
Age and Growth of the Spot-Tail Shark, Carcharhinus sorrah, in the Taiwan Strait


Abstract
:1. Introduction
2. Materials and Methods
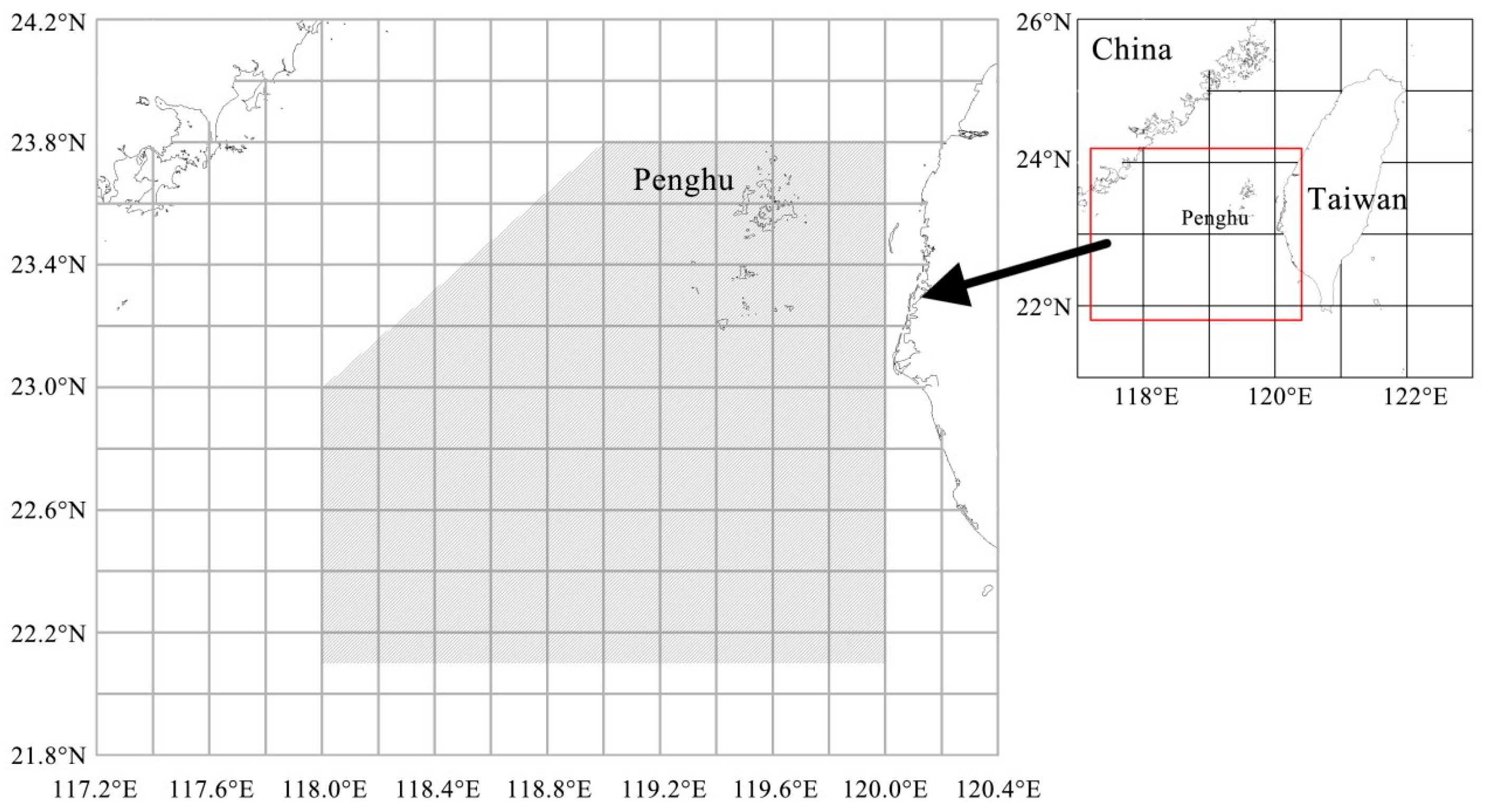
2.1. Specimen Collection and Processing
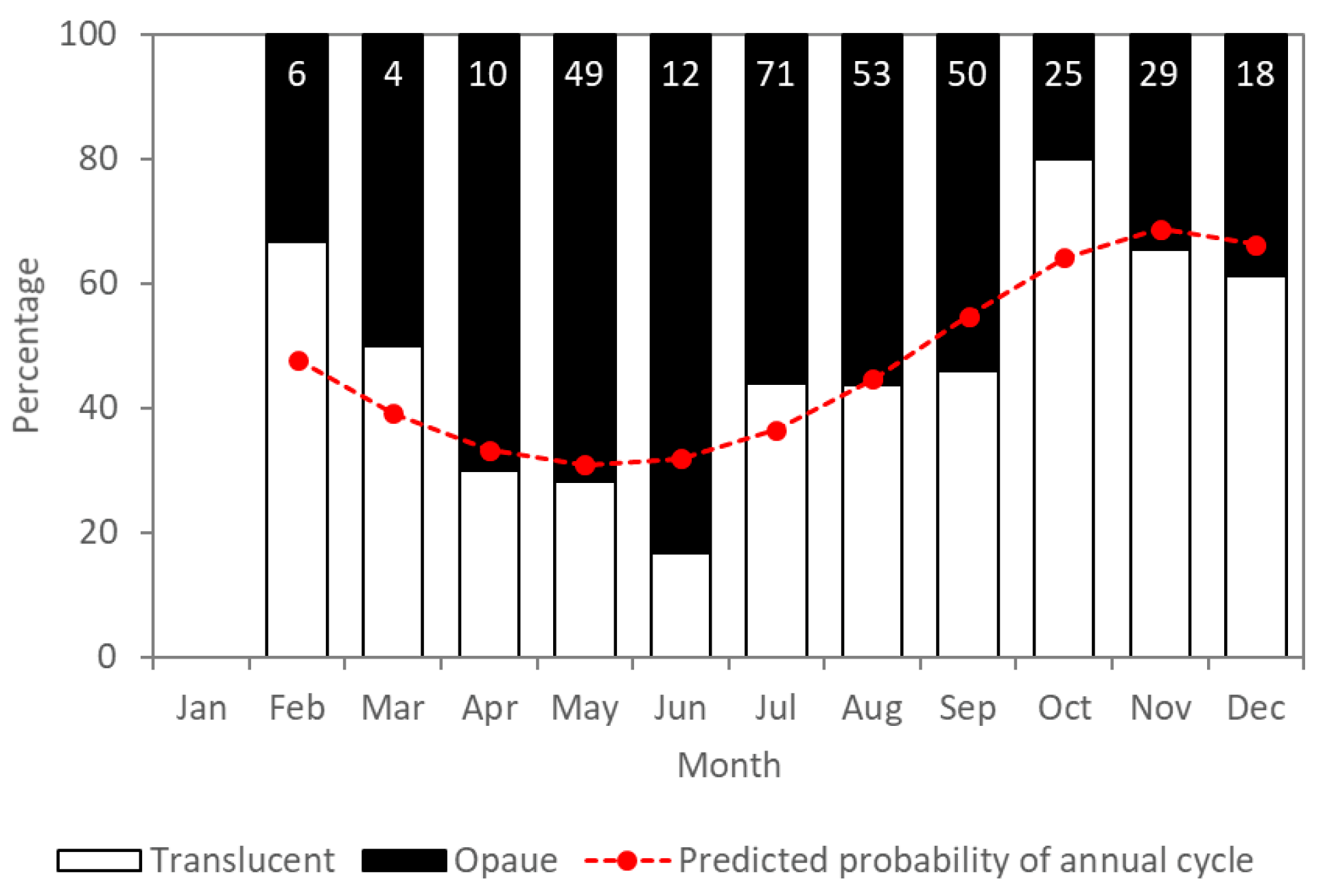
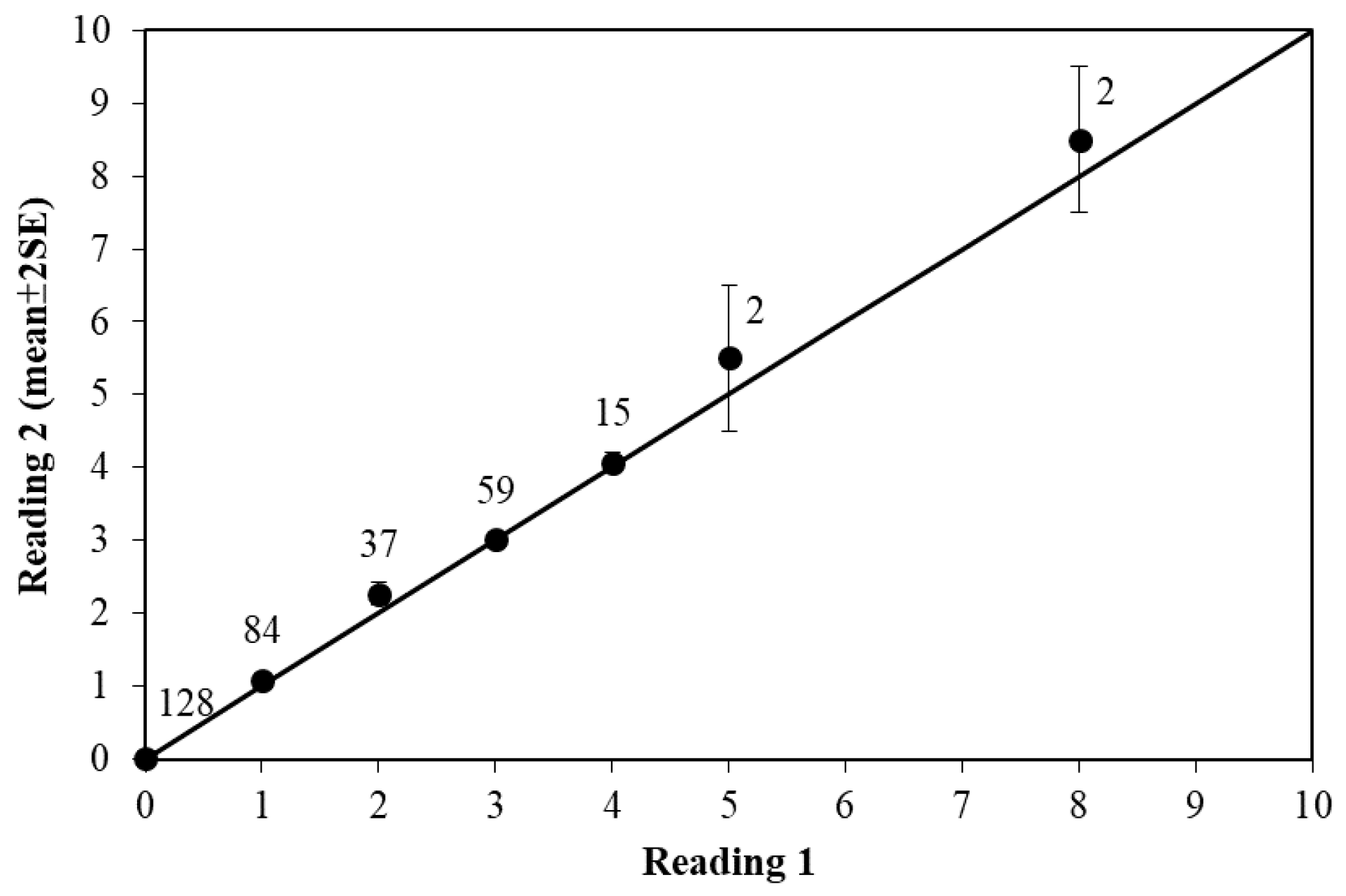
2.2. Age Estimation and Verification
2.3. Growth Functions
3. Results
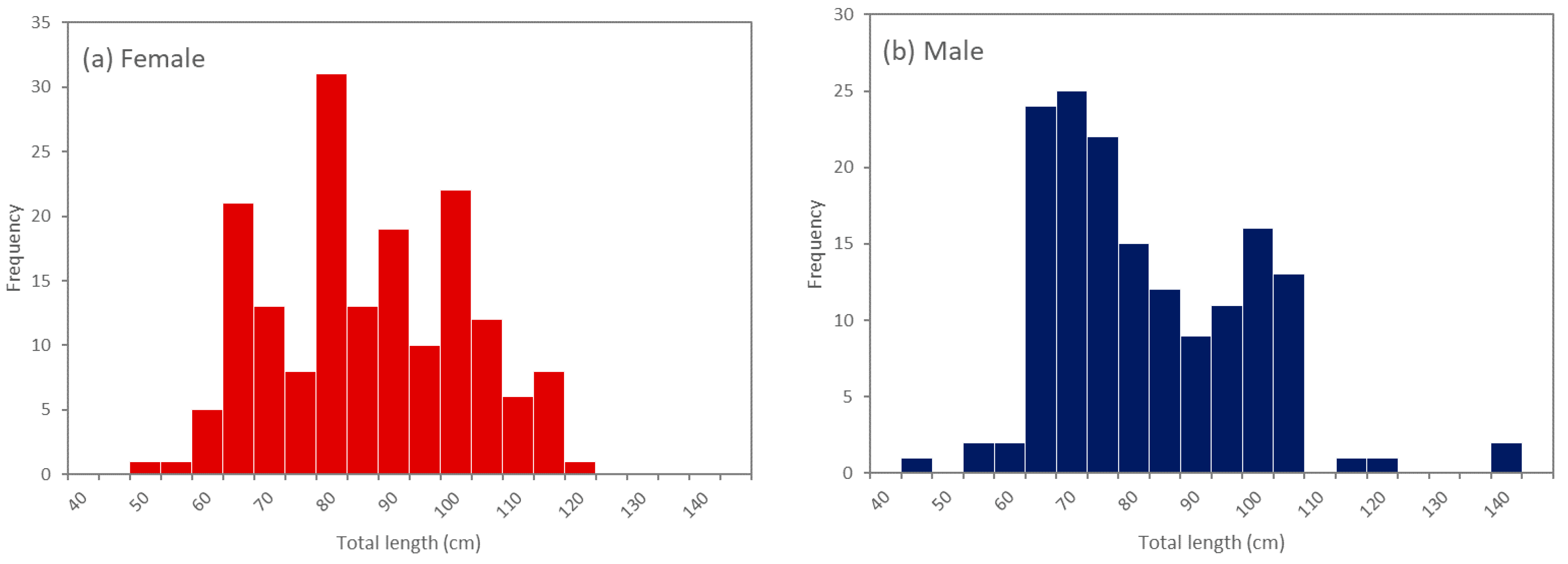
3.1. Size Range of Specimens
3.2. Meristic Relationships
3.3. TL–CD and W–TL Relationships
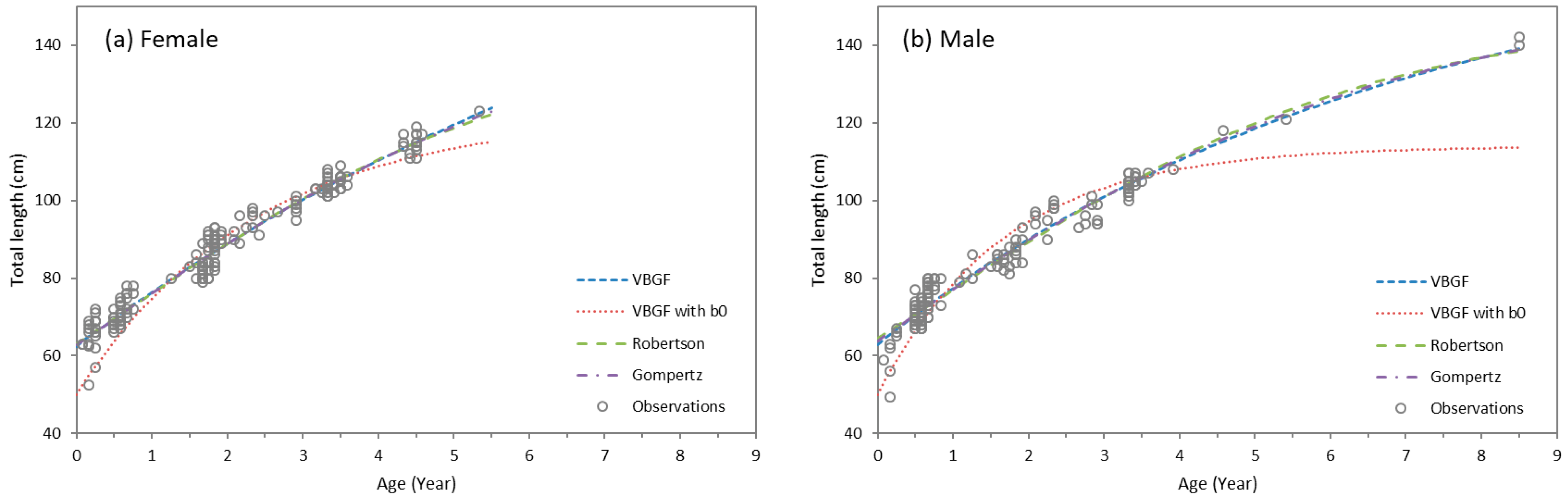
3.4. Growth Parameters
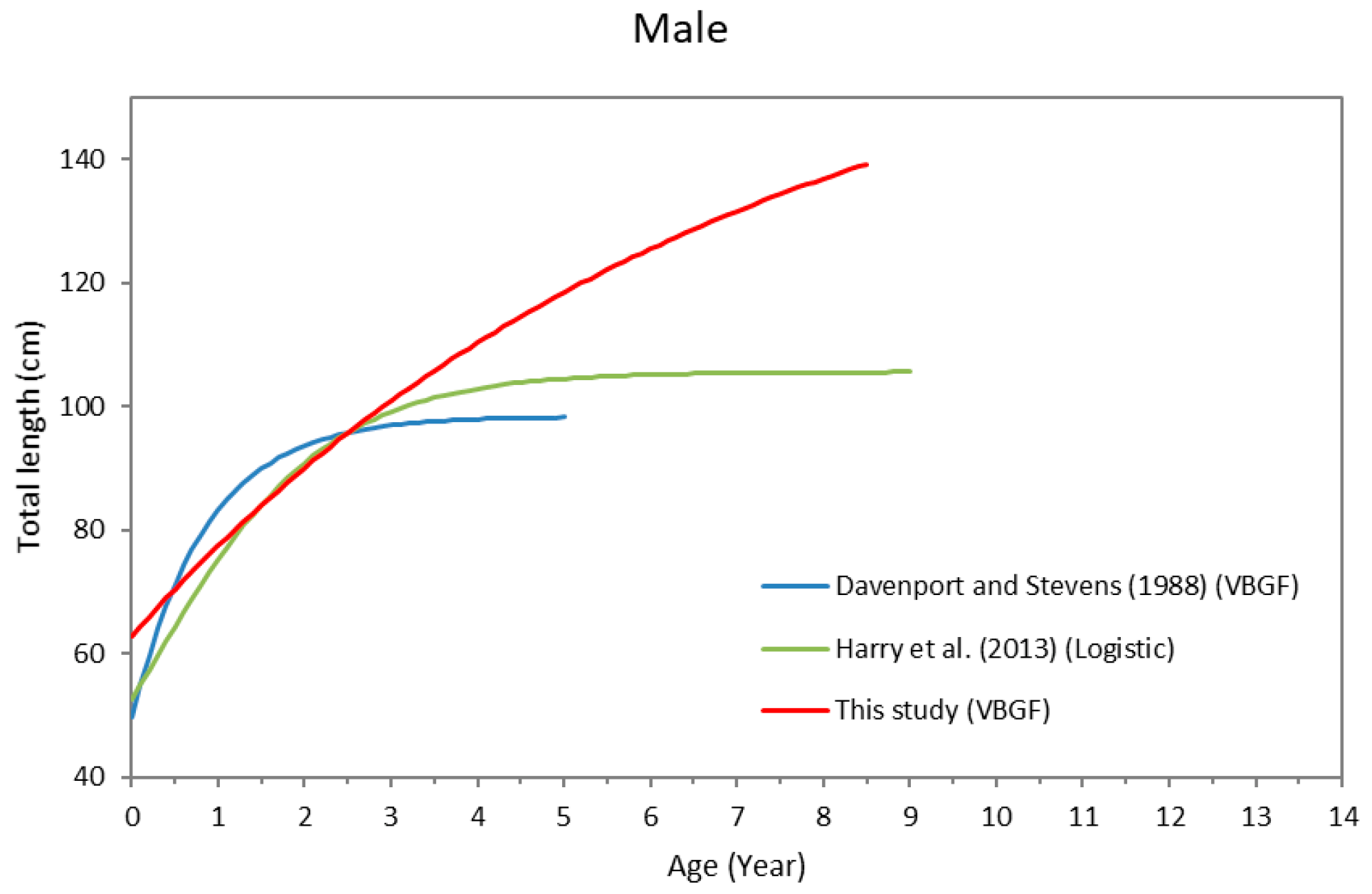
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fordham, S. Conservation and management status of spiny dogfish sharks (Squalus acanthias). AC20 Inf. 2004, 22, 1–7. [Google Scholar]
- Kruse, G.H.; Gallucci, V.F.; Tribuzio, C.A.; Rice, J. Abundance, Life History, and Population Demographics of Spiny Dogfish, Squalus Acanthias, in the Gulf of Alaska; School of Fisheries and Ocean Sciences, University of Alaska Fairbanks: Fairbanks, AK, USA, 2008; p. 173. [Google Scholar]
- Reynolds, J.D.; Dulvy, N.K.; Goodwin, N.B.; Hutchings, J.A. Biology of extinction risk in marine fishes. Proc. R. Soc. B Biol. Sci. 2005, 272, 2337–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anon. Taiwan Fisheries Statistic Year Book, Fisheries Agency, Council of Agriculture, Taiwan. 2014. Available online: https://www.fa.gov.tw/cht/PublicationsFishYear/ (accessed on 24 June 2015).
- Compagno, L.J.V. FAO species catalogue. Vol. 4. Sharks of the World. An annoted and illustrated catalogue of shark species known to date. Part 2. Carcharhiniformes. FAO Fish. Synop. 1984, 125, 251–655. [Google Scholar]
- Shao, K.T. The Fish Database of Taiwan. WWW Web Electronic Publication. 2018. Available online: http://fishdb.sinica.edu.tw (accessed on 20 December 2018).
- Compagno, L.J.V.; Dando, M.; Fowler, S. Sharks of the World: Collins Field Guide; Harper Collins Publishers: London, UK, 2005; p. 368. [Google Scholar]
- Giles, J.L.; Ovenden, J.R.; AlMojil, D.; Garvilles, E.; Khampetch, K.O.; Manjebrayakath, H.; Riginos, C. Extensive genetic population structure in the Indo–West Pacific spot-tail shark, Carcharhinus sorrah. Bull. Mar. Sci. 2014, 90, 427–454. [Google Scholar] [CrossRef] [Green Version]
- Stevens, J.D.; Wiley, P.D. Biology of two commercially important Carcharhinid sharks from northern Australia. Mar. Freshw. Res. 1986, 37, 671–688. [Google Scholar] [CrossRef]
- Davenport, S.; Stevens, J.D. Age and growth of two commercially important sharks (Carcharhinus tilstoni and C. sorrah) from Northern Australia. Mar. Freshw. Res. 1988, 39, 417–433. [Google Scholar] [CrossRef]
- White, W.T. Catch composition and reproductive biology of whaler sharks (Carcharhiniformes: Carcharhinidae) caught by fisheries in Indonesia. J. Fish. Biol. 2007, 71, 1512–1540. [Google Scholar] [CrossRef]
- Harry, A.V. Evidence for systemic age underestimation in shark and ray ageing studies. Fish Fish. 2018, 19, 185–200. [Google Scholar] [CrossRef]
- Pillans, R.; Stevens, J.D.; White, W.T. Carcharhinus sorrah. The IUCN Red List of Threatened Species 2009: E.T161376A5409506. Retrieved 2009, 10, 2009-2. [Google Scholar]
- Cailliet, G.M.; Goldman, K.J. Age determination and validation in chondrichthyan fishes. In Biology of Sharks and Their Relatives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice-Hall Pearson: Hoboken, NJ, USA, 2010. [Google Scholar]
- Kimura, D.K. Likelihood models for the von Bertalanffy growth curve. Fish. Bull. 1980, 77, 765–776. [Google Scholar]
- Joung, S.J.; Liao, Y.Y.; Chen, C.T. Age and growth of sandbar shark, Carcharhinus plumbeus, in northeastern Taiwan waters. Fish. Res. 2004, 70, 83–96. [Google Scholar] [CrossRef]
- Chen, C.T.; Leu, T.C.; Joung, S.J.; Lo, N.C.H. Age and growth of the scalloped hammerhead, Sphyrna lewini, in northeastern Taiwan waters. Pac. Sci. 1990, 44, 156–170. [Google Scholar]
- Okamura, H.; Semba, Y. A novel statistical method for validating the periodicity of vertebral growth band formation in elasmobranch fishes. Can. J. Fish. Aquat. Sci. 2009, 66, 771–780. [Google Scholar] [CrossRef]
- Hurvich, C.M.; Tsai, C.L. Regression and time series model selection in small samples. Biometrika 1989, 76, 297–307. [Google Scholar] [CrossRef]
- Beverton, R.J.H. Notes on the Use of Theoretical Models in the Study of the Dynamics of Exploited Fish Populations; Miscellaneous contributions, US Fishery Laboratory, number 2; Bureau of Commercial Fisheries: Beaufort, NC, USA, 1954. [Google Scholar]
- Fabens, A.J. Properties and fitting of the von Bertalanffy growth curve. Growth 1965, 29, 265–289. [Google Scholar]
- Robertson, T.B. The Chemical Basis of Growth and Senescence; Lippincott, J.B., Ed.; JB Lippincott Company: Philadelphia, PA, USA, 1923; p. 98. [Google Scholar]
- Gompertz, B. On the nature of the function expressive of the law of human mortality and on a new mode of determining life contingencies. Philos. Trans. R. Soc. Lond.–A Math. Phys. Sci. 1825, 115, 513–583. [Google Scholar] [CrossRef]
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In Second International Symposium on Information Theory; Petrov, B.N., Csaki, F., Eds.; Akademiai Kiado: Budapest, Hungary, 1973; pp. 267–281. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information–Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Harry, A.V.; Tobin, A.J.; Simpfendorfer, C.A. Age, growth and reproductive biology of the spot-tail shark, Carcharhinus sorrah, and the Australian blacktip shark, C. tilstoni, from the Great Barrier Reef World Heritage Area, northeastern Australia. Mar. Freshw. Res. 2013, 64, 277–293. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. (Eds.) FishBase, version (08/2021); World Wide Web Electronic Publication, 2021. Available online: https://www.fishbase.se/search.php (accessed on 1 August 2021).
- Joung, S.J.; Liao, Y.Y.; Liu, K.M.; Chen, C.T.; Leu, L.C. Age, growth, and reproduction of the spinner shark, Carcharhinus brevipinna, in the northeastern waters of Taiwan. Zool. Stud. 2005, 44, 102–110. [Google Scholar]
- Joung, S.J.; Chen, C.T.; Lee, H.H.; Liu, K.M. Age, growth, and reproduction of the silky sharks Carcharhinus falciformis in northeastern Taiwan waters. Fish. Res. 2008, 90, 78–85. [Google Scholar] [CrossRef]
- Joung, S.J.; Chen, J.H.; Chin, C.P.; Liu, K.M. Age and growth of the dusky shark, Carcharhinus obscurus, in the Northwest Pacific Ocean. Terr. Atmos. Ocean. Sci. 2015, 26, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Joung, S.J.; Chen, N.F.; Hsu, H.H.; Liu, K.M. Estimates of life history parameters of the oceanic whitetip shark, Carcharhinus longimanus, in the western North Pacific Ocean. Mar. Biol. Res. 2016, 12, 758–768. [Google Scholar] [CrossRef]
- Ocean Data Bank of Taiwan, WWW Web Electronic Publication. 2017. Available online: https://www.odb.ntu.edu.tw (accessed on 5 December 2017).
- Natanson, L.J.; Skomal, G.; Hoffmann, S.; Porter, M.E.; Goldman, K.J.; Serra, D. Age and growth of sharks: Do vertebral band pairs record age? Mar. Freshw. Res. 2018, 69, 1440–1452. [Google Scholar] [CrossRef]
- Francis, M.; Campana, S.E.; Jones, C.M. Age under-estimation in New Zealand porbeagle sharks (Lamna nasus): Is there an upper limit to ages that can be determined from shark vertebrae? Mar. Freshw. Res. 2007, 58, 10–23. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.M.; Wu, C.B.; Joung, S.J.; Tsai, W.P.; Su, K.Y. Multi-model approach on growth estimation and association with life history trait for elasmobranchs. Front. Mar. Sci. 2021, 8, 591692. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Female | Male | ||||
|---|---|---|---|---|---|
| Month | n | Rage of TL (cm) | n | Rage of TL (cm) | Total |
| January | - | - | - | - | - |
| February | 2 | 85.0–88.0 | 4 | 79–97 | 6 |
| March | 4 | 86.0–103.0 | - | - | 4 |
| April | 4 | 74.0–103.0 | 6 | 80–108 | 10 |
| May | 26 | 63.0–123.0 | 23 | 59–107 | 49 |
| June | 5 | 88.0–112.0 | 7 | 104–110 | 12 |
| July | 36 | 57.5–115.0 | 35 | 49.5–142 | 71 |
| August | 27 | 57.0–110.0 | 26 | 65–110 | 53 |
| September | 23 | 72.0–97.0 | 27 | 70–93 | 50 |
| October | 16 | 72.0–92.0 | 9 | 77–88 | 25 |
| November | 18 | 82.0–93.0 | 11 | 73–101 | 29 |
| December | 10 | 91.0–98.0 | 8 | 84–100 | 18 |
| Total | 171 | 57.0–123.0 | 156 | 49.5–142.0 | 327 |
| L∞ | (cm) | k/kR/kG | (Year−1) | t0/b/c | - | AICc | ∆AICc | ||
|---|---|---|---|---|---|---|---|---|---|
| Female | VBGF | 203.5 | (152.3–254.7) | 0.104 | (0.057–0.151) | −3.54 | (−4.14–−2.93) | 1270.82 | 0.00 |
| VBGF with L0 | 122.3 | (117.0–127.7) | 0.420 | (0.363–0.478) | 1447.79 | 176.97 | |||
| Roberstson | 142.7 | (132.2–153.1) | 0.366 | (0.316–0.416) | 0.64 | (0.25–1.02) | 1272.10 | 1.28 | |
| Gompertz | 158.6 | (140.7–176.6) | 0.235 | (0.186–0.283) | −0.32 | (−0.73–0.08) | 1271.22 | 0.40 | |
| Male | VBGF | 170.2 | (151.4–189.0) | 0.146 | (0.110–0.182) | −3.16 | (−3.57–−2.75) | 1178.97 | 0.00 |
| VBGF with L0 | 114.0 | (110.2–117.7) | 0.595 | (0.523–0.667) | 1329.68 | 150.71 | |||
| Roberstson | 148.7 | (140.0–157.3) | 0.338 | (0.302–0.374) | 0.79 | (0.44–1.13) | 1191.25 | 12.28 | |
| Gompertz | 155.8 | (143.8–167.8) | 0.241 | (0.203–0.278) | −0.47 | (−0.73–−0.21) | 1185.18 | 6.21 |
| Area | Sex | Model | L∞ | k/kR/kG | t0/L0/c | tmax | Reference |
|---|---|---|---|---|---|---|---|
| Northern Australia | Male | VBGF | 98.4 | 1.170 | −0.6 | 5 | Davenport and Stevens (1988) |
| Female | VBGF | 123.9 | 0.340 | −1.9 | 7 | ||
| Northeastern Australia | Male | Logistic | 105.6 | 0.916 | 52.5 | 9 | Harry et al., (2013) |
| Female | VBGF | 126.6 | 0.336 | 55.3 | 14 | ||
| Taiwan Strait | Male | VBGF | 170.2 | 0.146 | −3.16 | 8 | This study |
| Female | Gompertz | 158.6 | 0.235 | −0.32 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joung, S.-J.; Hsu, Z.-Y.; Su, K.-Y.; Liu, K.-M. Age and Growth of the Spot-Tail Shark, Carcharhinus sorrah, in the Taiwan Strait. J. Mar. Sci. Eng. 2022, 10, 413. https://doi.org/10.3390/jmse10030413
Joung S-J, Hsu Z-Y, Su K-Y, Liu K-M. Age and Growth of the Spot-Tail Shark, Carcharhinus sorrah, in the Taiwan Strait. Journal of Marine Science and Engineering. 2022; 10(3):413. https://doi.org/10.3390/jmse10030413
Chicago/Turabian StyleJoung, Shoou-Jeng, Zhi-Yu Hsu, Kuan-Yu Su, and Kwang-Ming Liu. 2022. "Age and Growth of the Spot-Tail Shark, Carcharhinus sorrah, in the Taiwan Strait" Journal of Marine Science and Engineering 10, no. 3: 413. https://doi.org/10.3390/jmse10030413