
First Suction Cup Tagging on a Small and Coastal Form Bryde’s Whale (Balaenoptera edeni edeni) in China to Investigate Its Dive Profiles and Foraging Behaviours



Abstract
1. Introduction
2. Materials and Methods
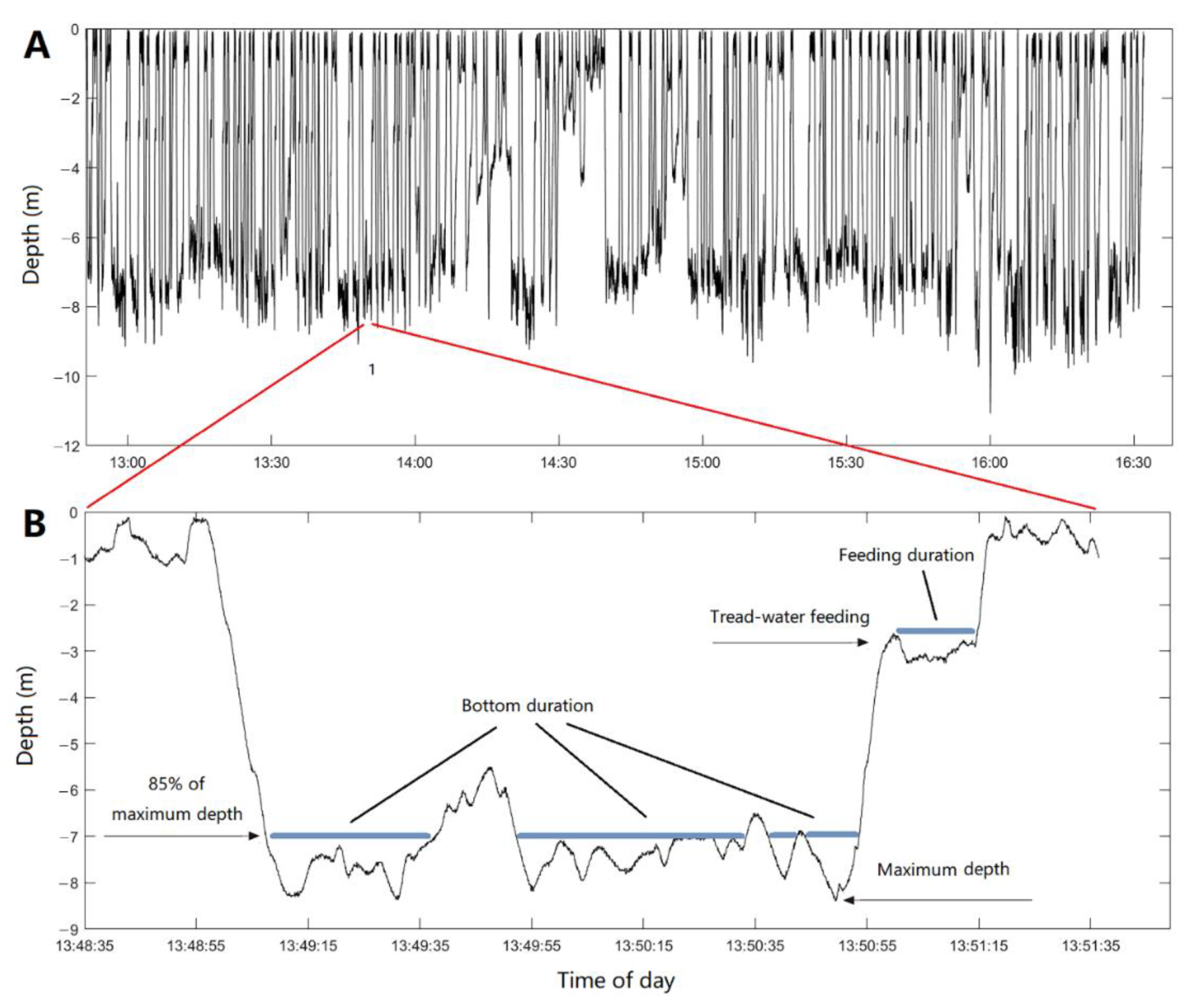
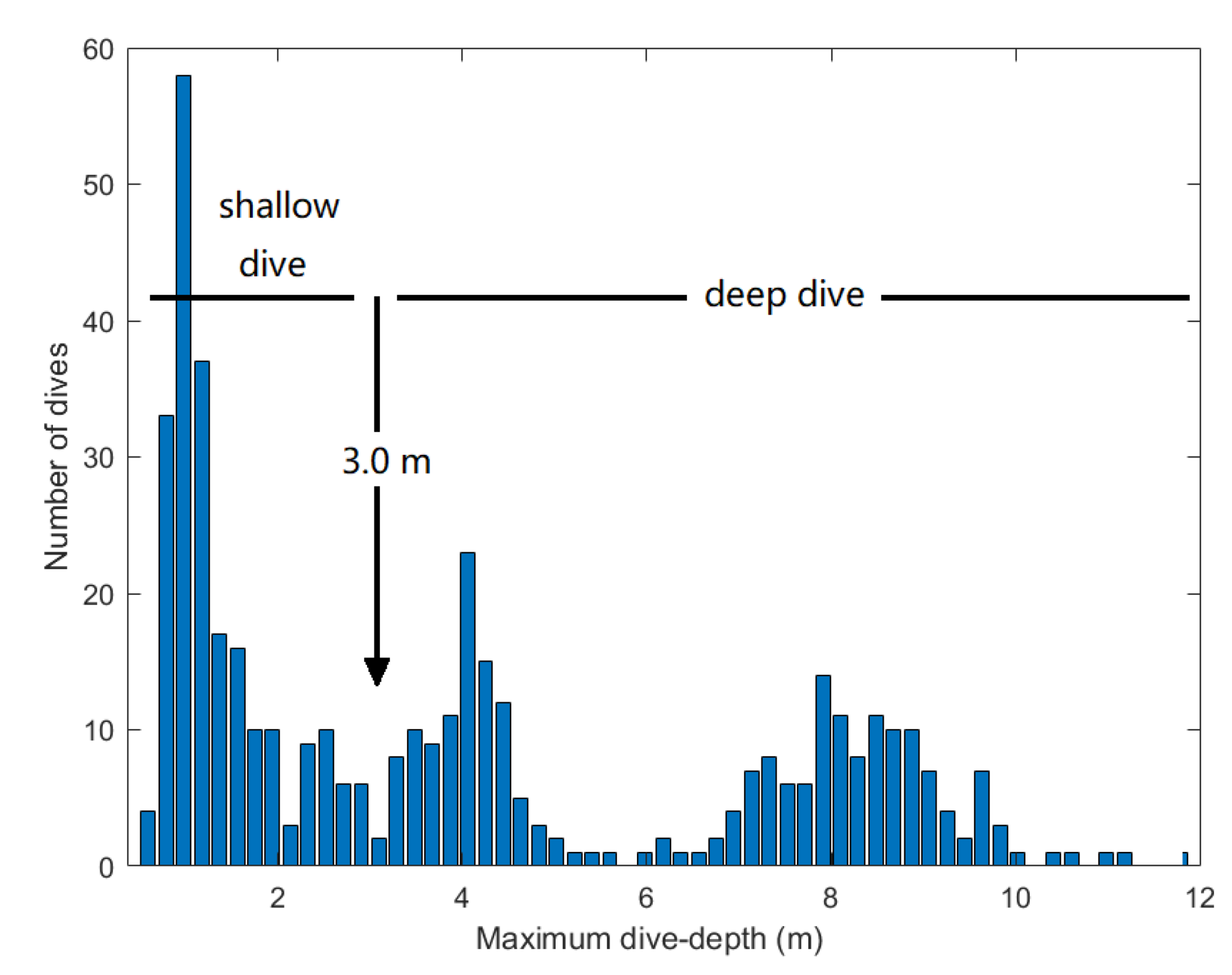
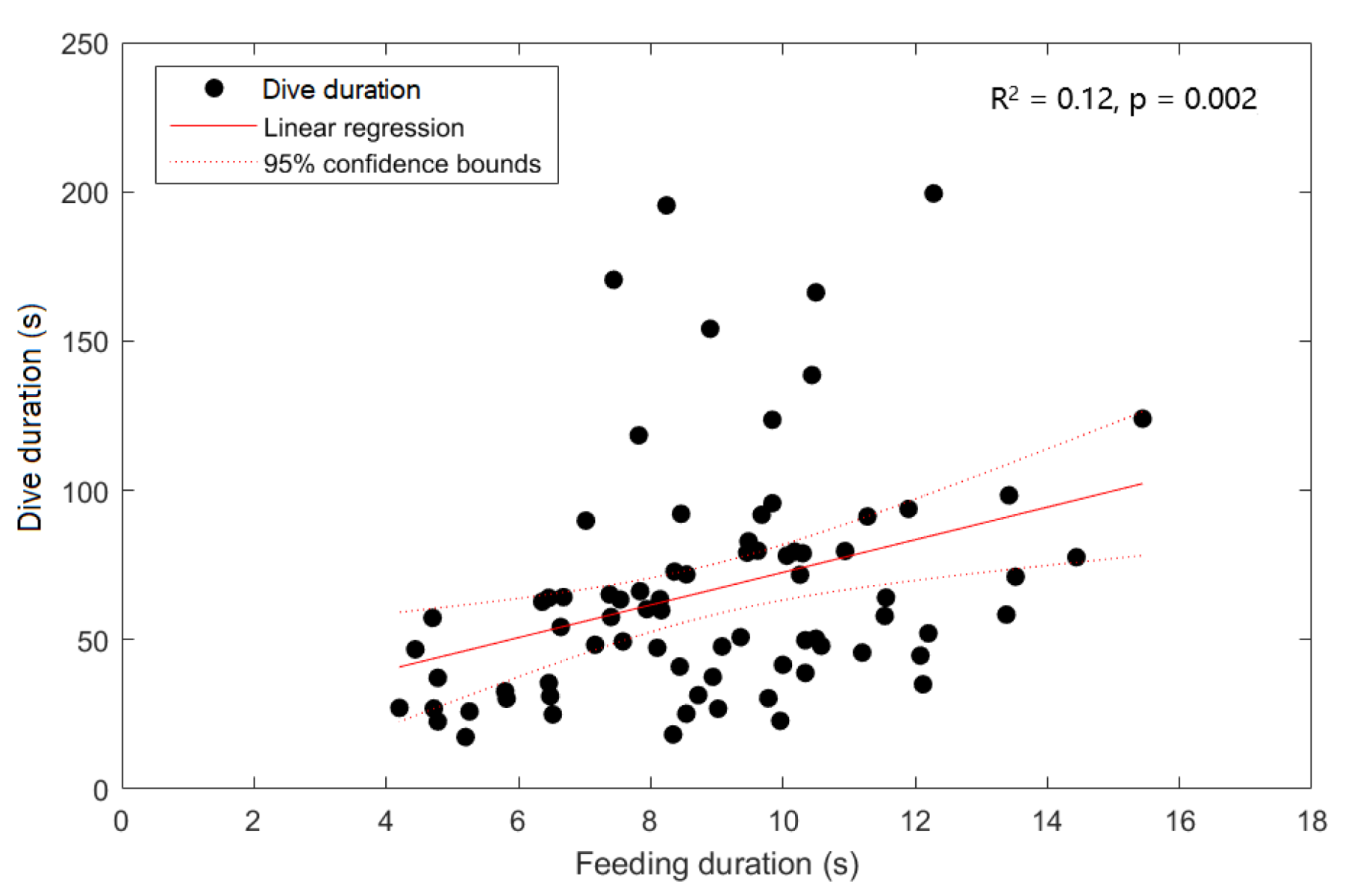
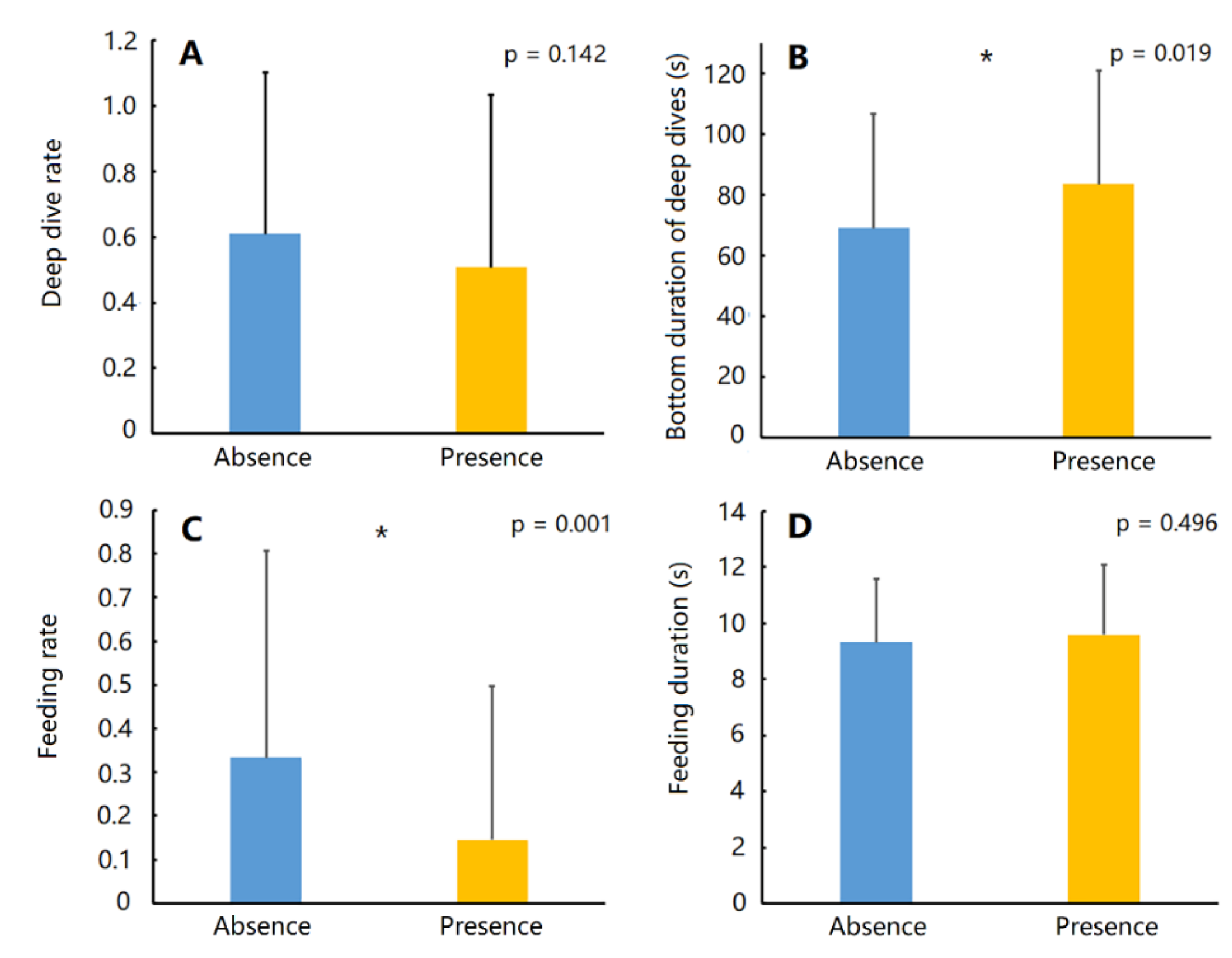
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boyd, I.L.; Kato, A.; Ropert-Coudert, Y. Bio-logging science: Sensing beyond the boundaries. Mem. Natl. Inst. Polar Res. 2004, 58, 1–14. [Google Scholar]
- Grothues, T.M. A review of acoustic telemetry technology and a perspective on its diversification relative to coastal tracking arrays. In Tagging and Tracking of Marine Animals with Electronic Devices; Nielsen, J.L., Ed.; Springer Science & Business Media: Heidelberg, Germany, 2009; pp. 77–90. [Google Scholar]
- Rutz, C.; Hays, G.C. New frontiers in biologging science. Biol. Lett. 2009, 5, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Hammerschlag, N.; Gallagher, A.; Lazarre, D. A review of shark satellite tagging studies. J. Exp. Mar. Biol. Ecol. 2011, 398, 1–8. [Google Scholar] [CrossRef]
- Stone, G.; Schubel, J.; Tausig, H. Electronic marine animal tagging: New frontier in ocean science. Oceanography 1999, 12, 24–27. [Google Scholar] [CrossRef][Green Version]
- Bograd, S.J.; Block, B.A.; Costa, D.P.; Godley, B.J. Biologging technologies: New tools for conservation. Introduction. Endang. Species Res. 2010, 10, 1–7. [Google Scholar] [CrossRef]
- Evans, K.; Lea, M.A.; Patterson, T.A. Recent advances in bio-logging science: Technologies and methods for understanding animal behaviour and physiology and their environments. Deep Sea Res. II Top. Stud. Oceanogr. 2013, 88–89, 1–6. [Google Scholar] [CrossRef]
- Baumgartner, M.F.; Hammar, T.; Robbins, J. Development and assessment of a new dermal attachment for short-term tagging studies of baleen whales. Methods Ecol. Evol. 2015, 6, 289–297. [Google Scholar] [CrossRef]
- Szesciorka, A.R.; Calambokidis, J.; Harvey, J.T. Testing tag attachments to increase the attachment duration of archival tags on baleen whales. Anim. Biotelemet. 2016, 4, 18. [Google Scholar] [CrossRef]
- Guzman, H.M.; Capella, J.J. Short-term recovery of humpback whales after percutaneous satellite tagging. J. Wildl. Manag. 2017, 81, 728–733. [Google Scholar] [CrossRef]
- Liu, M.; Lin, W.; Lin, M.; Liu, B.; Dong, L.; Zhang, P.; Yang, Z.; Wang, K.; Dai, L.; Li, S. The First Attempt of Satellite Tracking on Occurrence and Migration of Bryde’s Whale (Balaenoptera edeni) in the Beibu Gulf. J. Mar. Sci. Eng. 2021, 9, 796. [Google Scholar] [CrossRef]
- Tyson, R.B.; Friedlaender, A.S.; Ware, C.; Stimpert, A.K.; Nowacek, D.P. Synchronous mother and calf foraging behaviour in humpback whales Megaptera novaeangliae: Insights from multi-sensor suction cup tags. Mar. Ecol. Prog. Ser. 2012, 457, 209–220. [Google Scholar] [CrossRef]
- Goldbogen, J.A.; Calambokidis, J.; Friedlaender, A.S.; Francis, J.; DeRuiter, S.L.; Stimpert, A.K.; Falcone, E.; Southall, B.L. Underwater acrobatics by the world’s largest predator: 360 rolling manoeuvres by lunge-feeding blue whales. Biol. Lett. 2013, 9, 20120986. [Google Scholar] [CrossRef] [PubMed]
- Tackaberry, J.E.; Cade, D.E.; Goldbogen, J.A.; Wiley, D.N.; Friedlaender, A.S.; Stimpert, A.K. From a calf’s perspective: Humpback whale nursing behavior on two US feeding grounds. PeerJ 2020, 8, e8538. [Google Scholar] [CrossRef] [PubMed]
- Goldbogen, J.; Calambokidis, J.; Shadwick, R.; Oleson, E.; McDonald, M.; Hildebrand, J. Kinematics of diving and lunge-feeding in fin whales. J. Exp. Biol. 2006, 209, 1231–1244. [Google Scholar] [CrossRef]
- Chapple, T.K.; Gleiss, A.C.; Jewell, O.J.D.; Wikelski, M.; Block, B.A. Tracking sharks without teeth: A non-invasive rigid tag attachment for large predatory sharks. Anim. Biotelemet. 2015, 3, 14. [Google Scholar] [CrossRef]
- Cade, D.E.; Friedlaender, A.S.; Calambokidis, J.; Goldbogen, J.A. Kinematic Diversity in Rorqual Whale Feeding Mechanisms. Curr. Biol. 2016, 26, 2617–2624. [Google Scholar] [CrossRef]
- Caruso, F.; Hickmott, L.; Warren, J.D.; Segre, P.; Chiang, G.; Bahamonde, P.; ESPAÑOL-JIMÉNEZ, S.; Li, S.; Bocconcelli, A. Diel differences in blue whale (Balaenoptera musculus) dive behavior increase nighttime risk of ship strikes in northern Chilean Patagonia. Integr. Zool. 2020, 16, 594–611. [Google Scholar] [CrossRef]
- Meynecke, J.O.; Liebsch, N. Asset Tracking Whales—First Deployment of a Custom-Made GPS/GSM Suction Cup Tag on Migrating Humpback Whales. J. Mar. Sci. Eng. 2021, 9, 597. [Google Scholar] [CrossRef]
- Nelms, S.E.; Alfaro-Shigueto, J.; Arnould, J.P.; Avila, I.C.; Nash, S.B.; Campbell, E.; Carter, M.I.; Collins, T.; Currey, R.J.; Domit, C. Marine mammal conservation: Over the horizon. Endang. Species Res. 2021, 44, 291–325. [Google Scholar] [CrossRef]
- Kato, H.; Perrin, W.F. Bryde’s Whales: Balaenoptera edeni/brydei, in Encyclopedia of Marine Mammals; Elsevier: Burlington, MA, USA, 2009; pp. 158–163. [Google Scholar]
- Tezanos-Pinto, G.; Hupman, K.; Wiseman, N.; Dwyer, S.; Baker, C.; Brooks, L.; Outhwaite, B.; Lea, C.; Stockin, K. Local abundance, apparent survival and site fidelity of Bryde’s whales in the Hauraki Gulf (New Zealand) inferred from long-term photo-identification. Endang. Species Res. 2017, 34, 61–73. [Google Scholar] [CrossRef]
- Kershaw, F.; Leslie, M.S.; Collins, T.; Mansur, R.M.; Smith, B.D.; Minton, G.; Baldwin, R.; LeDuc, R.G.; Anderson, R.C.; Brownell Jr, R.L. Population differentiation of 2 forms of Bryde’s whales in the Indian and Pacific Oceans. J. Hered. 2013, 104, 755–764. [Google Scholar] [CrossRef]
- Cooke, J.G.; Brownell, R.L., Jr. Balaenoptera edeni. The IUCN Red List of Threatened Species 2018: ET2476A50349178. 2018. Available online: https://www.iucnredlist.org/species/2476/50349178 (accessed on 1 September 2022).
- Corkeron, P.; Reeves, R.; Rosel, P. Balaenoptera edeni (Gulf of MEXICO Subpopulation). The IUCN Red List of Threatened Species 2017; e.T117636167A117636174. 2017. Available online: https://www.iucnredlist.org/species/117636167/117636174 (accessed on 1 September 2022).
- Constantine, R.; Iwata, T.; Nieukirk, S.L.; Penry, G.S. Future directions in research on Bryde’s whales. Front. Mar. Sci. 2018, 5, 333. [Google Scholar] [CrossRef]
- Wang, P. Chinese Cetaceans; Chemical Industrial Press: Beijing, China, 2012. [Google Scholar]
- Parsons, E.; Felley, M.L.; Porter, L.J. An annotated checklist of cetaceans recorded from Hong Kong’s territorial waters. Asian Mar. Biol. 1995, 12, 79–100. [Google Scholar]
- Kato, H.; Shinohara, E.; Kishiro, T.; Noji, S. Distribution of Bryde’s whales off Kochi, southwest Japan, from the 1994/95 sighting survey. Rep. Int. Whaling. Comm. 1996, 46, 429–436. [Google Scholar]
- Jefferson, T.A.; Hung, S.K. An updated, annotated checklist of the marine mammals of Hong Kong. Mammalia 2007, 71, 105–114. [Google Scholar] [CrossRef]
- Liu, M.; Lin, M.; Li, S. Species diversity and spatiotemporal patterns based on cetacean stranding records in China, 1950–2018. Sci. Total Environ. 2022, 822, 153651. [Google Scholar] [CrossRef]
- Wada, S.; Oishi, M.; Yamada, T.K. A newly discovered species of living baleen whale. Nature 2003, 426, 278–281. [Google Scholar] [CrossRef]
- Chen, B.; Zhu, L.; Jefferson, T.; Zhou, K.; Yang, G. Coastal Bryde’s Whales’ (Balaenoptera edeni) Foraging Area Near Weizhou Island in the Beibu Gulf. Aquat. Mamm. 2019, 45, 274–279. [Google Scholar] [CrossRef]
- Chen, M.; Huang, S.-L.; Wu, H.; Mei, Z.; Chen, Y.; Zheng, J.; Cheng, W.; Deng, K.; Yang, C.; Mao, C.; et al. Occurrence of Bryde’s whales, Balaenoptera edeni, in the northern Beibu Gulf, China. Mar. Mamm. Sci. 2019, 35, 1643–1652. [Google Scholar] [CrossRef]
- Chantrapornsyl, S.; Adulyanukosol, K.; Kittiwathanawong, K. Records of cetaceans in Thailand. Phuket Mar. Biol. Res. Bull. 1996, 61, 39–63. [Google Scholar]
- Iwata, T.; Akamatsu, T.; Thongsukdee, S.; Cherdsukjai, P.; Adulyanukosol, K.; Sato, K. Tread-water feeding of Bryde’s whales. Curr. Biol. 2017, 27, R1154. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, M.; Mao, C.; Han, Y.; Hao, Y.; Wang, K.; Mei, Z.; Wang, D. Community-based population monitoring for large baleen whales: The case study of Bryde’s whale in Beibu Gulf of China. Integr. Zool. 2021, 16, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Cooke, S.J. Biotelemetry and biologging in endangered species research and animal conservation: Relevance to regional, national, and IUCN Red List threat assessments. Endang. Species Res. 2008, 4, 165–185. [Google Scholar] [CrossRef]
- Izadi, S.; Johnson, M.; de Soto, N.A.; Constantine, R. Night-life of Bryde’s whales: Ecological implications of resting in a baleen whale. Behav. Ecol. Sociobiol. 2018, 72, 78. [Google Scholar] [CrossRef]
- Croll, D.; Acevedo-Gutierrez, A.; Tershy, B. The diving behavior of blue and fin whales: Is dive duration shorter than expected based on oxygen stores? Comp. Biochem. Physiol. A 2001, 129, 797–809. [Google Scholar] [CrossRef]
- Akamatsu, T.; Wang, D.; Wang, K.; Wei, Z.; Zhao, Q.; Naito, Y. Diving behaviour of freshwater finless porpoises (Neophocaena phocaenoides) in an oxbow of the Yangtze River, China. ICES J. Mar. Sci. 2002, 59, 438–443. [Google Scholar] [CrossRef]
- Panigada, S.; Zanardelli, M.; Canese, S.; Jahoda, M. How deep can baleen whales dive? Mar. Ecol. Prog. Ser. 1999, 187, 309–311. [Google Scholar] [CrossRef]
- Alves, F.; Dinis, A.; Cascão, I.; Freitas, L. Bryde’s whale (Balaenoptera brydei) stable associations and dive profiles: New insights into foraging behavior. Mar. Mamm. Sci. 2010, 26, 202–212. [Google Scholar] [CrossRef]
- Soldevilla, M.S.; Hildebrand, J.A.; Frasier, K.E.; Dias, L.A.; Martinez, A.; Mullin, K.D.; Rosel, P.E.; Garrison, L.P. Spatial distribution and dive behavior of Gulf of Mexico Bryde’s whales: Potential risk of vessel strikes and fisheries interactions. Endang. Species Res. 2017, 32, 533–550. [Google Scholar] [CrossRef]
- Goldbogen, J.A.; Friedlaender, A.S.; Calambokidis, J.; McKenna, M.F.; Simon, M.; Nowacek, D.P. Integrative approaches to the study of baleen whale diving behavior, feeding performance, and foraging ecology. Bioscience 2013, 63, 90–100. [Google Scholar] [CrossRef]
- Friedlaender, A.; Goldbogen, J.; Nowacek, D.; Read, A.; Johnston, D.; Gales, N. Feeding rates and under-ice foraging strategies of the smallest lunge filter feeder, the Antarctic minke whale (Balaenoptera bonaerensis). J. Exp. Biol. 2014, 217, 2851–2854. [Google Scholar] [CrossRef] [PubMed]
- Cade, D.E.; Seakamela, S.M.; Findlay, K.P.; Fukunaga, J.; Kahane-Rapport, S.R.; Warren, J.D.; Calambokidis, J.; Fahlbusch, J.A.; Friedlaender, A.S.; Hazen, E.L. Predator-scale spatial analysis of intra-patch prey distribution reveals the energetic drivers of rorqual whale super-group formation. Funct. Ecol. 2021, 35, 894–908. [Google Scholar] [CrossRef]
- Villagra, D.; García-Cegarra, A.; Gallardo, D.I.; Pacheco, A.S. Energetic effects of whale-watching boats on humpback whales on a breeding ground. Front. Mar. Sci. 2021, 7, 600508. [Google Scholar] [CrossRef]
- Schaffar, A.; Madon, B.; Garrigue, C.; Constantine, R. Behavioural effects of whale-watching activities on an Endangered population of humpback whales wintering in New Caledonia. Endang. Species Res. 2013, 19, 245–254. [Google Scholar] [CrossRef]
- Di Clemente, J.; Christiansen, F.; Pirotta, E.; Steckler, D.; Wahlberg, M.; Pearson, H.C. Effects of whale watching on the activity budgets of humpback whales, Megaptera novaeangliae (Borowski, 1781), on a feeding ground. Aquat. Conserv. Mar. Freshwat. Ecosyst. 2018, 28, 810–820. [Google Scholar] [CrossRef]
- Sprogis, K.R.; Bejder, L.; Hanf, D.; Christiansen, F. Behavioural responses of migrating humpback whales to swim-with-whale activities in the Ningaloo Marine Park, Western Australia. J. Exp. Mar. Biol. Ecol. 2020, 522, 151254. [Google Scholar] [CrossRef]
- Hooker, S.K.; Baird, R.W. Diving and ranging behaviour of odontocetes: A methodological review and critique. Mamm. Rev. 2001, 31, 81–105. [Google Scholar] [CrossRef]
- Cade, D.E.; Barr, K.R.; Calambokidis, J.; Friedlaender, A.S.; Goldbogen, J.A. Determining forward speed from accelerometer jiggle in aquatic environments. J. Exp. Biol. 2018, 221, jeb170449. [Google Scholar] [CrossRef]
- Miller, P.J.; Johnson, M.P.; Tyack, P.L.; Terray, E.A. Swimming gaits, passive drag and buoyancy of diving sperm whales Physeter macrocephalus. J. Exp. Biol. 2004, 207, 1953–1967. [Google Scholar] [CrossRef]
- Stimpert, A.; Mattila, D.; Nosal, E.-M.; Au, W. Tagging young humpback whale calves: Methodology and diving behavior. Endang. Species Res. 2012, 19, 11–17. [Google Scholar] [CrossRef][Green Version]
- Keen, E.; Falcone, E.; Andrews, R.; Schorr, G. Diel Dive Behavior of Fin Whales (Balaenoptera physalus) in the Southern California Bight. Aquat. Mamm. 2019, 45, 233–243. [Google Scholar] [CrossRef]
- Irvine, B. Behavioral changes in dolphins in a strange environment. Q. J. Fla. Acad. Sci. 1971, 34, 206–212. [Google Scholar]
- Rheingold, H.L.; Eckerman, C.O. Familiar social and nonsocial stimuli and the kitten’s response to a strange environment. Dev. Psychobiol.: J. Int. Soc. Dev. Psychobiol. 1971, 4, 71–89. [Google Scholar] [CrossRef]
- Kummrow, M. Diagnostic and therapeutic guidelines to abnormal behavior in captive nonhuman primates. Vet. Clin. Exot. Anim. 2021, 24, 253–266. [Google Scholar] [CrossRef]
- Mate, B.R.; Lagerquist, B.A.; Calambokidis, J. Movements of North Pacific Blue whales during the feeding season off Southern California and their southern fall migration. Mar. Mamm. Sci. 1999, 15, 1246–1257. [Google Scholar] [CrossRef]
- Murase, H.; Tamura, T.; Otani, S.; Nishiwaki, S. Satellite tracking of Bryde’s whales Balaenoptera edeni in the offshore western North Pacific in summer 2006 and 2008. Fish. Sci. 2016, 82, 35–45. [Google Scholar] [CrossRef]
- Bordes, F.; Morand, S. The impact of multiple infections on wild animal hosts: A review. Infect. Ecol. Epidemiol. 2011, 1, 7346. [Google Scholar] [CrossRef]
- Ghai, R.R.; Fugere, V.; Chapman, C.A.; Goldberg, T.L.; Davies, T.J. Sickness behaviour associated with non-lethal infections in wild primates. Proc. R. Soc. B 2015, 282, 20151436. [Google Scholar] [CrossRef]
- Bohn, S.J.; Turner, J.M.; Warnecke, L.; Mayo, C.; McGuire, L.; Misra, V.; Bollinger, T.K.; Willis, C.K. Evidence of ‘sickness behaviour’in bats with white-nose syndrome. Behaviour 2016, 153, 981–1003. [Google Scholar] [CrossRef]
- Hamilton, D.G.; Jones, M.E.; Cameron, E.Z.; Kerlin, D.H.; McCallum, H.; Storfer, A.; Hohenlohe, P.A.; Hamede, R.K. Infectious disease and sickness behaviour: Tumour progression affects interaction patterns and social network structure in wild Tasmanian devils. Proc. R. Soc. B 2020, 287, 20202454. [Google Scholar] [CrossRef]
- Blersch, R.; Bonnell, T.R.; Ganswindt, A.; Young, C.; Barrett, L.; Henzi, S.P. Sick and tired: Sickness behaviour, polyparasitism and food stress in a gregarious mammal. Behav. Ecol. Sociobiol. 2021, 75, 169. [Google Scholar] [CrossRef]
- Mayer, J. Use of behavior analysis to recognize pain in small mammals. Lab. Anim. 2007, 36, 43–48. [Google Scholar] [CrossRef]
- Long, X.; Chen, P.; Yuan, H.; Feng, X.; Yu, J.; Shu, L.; Li, X.; Chen, W.; Chen, Z. Evaluation on carrying capacity of fishery resources in coastal waters of Dapeng Peninsula, Shenzhen. South China Fish. Sci. 2019, 15, 56–64. [Google Scholar]
- Zhang, J.; Qiu, J.-W.; Chen, C.-L.; Zeng, J.-W.; Wang, X.-F. Analysis of the Fish Community Structure and Its Relationship with Environmental Factors in Autumn and Spring in Northern Dapeng Bay, Guangdong. J. Guangdong Ocean. Univ. 2020, 40, 43–52. [Google Scholar]
- Penry, G.S.; Cockcroft, V.G.; Hammond, P.S. Seasonal fluctuations in occurrence of inshore Bryde’s whales in Plettenberg Bay, South Africa, with notes on feeding and multispecies associations. Afr. J. Mar. Sci. 2011, 33, 403–414. [Google Scholar] [CrossRef]
- Wiseman, N.; Parsons, S.; Stockin, K.A.; Baker, C.S. Seasonal occurrence and distribution of Bryde’s whales in the Hauraki Gulf, New Zealand. Mar. Mamm. Sci. 2011, 27, E253–E267. [Google Scholar] [CrossRef]
- Maciel, I.S.; Tardin, R.H.; Simão, S.M. Occurrence and habitat use of Bryde’s whales (Balaenoptera edeni) in the Cabo Frio region, South-eastern Brazil. J. Mar. Biol. Assoc. UK 2018, 98, 1081–1086. [Google Scholar] [CrossRef]
- Jia, C.; Zhuang, S. Current situation and evaluation of fishery resources in the east coast of Shenzhen city. China Fish. 2009, 8, 21–23. [Google Scholar]
- Yoshida, H.; Kato, H. Phylogenetic relationships of Bryde’s whales in theWestern North Pacific and adjacent waters inferred from mitochondrial DNA sequences. Mar. Mamm. Sci. 1999, 15, 1269–1286. [Google Scholar] [CrossRef]
- Kanda, N.; Goto, M.; Kato, H.; McPhee, M.V.; Pastene, L.A. Population genetic structure of Bryde’s whales (Balaenoptera brydei) at the inter-oceanic and trans-equatorial levels. Conserv. Genet. 2007, 8, 853–864. [Google Scholar] [CrossRef]
- Erbe, C. Underwater Acoustics: Noise and the Effects on Marine Mammal—A Pocket Handbook, 3rd ed.; JASCO Applied Sciences: Capalaba, Australia, 2011; Volume 164, p. 64. [Google Scholar]
- Popper, A.N.; Hawkins, A. The Effects of Noise on Aquatic Life; Springer Science & Business Media: Heidelberg, Germany, 2012; Volume 730. [Google Scholar]
- Finneran, J.J. Noise-induced hearing loss in marine mammals: A review of temporary threshold shift studies from 1996 to 2015. J. Acoust. Soc. Am. 2015, 138, 1702–1726. [Google Scholar] [CrossRef]
- Christiansen, F.; Rasmussen, M.; Lusseau, D. Whale watching disrupts feeding activities of minke whales on a feeding ground. Mar. Ecol. Prog. Ser. 2013, 478, 239–251. [Google Scholar] [CrossRef]
- Enrico, P.; Booth, C.G.; Costa, D.P.; Fleishman, E.; Kraus, S.D.; Lusseau, D.; Moretti, D.; New, L.F.; Schick, R.S.; Schwarz, L.K.; et al. Understanding the population consequences of disturbance. Ecol. Evol. 2018, 8, 9934–9946. [Google Scholar]
- Li, S.; Wu, H.; Xu, Y.; Peng, C.; Fang, L.; Lin, M.; Xing, L.; Zhang, P. Mid- to high-frequency noise from high-speed boats and its potential impacts on humpback dolphins. J. Acoust. Soc. Am. 2015, 138, 942–952. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Tag Deployment Time (GMT + 8) | Tag Retrieve Time (GMT + 8) | Deployment Location | Retrieve Location | Track Duration (min) | Total Numbers of Dives | Total Numbers of Feeding |
|---|---|---|---|---|---|---|---|
| 2 July 2021 | 12:51:16 | 16:32:14 | 22°35.681’ N 114°25.279’ E | 22°35.686’ N 114°25.551’ E | 220.97 | 278 | 45 |
| 4 July 2021 | 6:38:37 | 9:32:10 | 22°35.740’ N 114°25.473’ E | 22°35.478’ N 114°25.830’ E | 173.55 | 177 | 32 |
| Total | —— | —— | —— | —— | 394.52 | 455 | 77 |
| All Dives (n = 455) | Deep Dives (n = 238) | Shallow Dives (n = 217) | |
|---|---|---|---|
| Max. depth (m) | 4.03 ± 3.17 | 6.49 ± 2.48 | 1.33 ± 0.61 |
| Dive duration (s) | 46.52 ± 39.89 | 72.20 ± 38.71 | 18.36 ± 13.30 |
| Bottom duration (s) | 24.57 ± 30.67 | 38.11 ± 34.45 | 9.73 ± 15.90 |
| Max speed (m/s) | 2.40 ± 1.30 | 2.84 ± 1.52 | 1.91 ± 0.74 |
| Mean speed (m/s) | 0.61 ± 0.18 | 0.67 ± 0.15 | 0.53 ± 0.17 |
| Degree of descent angle | 29.35 ± 12.61 | 34.42 ± 13.60 | 23.77 ± 8.46 |
| Degree of ascent angle | 18.77 ± 14.33 | 25.50 ± 15.73 | 11.36 ± 6.97 |
| Descending rate (m/s) | 0.37 ± 0.19 | 0.44 ± 0.19 | 0.29 ± 0.15 |
| Ascending rate (m/s) | 0.49 ± 0.37 | 0.45 ± 0.20 | 0.53 ± 0.49 |
| Number of Tread-water feeding events | 77 | 77 | 0 |
| Statistics | With Feeding Behaviours (n = 77) | Without Feeding Behaviours (n = 161) | p Value | |
|---|---|---|---|---|
| Dive duration (s) | 66.59 ± 39.51 | 76.19 ± 37.78 | 0.015 * | |
| Descent | Duration (s) | 15.67 ± 7.88 | 18.16 ± 13.24 | 0.100 |
| Averaged rate (m/s) | 0.50 ± 0.21 | 0.42 ± 0.17 | 0.005 * | |
| Bottom | Duration (s) | 40.47 ± 38.84 | 36.98 ± 32.20 | 0.810 |
| Maximum depth (m) | 6.71 ± 2.19 | 6.39 ± 2.61 | 0.041 * | |
| Ascent | Duration (s) | 10.45 ± 3.37 | 21.05 ± 16.98 | 0.000 * |
| Averaged rate (m/s) | 0.55 ± 0.15 | 0.40 ± 0.21 | 0.000 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Liu, M.; Lin, W.; Li, S. First Suction Cup Tagging on a Small and Coastal Form Bryde’s Whale (Balaenoptera edeni edeni) in China to Investigate Its Dive Profiles and Foraging Behaviours. J. Mar. Sci. Eng. 2022, 10, 1422. https://doi.org/10.3390/jmse10101422
Dong L, Liu M, Lin W, Li S. First Suction Cup Tagging on a Small and Coastal Form Bryde’s Whale (Balaenoptera edeni edeni) in China to Investigate Its Dive Profiles and Foraging Behaviours. Journal of Marine Science and Engineering. 2022; 10(10):1422. https://doi.org/10.3390/jmse10101422
Chicago/Turabian StyleDong, Lijun, Mingming Liu, Wenzhi Lin, and Songhai Li. 2022. "First Suction Cup Tagging on a Small and Coastal Form Bryde’s Whale (Balaenoptera edeni edeni) in China to Investigate Its Dive Profiles and Foraging Behaviours" Journal of Marine Science and Engineering 10, no. 10: 1422. https://doi.org/10.3390/jmse10101422
APA StyleDong, L., Liu, M., Lin, W., & Li, S. (2022). First Suction Cup Tagging on a Small and Coastal Form Bryde’s Whale (Balaenoptera edeni edeni) in China to Investigate Its Dive Profiles and Foraging Behaviours. Journal of Marine Science and Engineering, 10(10), 1422. https://doi.org/10.3390/jmse10101422