
Comparative Analysis of the Fatty Acid Profiles of Antarctic Krill (Euphausia superba Dana, 1850) in the Atlantic Sector of the Southern Ocean: Certain Fatty Acids Reflect the Oceanographic and Trophic Conditions of the Habitat

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
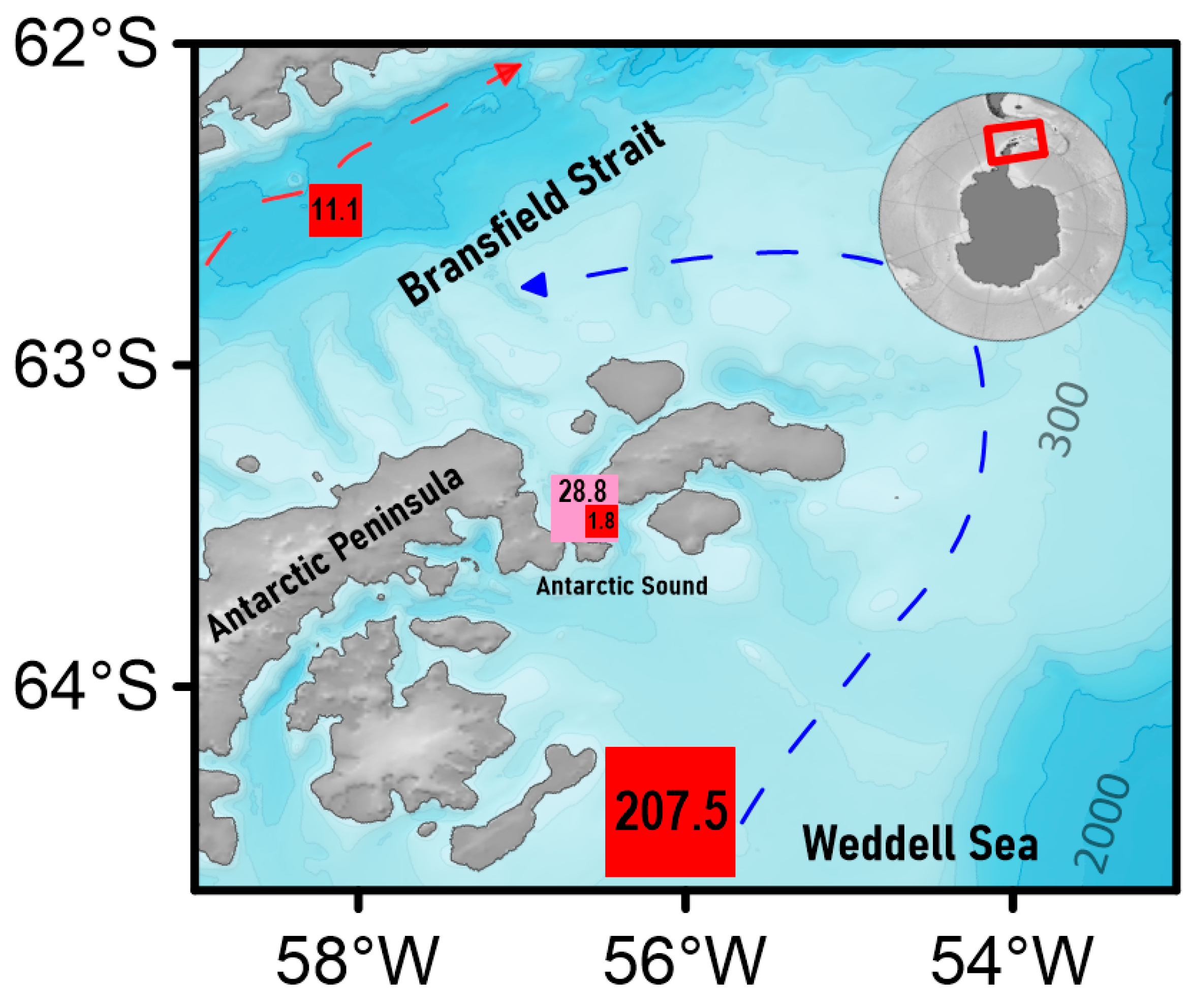
2.1. Sampling
2.2. Fatty Acid Analysis
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knox, G.A. The biology of the Southern Ocean; Cambridge University Press: New York, NY, USA, 1994; 444p. [Google Scholar]
- Mayzaud, P.; Boutoute, M.; Alonzo, F. Lipid composition of the euphausiids Euphausia vallentini and Thysanoessa macrura during summer in the Southern Indian Ocean. Antarct. Sci. 2003, 15, 463–475. [Google Scholar] [CrossRef]
- Johnston, N.M.; Murphy, E.J.; Atkinson, A.; Andrew, J.; Constable, A.J.; Cotté, C.; Cox, M.; Daly, K.L.; Driscoll, R.; Flores, H. at al. Status, Change and Futures of Zooplankton in the Southern Ocean. Front. Ecol. Evol. 2022, 9, 624692. [Google Scholar] [CrossRef]
- Wiedenmann, J.; Cresswell, K.; Mangel, M. Tenperature-dependent growth of Antarctic krill: Predictions for a changing climate from a cohort model. Mar. Ecol. Prog. Ser. 2008, 358, 191–202. [Google Scholar] [CrossRef]
- McBride, M.M.; Schram Stokke, O.; Renner, A.H.H.; Krafft, B.A.; Bergstad, O.A.; Biuw, M.; Lowther, A.D.; Stiansen, J.E. Antarctic krill Euphausia superba: Spatial distribution, abundance, and management of fisheries in a changing climate. Mar. Ecol. Prog. Ser. 2021, 668, 185–214. [Google Scholar] [CrossRef]
- Yu, G.; Piao, S.; Zhang, Y.; Liu, L.; Peng, J.; Niu, S. Moving toward a new era of ecosystem science. Geogr. Sustain. 2021, 2, 151–162. [Google Scholar] [CrossRef]
- Whitehouse, M.J.; Meredith, M.P.; Rothery, P.; Atkinson, A.; Ward, P.; Korb, R.E. Rapid warming of the ocean around South Georgia, Southern Ocean, during the 20th century: Forcing, characteristics and implications for lower trophic levels. Deep Sea Res. Part I Oceanogr. Res. Pap. 2008, 55, 1218–1228. [Google Scholar] [CrossRef]
- Schmidt, K.; Atkinson, A. Feeding and food processing in Antarctic krill (Euphausia superba Dana). In Biology ad Ecology of Antarctic Krill; Springer International Publishing: Cham, Switzerland, 2016; pp. 175–224. [Google Scholar]
- Hill, S.L.; Phillips, T.; Atkinson, A. Potential climate change effects on the habitat of Antarctic krill in the Weddel quadrant of the Sourthern Ocean. PLoS ONE 2013, 8, e72246. [Google Scholar] [CrossRef]
- Ericson, J.A.; Hellessey, N.; Nichols, P.D.; Nicol, S.; Kawaguchi, S.; Hoem, N.; Virtue, P. New insights into the seasonal diet of Antarctic krill using triacylglycerol and phospholipid fatty acids, and sterol composition. Polar Biol. 2019, 42, 1985–1996. [Google Scholar] [CrossRef]
- Yan, Y.; Lin, Y.; Zhang, L.; Gao, G.; Chen, S.; Chi, C.; Hu, S.; Sang, Y.; Chu, X.; Zhou, Q.; et al. Dietary supplementation with fermented antarctic krill shell improved the growth performance, digestive and antioxidant capability of Macrobrachium nipponense. Aquac. Rep. 2023, 30, 101587. [Google Scholar] [CrossRef]
- Hellessey, N.; Johnson, R.; Ericson, J.A.; Nichols, P.D.; Kawaguchi, S.; Nicol, S.; Hoem, N.; Virtue, P. Antarctic Krill Lipid and Fatty acid Content Variability is Associated to Satellite Derived Chlorophyll a and Sea Surface Temperatures. Sci. Rep. 2020, 10, 6060. [Google Scholar] [CrossRef]
- Gigliotti, J.C.; Davenport, M.P.; Beamer, S.K.; Tou, J.C.; Jaczynski, J. Extraction and characterization of lipids from Antarctic krill (Euphausia superba). Food Chem. 2011, 125, 1028–1036. [Google Scholar] [CrossRef]
- Sologub, D.O. Modern Features of Distribution, Biology and Horizontal Migrations of Antarctic Krill (Euphausia superba) in the Atlantic Sector of Antarctica. Ph.D. Thesis, VNIRO Publ. House, Moscow, Russia, 2016; pp. 1–247. [Google Scholar]
- Nicol, S.; Foster, J. The Fishery for Antarctic Krill: Its Current Status and Management Regime. In Biology and Ecology of Antarctic Krill; Springer: Cham, Switzerland, 2016; pp. 387–421. [Google Scholar]
- Atkinson, A.; Siegel, V.; Pakhomov, E.A.; Rothery, P.; Loeb, V.; Ross, R.M.; Quetin, L.B.; Schmidt, K.; Fretwell, P.; Murphy, E.J.; et al. Oceanic Circumpolar Habitats of Antarctic Krill. Mar. Ecol. Prog. Ser. 2008, 362, 1–23. [Google Scholar] [CrossRef]
- Siegel, V.; Watkins, J.L. Distribution, Biomass and Demography of Antarctic Krill, Euphausia superba. In Biology and Ecology of Antarctic Krill; Springer: Cham, Switzerland, 2016; pp. 21–100. [Google Scholar]
- Marchand, D.; Marty, J.-C.; Miquel, J.-C.; Rontani, J.-F. Lipids and their oxidation products as biomarkers for carbon cycling in the northwestern Meditterranean Sea: Results from a sediment trap study. Mar. Chem. 2005, 95, 129–147. [Google Scholar] [CrossRef]
- Rush, D.; Sinninghe Damsté, J.S. Lipids as paleomarkers to constrain the marine nitrogen cycle. Environ. Microbiol. 2017, 19, 2119–2132. [Google Scholar] [CrossRef] [PubMed]
- Shilla, D.A. Sources and major biogeochemical pathways of organic matter in the mangrove system of Rufiji estuary. Chem. Ecol. 2019, 35, 524–536. [Google Scholar] [CrossRef]
- Chen, X.; Dong, L.; Zhao, W.; Jian, H.; Wang, J.; Wang, F. The effects of metabolism and temperature on carbon isotope composition of lipids in marine bacterium Shewanella piezotolerans WP3. Chem. Geol. 2022, 606, 120963. [Google Scholar] [CrossRef]
- Murzina, S.A. Role of Lipids and Their Fatty Acid Components in Ecological and Biochemical Adaptations of Fish of the Northern Seas. Ph.D. Thesis, IPEE RAS, Moscow, Russia, 2019; 376p. [Google Scholar]
- Morozov, E.G.; Flint, M.V.; Orlov, A.M.; Frey, D.I.; Molodtsova, T.N.; Krechik, V.A.; Latushkin, A.A.; Salyuk, P.A.; Murzina, S.A.; Minin, K.V.; et al. Oceanographic and Ecosystem Studies in the Atlantic Sector of Antarctica (Cruise 87 of the Research Vessel Akademik Mstislav Keldysh). Oceanology 2022, 62, 721–723. [Google Scholar] [CrossRef]
- Kasyan, V.V.; Bitiutskii, D.G.; Mishin, A.V.; Zuev, O.A.; Murzina, S.A.; Sapozhnikov, P.V.; Kalinina, O.Y.; Syomin, V.L.; Kolbasova, G.D.; Voronin, V.P.; et al. Composition and Distribution of Plankton Communities in the Atlantic Sector of the Southern Ocean. Diversity 2022, 14, 923. [Google Scholar] [CrossRef]
- Bitiutskii, D.G.; Samyshev, E.Z.; Minkina, N.I.; Melnikov, V.V.; Chudinovskih, E.S.; Usachev, S.I.; Salyuk, P.A.; Serebrennikov, A.N.; Zuev, O.A.; Orlov, A.M. Distribution and demography of Antarctic krill and salps in the Atlantic sector of the Southern Ocean during austral summer 2021–2022. Water 2022, 14, 3812. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Pekkoeva, S.N.; Voronin, V.P.; Mishin, A.V.; Orlov, A.M.; Murzina, S.A. Living in the Extreme: Fatty Acid Profiles and Their Specificity in Certain Tissues of Dominant Antarctic Silverfish, Pleuragramma antarcticum, from the Antarctic Sound (Southern Ocean) Collected during the Austral Summer. Diversity 2022, 14, 817. [Google Scholar] [CrossRef]
- Murzina, S.A.; Pekkoeva, S.N.; Kondakova, E.A.; Nefedova, Z.A.; Filippova, K.A.; Nemova, N.N.; Orlov, A.M.; Berge, J.; Falk-Petersen, S. Tiny but fatty: Lipids and fatty acids in the daubed shanny (Leptoclinus maculatus), a small fish in Svalbard waters. Biomolecules 2020, 10, 368. [Google Scholar] [CrossRef]
- Petursdottir, H.; Gislason, A.; Falk-Petersen, S. Lipid classes and fatty acid composition of muscle, liver and skull oil in deep-sea redfish Sebastes mentella over the Reykjanes Ridge. J. Fish Biol. 2008, 73, 2485–2496. [Google Scholar] [CrossRef]
- Shitikov, V.K.; Mastitsky, S.E. Classification, Regression, Data Mining Algorithms Using R. 2017. Available online: https://github.com/ranalytics/data-mining (accessed on 1 June 2023).
- Wood, S.N. Generalized Additive Models: An Introduction with R; Hall/CRC and Chapman: Boca Raton, FL, USA, 2006; 410p. [Google Scholar]
- Kabakoff, R. R in Action: Data Analysis and Graphics with R; Volkova, P.A., Translator; DMK Press: Moscow, Russia, 2014; p. 588. [Google Scholar]
- Heywood, K.J.; Naveira Garabato, A.C.; Stevens, D.P.; Muench, R.D. On the fate of the Antarctic Slope Front and the origin of the Weddell Front. J. Geophys. Res. 2004, 109, C06021. [Google Scholar] [CrossRef]
- Thompson, A.F.; Stewart, A.L.; Spence, P.; Heywood, K.J. The Antarctic Slope Current in a Changing Climate. Rev. Geophys. 2018, 56, 741–770. [Google Scholar] [CrossRef]
- Frey, D.I.; Krechik, V.A.; Morozov, E.G.; Drozd, I.D.; Gordey, A.S.; Latushkin, A.A.; Mekhova, O.S.; Mukhametianov, R.Z.; Murzina, S.A.; Ostroumova, S.A.; et al. Water Exchange between Deep Basins of the Bransfield Strait. Water 2022, 14, 3193. [Google Scholar] [CrossRef]
- Voronin, V.P.; Nemova, N.N.; Ruokolainen, T.R.; Artemenkov, D.V.; Rolskii, A.Y.; Orlov, A.M.; Murzina, S.A. Into the Deep: New Data on the Lipid and Fatty Acid Profile of Redfish Sebastes mentella Inhabiting Different Depths in the Irminger Sea. Biomolecules 2021, 11, 704. [Google Scholar] [CrossRef] [PubMed]
- Tokarczyk, R. Classification of water masses in the Bransfield Strait and Southern part of the Drake Passage using a method of statistical multidimensional analysis. Pol. Polar Res. 1987, 8, 333–336. [Google Scholar]
- Frey, D.I.; Krechik, V.A.; Gordey, A.S.; Gladyshev, S.V.; Churin, D.A.; Drozd, I.D.; Osadchiev, A.A.; Kashin, S.V.; Morozov, E.G.; Smirnova, D.A. Austral summer circulation in the Bransfield Strait based on SADCP measurements and satellite altimetry. Front. Mar. Sci. 2023, 10, 1111541. [Google Scholar] [CrossRef]
- Tarling, G.A.; Shreeve, R.S.; Hirst, A.G.; Atkinson, A.; Pond, D.W.; Murphy, E.J. Natural growth rates in Antarctic krill (Euphausia superba): I. Improving methodology and predicting intermolt period. Limnol. Oceanogr. 2006, 51, 959–972. [Google Scholar] [CrossRef]
- Reiss, C.; Cossio, A.; Santora, J.A.; Dietrich, K.S.; Murray, A.; Greg Mitchell, B.; Walsh, J.; Weiss, E.L.; Gimpel, C.; Jones, C.D.; et al. Overwinter habitat selection by Antarctic krill under varying sea-ice conditions: Implications for top predators and fishery management. Mar. Ecol. Prog. Ser. 2017, 568, 1–16. [Google Scholar] [CrossRef]
- Krek, A.V.; Krek, E.V.; Krechik, V.A. The Circulation and Mixing Zone in the Antarctic Sound in February 2020. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 83–100. [Google Scholar] [CrossRef]
- Khimchenko, E.; Frey, D.; Morozov, E.G. Tidal internal waves in the Bransfield Strait, Antarctica. Russ. J. Earth Sci. 2020, 20, ES2006. [Google Scholar] [CrossRef]
- Thompson, A.F.; Heywood, K.J. Frontal structure and transport in the northwestern Weddell Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2008, 55, 1229–1251. [Google Scholar] [CrossRef]
- Izhitskiy, A.; Romanova, N.; Vorobieva, O.; Frey, D. Impact of Ice Melting on Oceanographic and Hydrobiological Characteristics of Surface Waters in the Powell Basin, Weddell Sea, in January–February 2020. Oceanology 2022, 62, 439–446. [Google Scholar] [CrossRef]
- Morozov, E.G.; Krechik, V.; Frey, D.; Zamshin, V. Currents in the Western Part of the Weddell Sea and Drift of large Iceberg A68A. Oceanology 2021, 61, 589–601. [Google Scholar] [CrossRef]
- Ozernyuk, N.D. Ontogenetic temperature adaptations of enzymes of poikilothermic animals. Uspechi Sovrem. Biol. 2004, 124, 534–541. [Google Scholar]
- Ross, R.M.; Quetin, L.B.; Baker, K.S.; Vernet, M.; Smith, R.S. Growth limitation in young Euphausia superba under field conditions. Limnol. Oceanogr. 2000, 45, 31–43. [Google Scholar] [CrossRef]
- Stübing, D.; Hagen, W.; Schmidt, K. On the use of lipid biomarkers in marine food web analyses: An experimental case study on the Antarctic krill, Euphausia superba. Limnol. Oceanogr. 2003, 48, 1685–1700. [Google Scholar] [CrossRef]
- De Felice, A.; Manini, E.; Biagiotti, I.; Leonori, I. Bioenergetics of Euphausia superba and Euphausia crystallorophias in the Ross Sea. Diversity 2023, 15, 480. [Google Scholar] [CrossRef]
- Smyntek, P.M.; Teece, M.A.; Schultz, K.L.; Storch, A.J. Taxonomic differences in the essential fatty acid composition groups of freshwater zooplankton relate to reproductive demands and generation time. Freshw. Biol. 2008, 53, 1768–1782. [Google Scholar] [CrossRef]
- Sargent, J.R.; Henderson, J.R.; Tocher, D.R. The Lipids. In Fish Nutrition; Halver, J.E., Ed.; Academic Press: New York, NY, USA, 1989; pp. 154–218. [Google Scholar]
- Lee, R.; Hagen, W.; Kattner, G. Lipid storage in marine zooplankton. Mar. Ecol. Prog. Ser. 2006, 307, 273–306. [Google Scholar] [CrossRef]
- Kattner, G.; Hagen, W. Lipids in marine copepods: Latitudinal characteristics and perspective to global warming. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M., Kainz, M.J., Eds.; Springer: New York, NY, USA, 2009; pp. 257–280. [Google Scholar]
- Schmidt, K.; Atkinson, A.; Petzke, K.J.; Voss, M.; Pond, D.W. Protozoans as a food source for Antarctic krill, Euphausia superba: Complementary insights from stomach content, fatty acids, and stable isotopes. Limnol. Oceanogr. 2006, 51, 2409–2427. [Google Scholar] [CrossRef]
- Dalsgaard, J.; John, M.S.; Kattner, G.; Müller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar] [CrossRef] [PubMed]
- Falk-Petersen, S.; Sargent, J.R.; Lønne, O.J.; Timofeev, S. Functional biodiversity of lipids in Antarctic zooplankton: Calanoides acutus, Calanus propinquus, Thysanoessa macrura and Euphausia crystallorophias. Polar Biol. 1999, 21, 37–47. [Google Scholar] [CrossRef]
- Graeve, M.; Hagen, W.; Kattner, G. Herbivorous or omnivorous? On the significance of lipid compositions as trophic markers in Antarctic copepods. Deep. Sea Res. Part I Oceanogr. Res. Pap. 1994, 41, 915–924. [Google Scholar] [CrossRef]
- Murzina, S.A.; Voronin, V.P.; Bitiutskii, D.G.; Orlov, A.M. Modeling of the Distribution and Relationship with Abiotic Environmental Factors of Juvenile and Adult Antarctic Krill Euphausia superba Based on Actual Hydrophysical Measurements. Oceanology 2023, 63, 568–573. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Developmental Stage | Juvenile | Mature | ||||
|---|---|---|---|---|---|---|
| Area | AP | AS | BS | AP | AS | BS |
| TL | 24.47 ± 1.17 | 13.85 ± 1.68 a | 9.4 ± 0.99 a | 21.3 ± 0.66 * | 14.43 ± 0.77 a | 7.51 ± 0.32 a,b |
| 12:0 | 0.12 ± 0 | 0.09 ± 0.01 a | 0.08 ± 0.01 a | 0.12 ± 0 | 0.07 ± 0 a | 0.04 ± 0.01 a,b |
| 14:0 | 9.84 ± 0.23 | 6.91 ± 0.36 a | 8.75 ± 0.35 a,b | 9.47 ± 0.14 | 7.56 ± 0.24 a | 4.14 ± 0.24 a,b,* |
| 15:0 | 0.18 ± 0.01 | 0.18 ± 0.01 | 0.32 ± 0.01 a,b | 0.18 ± 0.01 * | 0.16 ± 0 | 0.26 ± 0.01 a,b |
| 16:0 | 21.51 ± 0.21 | 21.34 ± 0.45 | 22.57 ± 0.18 a,b | 21.65 ± 0.1 | 22.26 ± 0.2 a | 20.3 ± 0.21 a,b,* |
| 18:0 | 3.22 ± 0.09 | 3.35 ± 0.1 | 2.22 ± 0.15 a,b | 3.32 ± 0.13 | 2.42 ± 0.16 a,* | 2.74 ± 0.17 a,b |
| 19:0 | 0.04 ± 0.01 | 0.04 ± 0.01 | 0.1 ± 0.02 a,b | 0.04 ± 0 | 0.04 ± 0.01 a | 0.15 ± 0.02 a,b |
| 20:0 | 6.68 ± 0.22 | 7.83 ± 0.75 | 1.68 ± 0.26 a,b | 6.86 ± 0.18 | 3.09 ± 0.6 a,* | 2.27 ± 0.15 a,b |
| 23:0 | 0.05 ± 0 | 0.15 ± 0.04 a | 0.98 ± 0.19 a,b | 0.05 ± 0 | 0.24 ± 0.04 a | 1.59 ± 0.23 a,b |
| 24:0 | 1.25 ± 0.04 | 1.46 ± 0.14 | 0.32 ± 0.04 a,b | 1.29 ± 0.03 | 0.57 ± 0.11 a,* | 0.45 ± 0.03 a,b |
| cis16:1(n-7) | 5.95 ± 0.17 | 6.73 ± 0.44 | 5.21 ± 0.29 a | 6.31 ± 0.1 * | 6.43 ± 0.16 | 3.59 ± 0.32 a,b,* |
| cis17:1(n-7) | 0.7 ± 0.03 | 1.06 ± 0.05 a | 0.73 ± 0.02 b | 0.7 ± 0.02 | 0.74 ± 0.05 a,* | 0.27 ± 0.05 a,b,* |
| cis18:1(n-9) | 13.35 ± 0.16 | 10.92 ± 0.72 a | 10.2 ± 0.75 a | 13.34 ± 0.14 | 12.51 ± 0.24 a | 9.24 ± 0.39 a,b,* |
| cis18:1(n-7) | 4.53 ± 0.04 | 4.23 ± 0.32 | 5.91 ± 0.13 a,b | 4.73 ± 0.04 * | 5.82 ± 0.14 a,* | 7.69 ± 0.19 a,b,* |
| cis20:1(n-9) | 0.98 ± 0.03 | 0.66 ± 0.06 a | 0.61 ± 0.07 a | 1 ± 0.02 | 0.89 ± 0.04 a | 0.56 ± 0.04 a,b |
| cis18:2(n-6) | 1.39 ± 0.05 | 0.81 ± 0.1 a | 1.83 ± 0.05 a,b | 1.24 ± 0.04 * | 0.95 ± 0.03 a | 1.62 ± 0.05 a,b |
| cis18:3(n-6) | 0.08 ± 0 | 0.06 ± 0.01 a | 0.16 ± 0.01 a,b | 0.08 ± 0 | 0.08 ± 0 | 0.07 ± 0.01 * |
| cis20:2(n-6) | 0.05 ± 0 | 0.05 ± 0.01 | 0.18 ± 0.05 a,b | 0.05 ± 0 | 0.11 ± 0.01 a | 0.28 ± 0.06 a,b |
| cis20:4(n-6) | 0.02 ± 0 | 0.04 ± 0.01 a | 0.09 ± 0.02 a,b | 0.02 ± 0 | 0.04 ± 0 a | 0.13 ± 0.01 a,b |
| cis18:3(n-3) | 0.72 ± 0.02 | 0.41 ± 0.05 a | 1.19 ± 0.06 a,b | 0.66 ± 0.02 | 0.53 ± 0.02 a | 1 ± 0.06 a,b |
| cis18:4(n-3) | 4.64 ± 0.28 | 2.34 ± 0.4 a | 5.23 ± 0.14 b | 3.77 ± 0.13 * | 2.6 ± 0.12 a | 2 ± 0.15 a,b,* |
| cis20:5(n-3) | 15.41 ± 0.31 | 22.61 ± 0.58 a | 18.35 ± 0.57 a,b | 15.97 ± 0.22 | 22.22 ± 0.34 a | 22.54 ± 0.46 a,* |
| cis22:5(n-3) | 0.29 ± 0.01 | 0.42 ± 0.04 a | 0.44 ± 0.04 a | 0.29 ± 0.01 | 0.36 ± 0.01 a | 0.62 ± 0.02 a,b,* |
| cis22:6(n-3) | 8.31 ± 0.2 | 7.66 ± 0.76 | 12.11 ± 0.81 a,b | 8.17 ± 0.18 | 9.75 ± 0.33 a | 17.79 ± 0.58 a,b,* |
| SFA | 43.34 ± 0.23 | 41.8 ± 0.58 a | 37.49 ± 0.64 a,b | 43.4 ± 0.22 | 36.72 ± 0.72 a,* | 32.4 ± 0.54 a,b,* |
| MUFA | 25.74 ± 0.32 | 23.8 ± 1.04 | 22.88 ± 1.04 a | 26.33 ± 0.23 * | 26.6 ± 0.47 | 21.52 ± 0.81 a,b |
| (n-3) PUFA | 29.38 ± 0.37 | 33.44 ± 1.44 a | 37.37 ± 1.42 a | 28.87 ± 0.31 | 35.5 ± 0.63 a | 43.96 ± 0.94 a,b,* |
| (n-6) PUFA | 1.54 ± 0.05 | 0.96 ± 0.12 a | 2.27 ± 0.02 a,b | 1.39 ± 0.04 * | 1.18 ± 0.04 a | 2.11 ± 0.08 a,b |
| PUFA | 30.92 ± 0.4 | 34.4 ± 1.56 | 39.63 ± 1.43 a | 30.26 ± 0.33 | 36.68 ± 0.66 a | 46.07 ± 0.95 a,b,* |
| 18:1(n-9)/18:1(n-7) | 3.07 | 2.58 a | 1.73 a,b | 2.82 | 2.15 | 1.20 |
| 20:5(n-3)/22:6(n-3) | 1.85 | 2.95 a | 1.51 a,b | 1.95 | 2.28 a | 1.27 a,b |
| 16:1(n-7)/18:4(n-3) | 1.28 | 2.87 a | 0.99 a,b | 1.70 | 2.47 a | 1.8 a,b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murzina, S.A.; Voronin, V.P.; Bitiutskii, D.G.; Mishin, A.V.; Khurtina, S.N.; Frey, D.I.; Orlov, A.M. Comparative Analysis of the Fatty Acid Profiles of Antarctic Krill (Euphausia superba Dana, 1850) in the Atlantic Sector of the Southern Ocean: Certain Fatty Acids Reflect the Oceanographic and Trophic Conditions of the Habitat. J. Mar. Sci. Eng. 2023, 11, 1912. https://doi.org/10.3390/jmse11101912
Murzina SA, Voronin VP, Bitiutskii DG, Mishin AV, Khurtina SN, Frey DI, Orlov AM. Comparative Analysis of the Fatty Acid Profiles of Antarctic Krill (Euphausia superba Dana, 1850) in the Atlantic Sector of the Southern Ocean: Certain Fatty Acids Reflect the Oceanographic and Trophic Conditions of the Habitat. Journal of Marine Science and Engineering. 2023; 11(10):1912. https://doi.org/10.3390/jmse11101912
Chicago/Turabian StyleMurzina, Svetlana A., Viktor P. Voronin, Dmitry G. Bitiutskii, Aleksej V. Mishin, Svetlana N. Khurtina, Dmitry I. Frey, and Alexei M. Orlov. 2023. "Comparative Analysis of the Fatty Acid Profiles of Antarctic Krill (Euphausia superba Dana, 1850) in the Atlantic Sector of the Southern Ocean: Certain Fatty Acids Reflect the Oceanographic and Trophic Conditions of the Habitat" Journal of Marine Science and Engineering 11, no. 10: 1912. https://doi.org/10.3390/jmse11101912
APA StyleMurzina, S. A., Voronin, V. P., Bitiutskii, D. G., Mishin, A. V., Khurtina, S. N., Frey, D. I., & Orlov, A. M. (2023). Comparative Analysis of the Fatty Acid Profiles of Antarctic Krill (Euphausia superba Dana, 1850) in the Atlantic Sector of the Southern Ocean: Certain Fatty Acids Reflect the Oceanographic and Trophic Conditions of the Habitat. Journal of Marine Science and Engineering, 11(10), 1912. https://doi.org/10.3390/jmse11101912