628. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-J.; Wang, S.; Borschel, W.; Heyman, S.; Gyore, J.; Nichols, C.G. Secondary anionic phospholipid binding site and gating mechanism in Kir2.1 inward rectifier channels. Nat. Commun. 2013, 4, 2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, R.; He, S.; Black, K.A.; Clarke, O.B.; Wu, D.; Bolla, J.R.; Johnson, P.; Periasamy, A.; Wardak, A.; Czabotar, P.; et al. Ion currents through Kir potassium channels are gated by anionic lipids. Nat. Commun. 2022, 13, 490. [Google Scholar] [CrossRef] [PubMed]
- McNicholas, C.M.; MacGregor, G.G.; Islas, L.D.; Yang, Y.; Hebert, S.C.; Giebisch, G. pH-dependent modulation of the cloned renal K+ channel, ROMK. Am. J. Physiol. 1998, 275, F972–F981. [Google Scholar] [CrossRef]
- Ruknudin, A.; Schulze, D.H.; Sullivan, S.K.; Lederer, W.J.; Welling, P.A. Novel Subunit Composition of a Renal Epithelial KATP Channel. J. Biol. Chem. 1998, 273, 14165–14171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, K.; Tang, L.; MacGregor, G.G.; Hebert, S.C. Localization of the ATP/Phosphatidylinositol 4,5 Diphosphate-binding Site to a 39-Amino Acid Region of the Carboxyl Terminus of the ATP-regulated K+ Channel Kir1.1. J. Biol. Chem. 2002, 277, 49366–49373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luzi, L.; Pozza, G. Glibenclamide: An old drug with a novel mechanism of action? Acta Diabetol. 1997, 34, 239–244. [Google Scholar] [CrossRef]
- Wang, M.; Wu, J.-X.; Ding, D.; Chen, L. Structural insights into the mechanism of pancreatic KATP channel regulation by nucleotides. Nat. Commun. 2022, 13, 2770. [Google Scholar] [CrossRef]
- Palácio, P.B.; Lucas, A.M.B.; Alexandre, J.V.D.L.; Cunha, P.L.O.; Viana, Y.I.P.; Albuquerque, A.C.; Varela, A.L.N.; Facundo, H.T. Pharmacological and molecular docking studies reveal that glibenclamide competitively inhibits diazoxide-induced mitochondrial ATP-sensitive potassium channel activation and pharmacological preconditioning. Eur. J. Pharmacol. 2021, 908, 174379. [Google Scholar] [CrossRef]
- Coetzee, W.A. Multiplicity of effectors of the cardioprotective agent, diazoxide. Pharmacol. Ther. 2013, 140, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contessi, S.; Metelli, G.; Mavelli, I.; Lippe, G. Diazoxide affects the IF1 inhibitor protein binding to F1 sector of beef heart F0F1ATPsynthase. Biochem. Pharmacol. 2004, 67, 1843–1851. [Google Scholar] [CrossRef] [PubMed]
- Hambrock, A.; Löffler-Walz, C.; Quast, U. Glibenclamide binding to sulphonylurea receptor subtypes: Dependence on adenine nucleotides. Br. J. Pharmacol. 2002, 136, 995–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, I.; Zoratti, M. Mitochondrial Channels: Ion Fluxes and More. Physiol. Rev. 2014, 94, 519–608. [Google Scholar] [CrossRef]
- Jabůrek, M.; Yarov-Yarovoy, V.; Paucek, P.; Garlid, K.D. State-dependent Inhibition of the Mitochondrial KATP Channel by Glyburide and 5-Hydroxydecanoate. J. Biol. Chem. 1998, 273, 13578–13582. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Li, Y.; Saito, T.; Nakaya, H. Minoxidil opens mitochondrial KATP channels and confers cardioprotection. Br. J. Pharmacol. 2004, 141, 360–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bednarczyk, P.; Kicińska, A.; Kominkova, V.; Ondrias, K.; Dolowy, K.; Szewczyk, A. Quinine Inhibits Mitochondrial ATP-regulated Potassium Channel from Bovine Heart. J. Membr. Biol. 2004, 199, 63–72. [Google Scholar] [CrossRef]
- Bednarczyk, P.; Dołowy, K.; Szewczyk, A. New properties of mitochondrial ATP-regulated potassium channels. J. Bioenerg. Biomembr. 2008, 40, 325–335. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krajewska, M.; Szewczyk, A.; Kulawiak, B.; Koprowski, P. Pharmacological Characterization of a Recombinant Mitochondrial ROMK2 Potassium Channel Expressed in Bacteria and Reconstituted in Planar Lipid Bilayers. Membranes 2023, 13, 360. https://doi.org/10.3390/membranes13030360
Krajewska M, Szewczyk A, Kulawiak B, Koprowski P. Pharmacological Characterization of a Recombinant Mitochondrial ROMK2 Potassium Channel Expressed in Bacteria and Reconstituted in Planar Lipid Bilayers. Membranes. 2023; 13(3):360. https://doi.org/10.3390/membranes13030360
Chicago/Turabian StyleKrajewska, Milena, Adam Szewczyk, Bogusz Kulawiak, and Piotr Koprowski. 2023. "Pharmacological Characterization of a Recombinant Mitochondrial ROMK2 Potassium Channel Expressed in Bacteria and Reconstituted in Planar Lipid Bilayers" Membranes 13, no. 3: 360. https://doi.org/10.3390/membranes13030360
APA StyleKrajewska, M., Szewczyk, A., Kulawiak, B., & Koprowski, P. (2023). Pharmacological Characterization of a Recombinant Mitochondrial ROMK2 Potassium Channel Expressed in Bacteria and Reconstituted in Planar Lipid Bilayers. Membranes, 13(3), 360. https://doi.org/10.3390/membranes13030360