Abstract
Seagrass ecosystems have been declining, and restorations are conducted in many parts of the world to compensate for habitat loss and restore the ecosystem services seagrasses provide. Assessment of transplantation success requires the monitoring of the level of biodiversity between the donor and transplanted sites. In this study, we assessed a seagrass ecosystem after restoration in terms of the diversity of marine organisms using environmental DNA (eDNA) to compare four sites: (1) bare sand, (2) a natural meadow of Cymodocea serrulata, (3) a natural meadow of Halophila ovalis, and (4) a transplanted seagrass meadow. The results showed the presence of 3 domains, 34 phyla, 59 classes, 92 orders, 155 families, 156 genera, and 121 species. Proteobacteria, Actinobacteria, Cyanobacteria, and Bacteroidetes were the dominant bacterial phyla. Among eukaryotes, Phragmoplastophyta/Charophyta (epiphytes), Ascomycota (fungi), Cnidaria (jelly fish), and Arthropoda (Crabs and bivalves) were the dominant phyla. Dugong tails and commercial species (sea cucumber, dog conch, and swimming crab) have been observed in both the natural and transplanted meadows. Relative abundance among the four sites was significantly different. There were no differences in species richness and evenness between the four sites and no differences in species richness and evenness between the natural meadows and the transplanted seagrass meadow. It is possible that transplanted seagrass meadow can be successfully restored and established and can provide habitat for fauna and microbes. Additionally, fauna are not limited in their capacity to move between the natural and transplanted habitats. This study provides an assessment of biodiversity of restored seagrass patches and a better understanding of a seagrass ecosystem after restoration. However, to assess seagrass ecosystem services after restoration and the success of restoration actions, long-term monitoring of marine organism diversity and additional assessments are needed.
1. Introduction
Seagrasses provide ecosystem functions and processes such as food provision, pathogen trapping, shoreline protection, climate regulation, carbon sequestration, sediment trapping, global biogeochemical cycling, and nursery grounds, as well as shelters to diverse communities of marine organisms [1,2,3]. However, the worldwide area of seagrass meadows has shrunk by around 29% compared with the area first recorded in 1879 [4], and some research has predicted that seagrass coverage will decrease by around 30–40% in the next 100 years [5]. Destructive fishing practices, sediment loading, boating, shipping, and coastal development are the main anthropogenic activities driving seagrass meadow degradation [6,7]. Climate change also plays a role in seagrass degradation. Higher temperatures and carbon dioxide levels were reported to reduce growth rates and increase the mortality of seagrass [8,9]. The reduced area of seagrass meadows has changed the diversity and abundance of the seagrass benthic community because of habitat loss [10]. Fauna growth rates change [11], epifauna community structures shift [10], and in turn, the benthic bacterial community adapts [12].
A number of experimental restorations have been implemented to regenerate declining seagrass meadows [13,14]. Successful restorations have been demonstrated by studies of Zostera marina in Virginia, USA [15] and of Z. muelleri at Whangarei, New Zealand [16]. Restored seagrass beds may attract mobile macrofauna and provide refuge and food for other organisms [17,18]. As time passes, newly restored seagrass beds are expected to improve ecosystem functions and services such as seawater quality, carbon sequestration, nursery habitats, and coastal protection [19,20,21]. However, some studies have demonstrated that success in restoring seagrass meadows can be patchy and that few have been successful in the long term [22].
The restoration and rehabilitation of seagrass meadows is influenced by biotic and abiotic factors such as predation, sediment resuspension, and sediment type [23,24]. Seagrass transplantation has been the most widely employed restoration technique but may be impacted by the choice of donor site, transplant site, seagrass species, and transplantation methods. The level of biodiversity may also play an important role in seagrass restoration. Therefore, the success of seagrass transplantation requires an accurate assessment that includes a comparison of diversity between the transplant area and the donor site. Environmental DNA or eDNA has been used for detecting the DNA of organisms and assessing biodiversity in seagrass ecosystems [25]. This approach can rapidly and efficiently evaluate and monitor biodiversity. However, small segments and degradation of eDNA limits the ability to detect species composition. eDNA in combination with traditional monitoring methods such as field survey and underwater video have the potential to assess and monitor the ecosystem services of restored seagrass meadows.
In Thailand, seagrass loss is around 20–30%, and it is driven by human activities such as coastal development and sediment runoff [26,27]. To restore or rehabilitate seagrass ecosystem and services, seagrass restoration is recommended. Restoration has become a strategy to repopulate degraded seagrass meadows. However, there was no assessment of biodiversity. Then, in this observational study, we assess the biodiversity of a restored seagrass meadow and compare this with the biodiversity of donor seagrass meadows using environmental DNA (eDNA) and field surveys.
2. Materials and Methods
2.1. Study Site and Sample Collections
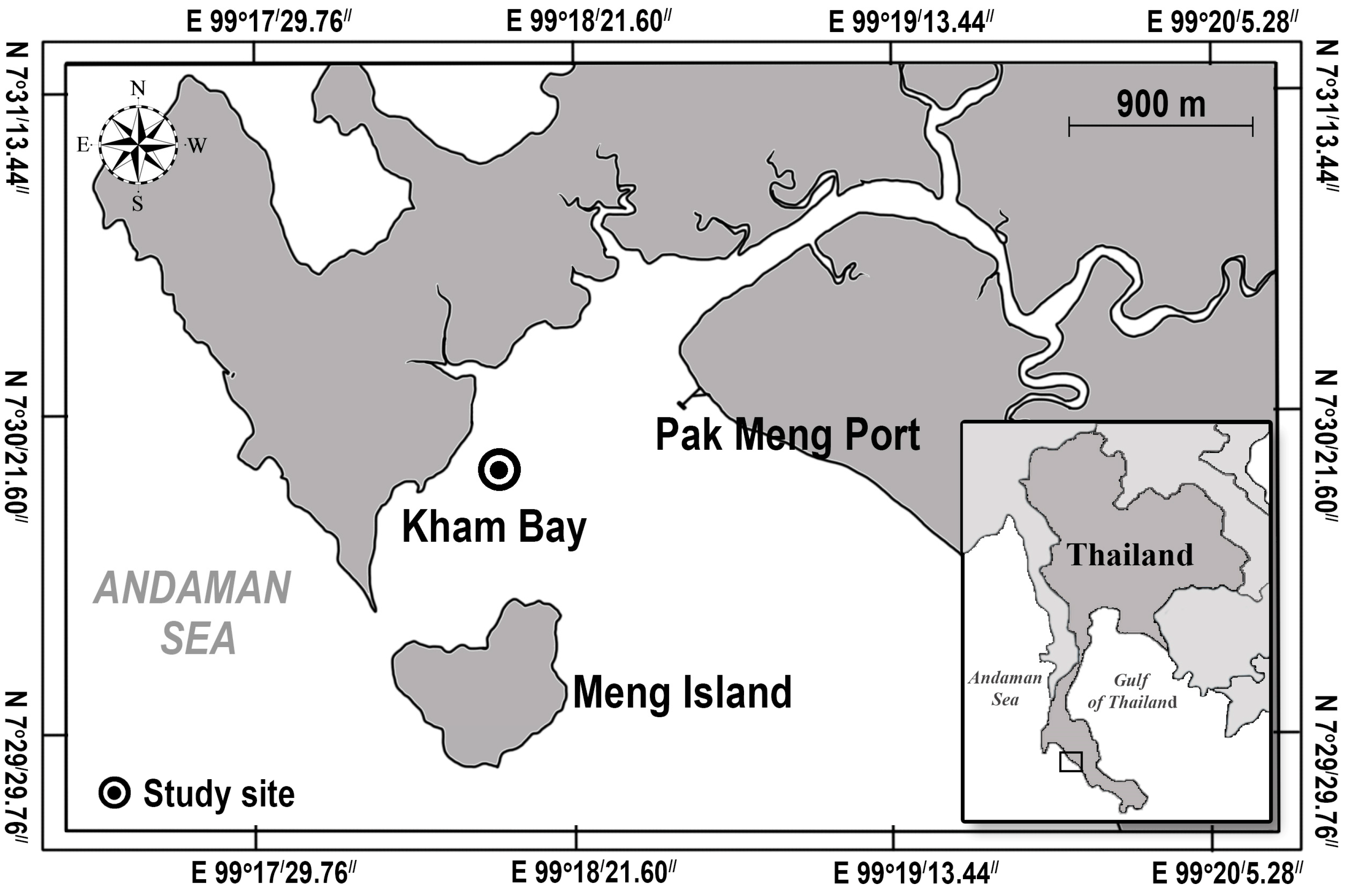
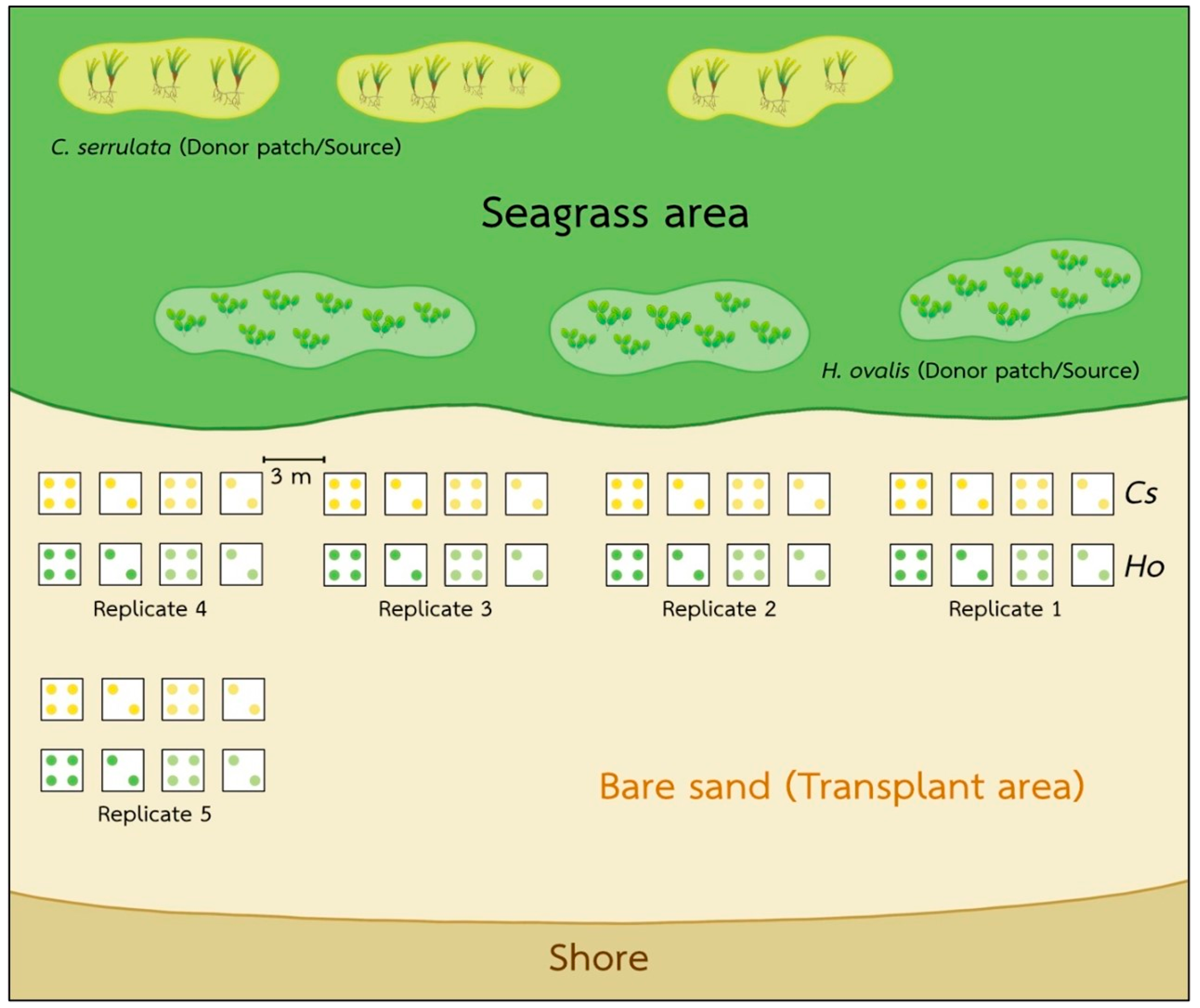
We conducted our investigation on the intertidal seagrass meadow at Kham Bay (7°30.414′ N, 99°18.406′ E), Trang Province, in the Andaman Sea, Thailand (Figure 1). In this area, two main seasons are monsoon dominated. The rainy season from May to October is dominated by the southwest monsoon, and the dry season from November to April is dominated by the northeast monsoon. This study site has been dominated by two seagrass species: Halophila ovalis and Cymodocea serrulata. However, the coverage of seagrasses in Trang province and Libong Island, which are the largest seagrass meadows in Thailand, have declined by 30% [26]. Coastal development and sedimentation from human activities were suggested as the main threats to seagrass meadows [27]. Then, the restoration and management activities in Trang province have been undertaken to enhance seagrass recovery and promote ecosystem services and biodiversity [28]. At Kham Bay, restoration was conducted in March 2021 by the Seaweed and Seagrass Research Unit (SSRU) team, Prince of Songkla University (PSU), to enhance the recovery of seagrass and compensate for seagrass habitat loss. This transplantation was attempted by using the plug method (15 cm PVC core), which is suitable in soft substrates with small and thin-leaved seagrass species. A 15 cm PVC core was used to extract the plants with the sediment, roots, and rhizomes intact from the donor site, and then, the shoots were extruded from the plug and buried in the sediment at the transplanted site. For the transplant meadow, two densities of each seagrass species, H. ovalis and C. serrulata in the planting unit (20% and 60%, respectively) and two conditions of planting unit (PU) densities/plot (2PU and 4PU) were deployed with five replicates of each condition. So, the number of PVCs implanted was 24 per replicate. Then, with five replicates, there were 120 PVCs in total. The initial transplanted area of each species was around 1.06 m2. The transplanted sites of these two species were close to each other, around 3–5 m apart, and were around 50–100 m away from the donor sites (Figure 2). All transplantation experiments were carried out on the intertidal flats with the same topography and had similar environmental conditions, such as water depth, wave current, sediment type, and light intensity. In this study site, light intensity was around 304–1090 μmol photons m−2 s−1, and the average temperature was 26–30 °C.
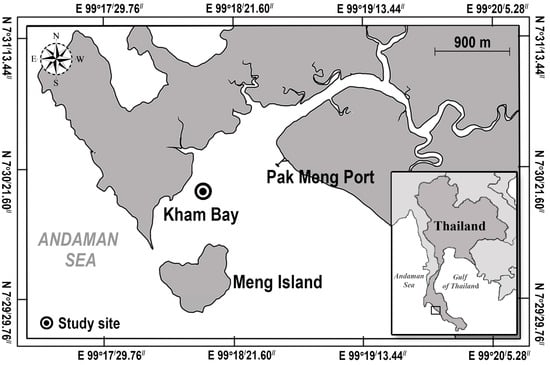
Figure 1.
The transplantation site (the blacked circle) located in the western part of Trang province, Thailand.
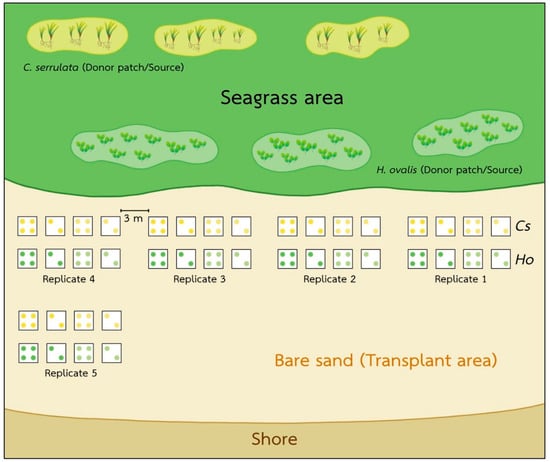
Figure 2.
The locality showing the patches of bare sand, the two different seagrass meadows, and the transplanted meadow. Each square was 1 m2, and there were 120 PCV cores in total.
To monitor the success of this transplantation, the seagrass ecosystem service in terms of diversity of marine organisms was measured using environmental DNA (eDNA) and a field survey. Degradation of eDNA and small segments of genetic material remaining in the environment may limit the biodiversity assessment. Then, the combination of eDNA and conventional methods such as a field survey have the potential to monitor and assess biodiversity. In this study, we assessed the biodiversity using eDNA and compared this with a transplanted meadow, donor seagrass meadows, and a bare sand seabed (Figure 2). H. ovalis and C. serrulata samples and soil were collected from four different sites: (1) a bare sand seabed, (2) a natural meadow of C. serrulata, (3) a natural meadow of H. ovalis, and (4) a transplanted meadow.
For seagrass samples, around 50 g of each species (3 replicates) were collected and kept in sterile microcentrifuge tubes. At each different site, three soil samples were collected from the center of the site and stored in 50 mL sterile centrifuge tubes. Then, all seagrass samples and soil were transported to the laboratory at the Division of Biological Science, Faculty of Science, Prince of Songkla University for DNA extraction. All samples were randomly collected from each condition every month from August to November 2021 to monitor the diversity and abundance of organisms. The environmental DNA methodology (eDNA) was used [29].
2.2. DNA Extraction and Sequencing
For 16S and 18S rRNA sequencing, genomics DNA was extracted from the soil using a DNeasy Power Soil Kit (Qiagen, Hilden, Germany), following the manufacturer’s instructions. Seagrass DNA was extracted using a DNeasy Plant Mini Kit (Qiagen, USA), following the manufacturer’s protocol. DNA products were preserved at −20 °C before PCR amplification. The V7 region of 18S rRNA was amplified. The forward and reverse primers (Euka02 from Guardiola et al. [30]) were F-TTTGTCTGSTTAATTSCG and R-CACAGACCTGTTATTGC, respectively [31]. We amplified an 18S rRNA sequence of vertebrate mitochondrial 12S rRNA using the primers F-GTCGGTAAAACTCGTGCCAGC and R-CATAGTGGGGTATCTAATCCCAGTTTG [32]. For the 16S rRNA, we amplified the V3-V4 variable region of the 16S rRNA bacteria and archaea genes. The sequences of forward and reverse primers were 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG-3′ and 5′ GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG-3′, respectively. The amplification protocol consisted of an initial denaturing step at 95 °C for 5 min, followed by 10 cycles of 94 °C for 30 s, 57 °C for 45 s, 72 °C for 1 min, and subsequently, 15 cycles of 94 °C for 30 s, 47 °C for 45 s, and 72 °C for 1 min, concluding with an extension step at 72 °C for 10 min. The size of the library was verified through analysis on an Agilent Technologies 2100 Bioanalyzer using a DNA 1000 chip (Agilent, Santa Clara, CA, USA). Library quantification was performed using qPCR, following the procedures outlined in the Illumina qPCR quantification protocol guide. The Illumina platform (specifically, the Illumina MiSeq, AZENTA, Suzhou, China) was employed for a paired-end read configuration.
2.3. Data Processing and Statistics
All sequencing of each 16S rRNA and 18S rRNA from three replicates of each seagrass and soil sample are pooled and used as a representative sequencing dataset for each sampling site, (1) a bare sand seabed, (2) a natural meadow of C. serrulata, (3) a natural meadow of H. ovalis, and (4) a transplanted meadow, and then the biodiversity between sampling sites was compared. The Quantitative Insights Into Microbial Ecology 2 (QIIME2) pipeline v2021.4 was used to process the specimen sequencing [33]. Quality filtering and denoising of reads were undertaken in DADA2 [34] to construct a feature table and a dataset of representative sequences. Taxonomic assignment was classified at 95% similarity using a Naive Bayes classifier trained on the SILVA database [35]. Faith’s phylogenetic diversity [36] and Pielou’s evenness index [37] were used to calculate species richness and species evenness, respectively. Alpha diversity (Chao-1, ACE, and Simpson indices) were calculated. Species richness and species evenness among sites were analyzed and compared using a pairwise Kruskal–Wallis test. Species diversity was calculated using Shannon–Wiener’s diversity index. Community dissimilarity was visualized on principal coordinates analysis (PCoA) using a weighted UniFrac distance matrix. Non-metric multidimensional scaling (nMDS) was calculated with the Bray–Curtis similarity matrix of relative abundance of the phylum level of 16S rRNA and 18S rRNA and visualized using Past software, version 4.03. To quantify and test for the differences revealed in the 16S and 18S rRNA communities among samples, we computed with permutational multivariate analysis of variance (PERMANOVA) [38].
3. Results
In this study, for the sequencing dataset of 16S rRNA, we extracted samples from the four sites and four sampling times: (1) a bare sand seabed, (2) a natural meadow of C. serrulata, (3) a natural meadow of H. ovalis, and (4) a transplanted meadow of these two species. After demultiplexing, sequencing returned a total of 1,980,636 reads. Sequencing depth ranged from a low of 129,246 to a high of 218,946 reads per sample. The mean and median depths were 153,965 and 140,176 reads per sample, respectively. Taxonomic classification by QIIME identified 2 domains, 23 phyla, 41 classes, 69 orders, 89 families, 124 genera, and 97 species.
For the sequencing dataset of 18S rRNA, samples from the four sites and four sampling times were extracted. After demultiplexing, sequencing yielded a total of 14,298,451 reads. Sequencing depth ranged from a low of 100,360 to a high of 429,968 reads per sample. The mean and median depths were 201,386 and 213,892 reads per sample, respectively. In total, 1 domain, 12 phyla, 18 classes, 23 orders, 66 families, 32 genera, and 24 species were identified by QIIME.
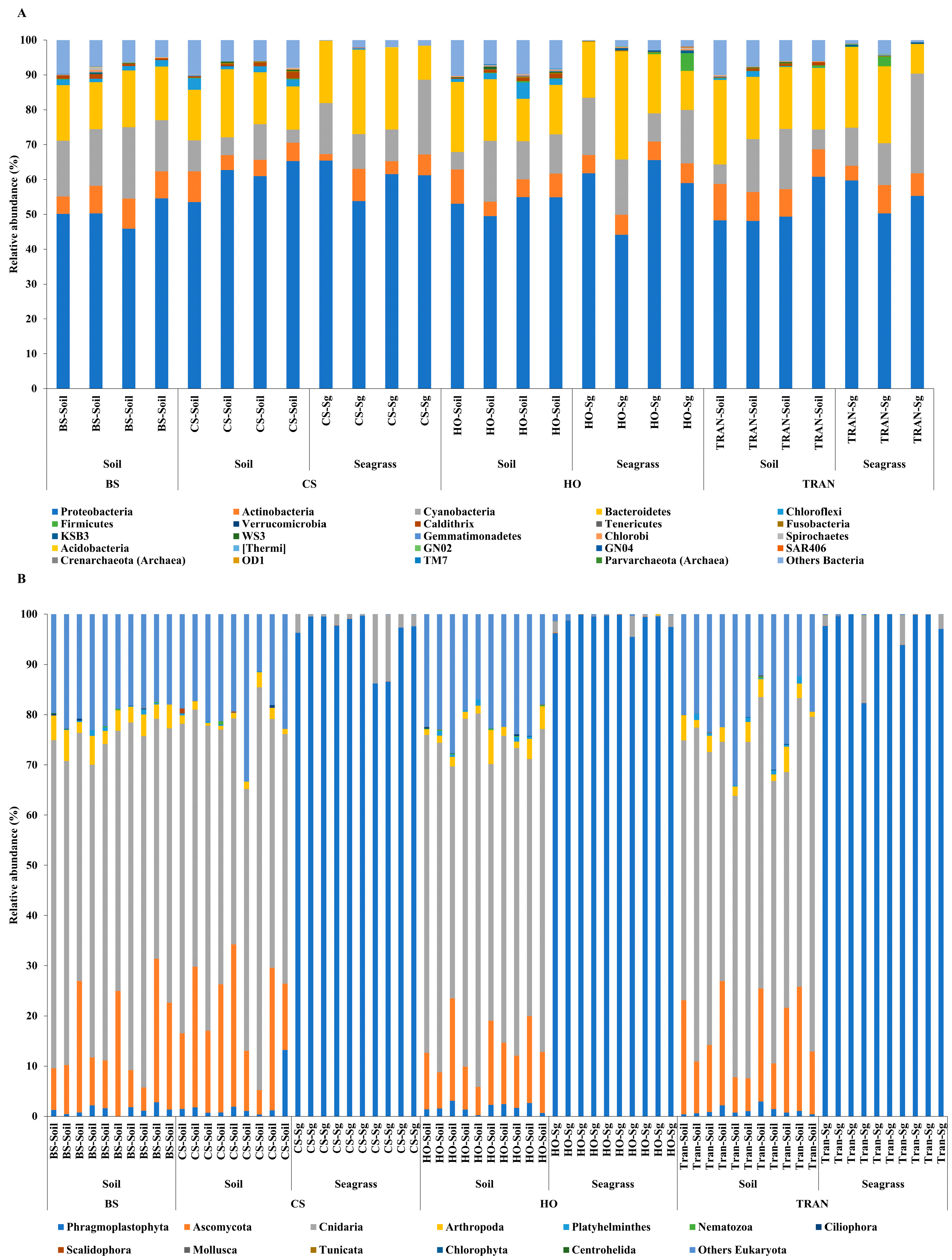
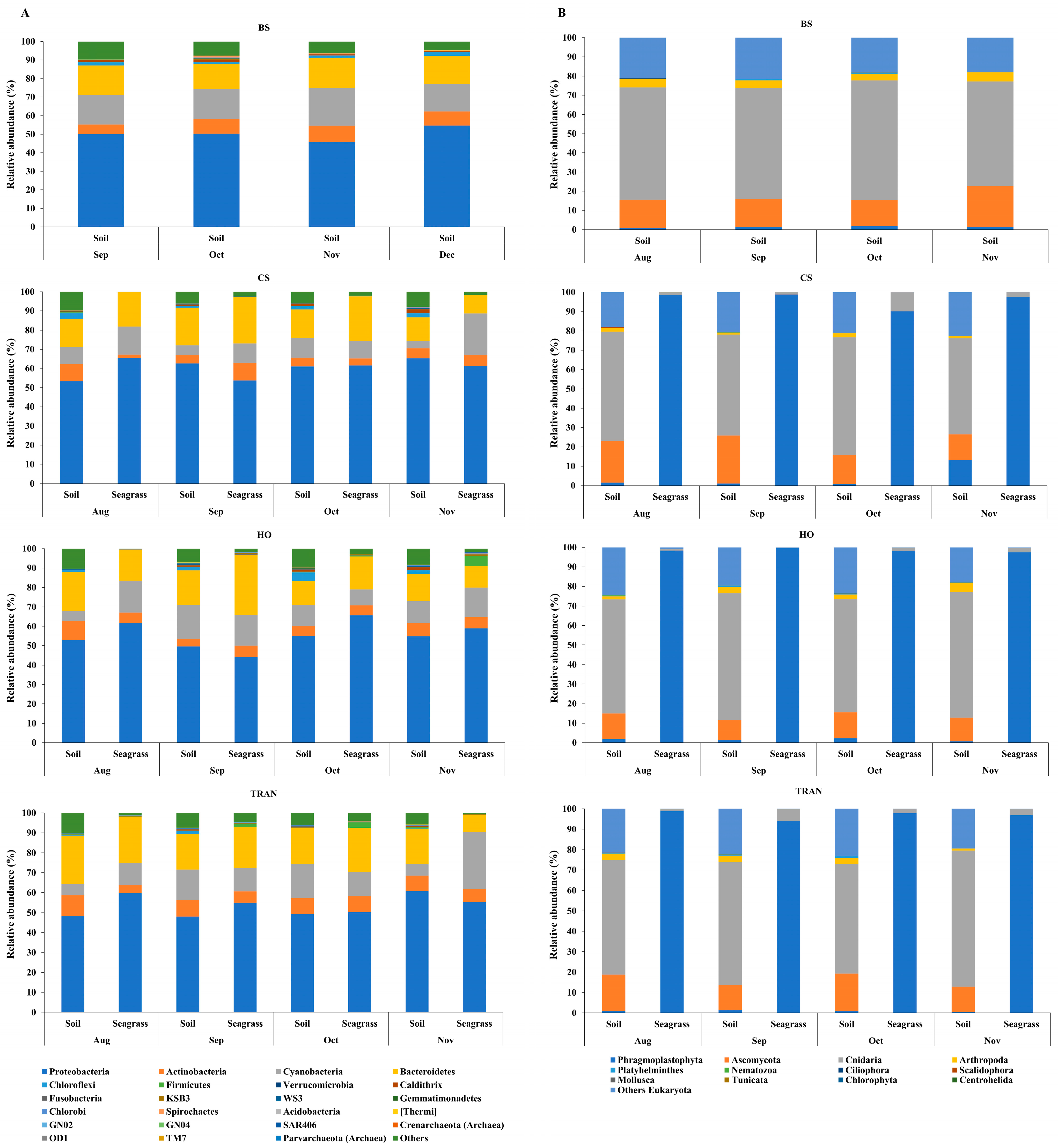
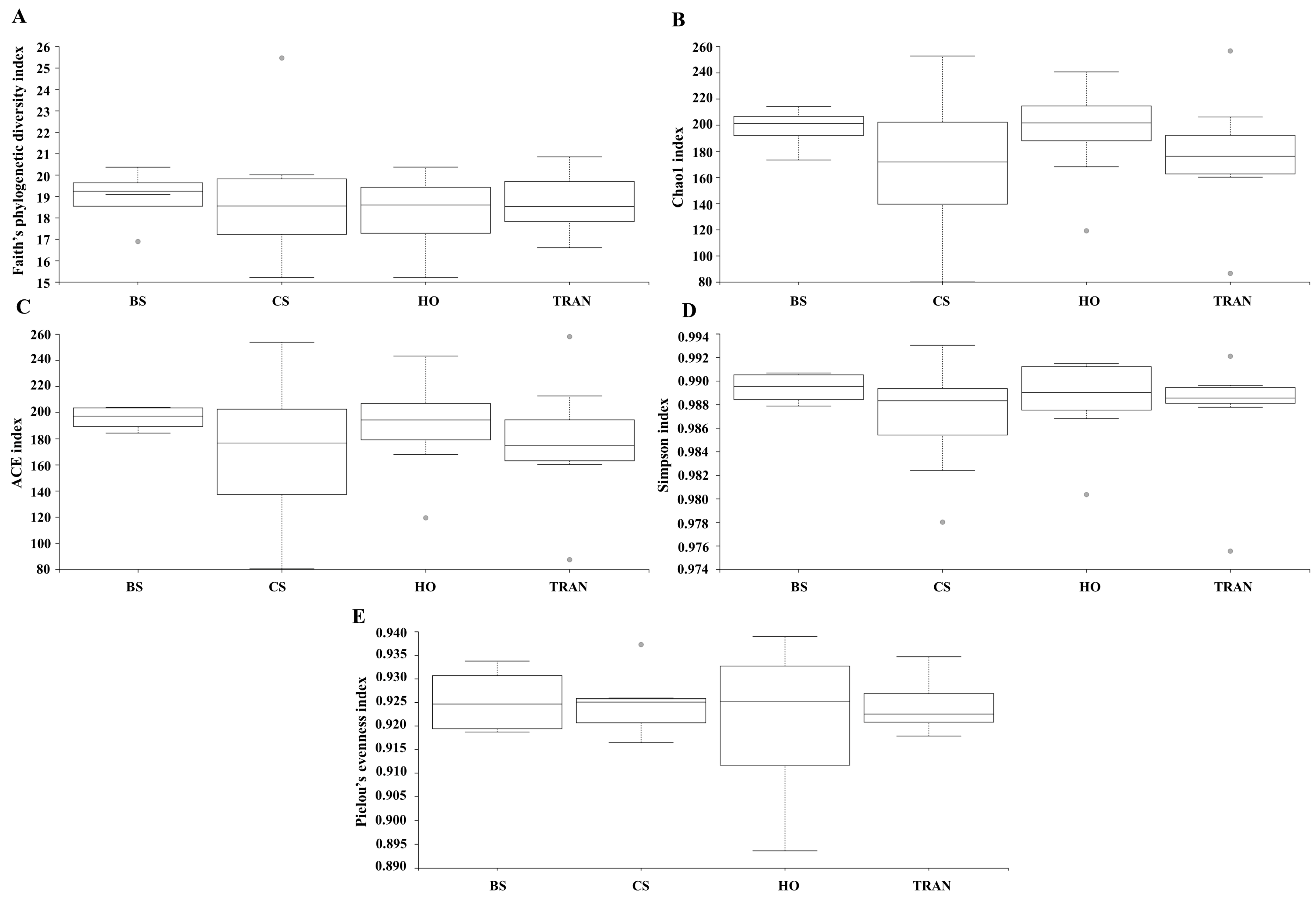
For the community structure of 16S rRNA, the domains Archaea and Bacteria were found from seagrass and soil samples of four sites and four sampling times (Figure 3A and Figure 4A). The abundance of Archaea was high in samples from bare sand, while the abundance of Bacteria was equal in all sites. Our results indicated that Proteobacteria was the most dominant bacterial phyla at 52.24 ± 1.22% (44–65%), followed by Bacteroidetes at 16.39 ± 0.97% (8–31%), Cyanobacteria at 12.12 ± 1.11% (3–28%), and Actinobacteria at 6.08 ± 0.40% (2–10%) (Figure 3A and Figure 4A). Additionally, we found a low relative abundance of Firmicutes, Campilobacterota, Deferribacterota, Desulfobacterota, Acidobacteria, and Myxococcota (<1%). In the bare sand seabed, Proteobacteria and Cyanobacteria dominated in soil samples with 50.22% ± 1.78% and 16.86% ± 1.25%, respectively. For a natural meadow of C. serrulata, Proteobacteria and Bacteroidetes were prevalent across all samples, including soil and seagrasses. The relative abundance of Proteobacteria was 60.61% ± 2.53% and 60.49% ± 2.44% in soil and seagrass samples, respectively. Meanwhile, Bacteroidetes accounted for 15.32% ± 1.52% and 18.82% ± 3.31% in the corresponding sample types. For a natural meadow of H. ovalis, Proteobacteria and Bacteroidetes were abundant across all sample types; the relative abundances of Proteobacteria in soil and seagrass were 53.11% ± 1.28% and 57.59% ± 4.69%, respectively. The relative abundance of Bacteroidetes was around 16.02% ± 1.77% and 18.79% ± 4.27% in soil and seagrasses, respectively. For a transplanted meadow, the relative abundance of Proteobacteria in soil and seagrass was 51.60% ± 3.07% and 55.11% ± 2.73%, respectively. Meanwhile, the relative abundance of Bacteroidetes was around 19.38% ± 1.62% and 17.87% ± 4.70% in soil and seagrass, respectively (Figure 3A and Figure 4A). Species richness, diversity, and evenness are shown in Figure 5A–E. Species richness and species evenness were not significantly different among these four sites (p > 0.05) (Figure 5A–E; Table 1). Species diversity in the natural meadow of C. serrulata (H’ = 6.89) and H. ovalis (H’ = 6.83) and the transplanted meadow (H’ = 6.90) were higher than the bare sand seabed (H’ = 6.90). However, there was no significant difference in species diversity between study sites and months (p > 0.05). The dominant classes were Alphaproteobacteria, Gammaproteobacteria, and Flavobacteriia in all sites. Many classes of bacteria were present only in the H. ovalis site, including Pedosphaerae, 3BR-5F, TM7–3m, and Leptospirae. Solibacteres, MCG, Dehalococcoidetes, and 5bavB12 were highly abundant in the bare sand site. The class Parvarchaea was present only in the transplanted site, while the class Thermoleophilia was present only in the C. serrulata site. Dominant bacterial species included Actibacter sediminis, Ilumatobacter fluminis, Flavobacterium frigidarium, and Lishizhenia caseinilytica. Coccinistipes vermicola, Filomicrobium fusiforme, Desulfobulbus rhabdoformis, Pseudoalteromonas luteoviolacea, and Roseicyclus mahoneyensis were dominant bacterial species in all sites. However, bacterial species composition between sites was not significantly different (p > 0.05) (Figure 6).
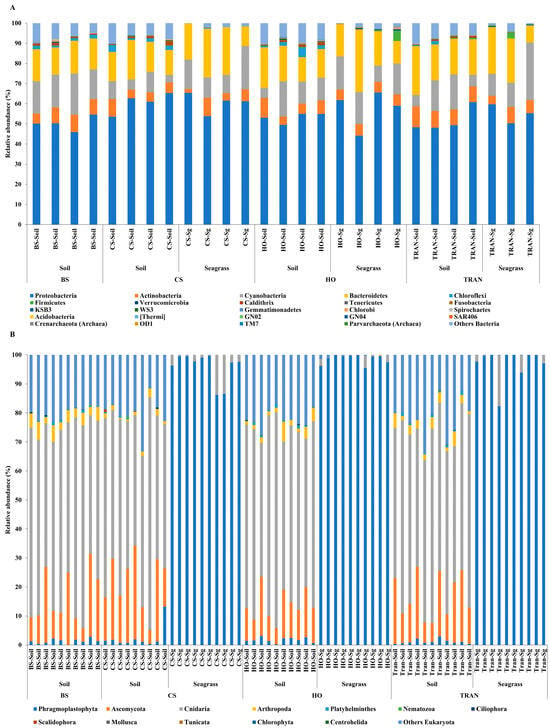
Figure 3.
Relative abundances at the phylum level of (A) 16S rRNA and (B) 18S rRNA in four conditions and two sampling sources. BS = bare sand; CS = Cymodocea serrulata; HO = Halophila ovalis; TRAN = transplanted seagrass patch. Read counts were transformed to percent of relative abundance. For 16s rRNA, blue = Proteobacteria, orange = Actinobacteria, gray = Cyanobacteria, and yellow = Bacteroidetes. For 18s rRNA, blue = Phragmoplastophyta, orange = Ascomycota, gray = Cnidaria, and yellow = Arthropoda. The fewest are organisms detected at less than 1% relative abundance.
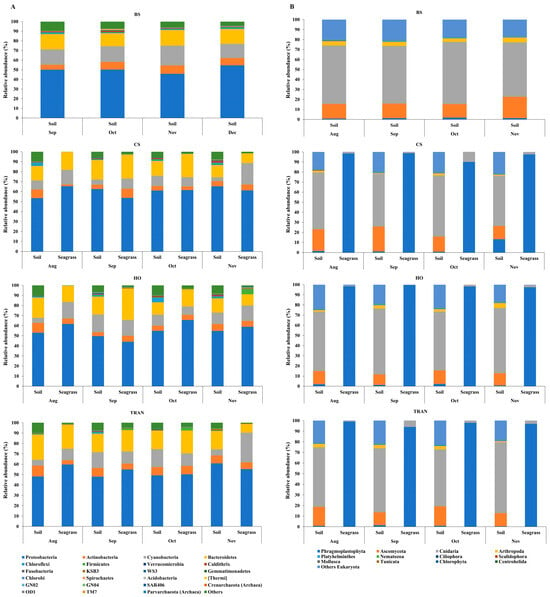
Figure 4.
Relative abundances at the dominant phylum level of (A) 16S rRNA and (B) 18S rRNA of four conditions, two sampling sources, and four sampling times. BS = bare sand; CS = Cymodocea serrulata; HO = Halophila ovalis; TRAN = transplanted seagrass patch. Read counts were transformed to percent relative abundance. For 16s rRNA, blue = Proteobacteria, orange = Actinobacteria, gray = Cyanobacteria, and yellow = Bacteroidetes. For 18s rRNA, blue = Phragmoplastophyta, orange = Ascomycota, gray = Cnidaria, and yellow = Arthropoda. The fewest are organisms detected at less than 1% relative abundance.
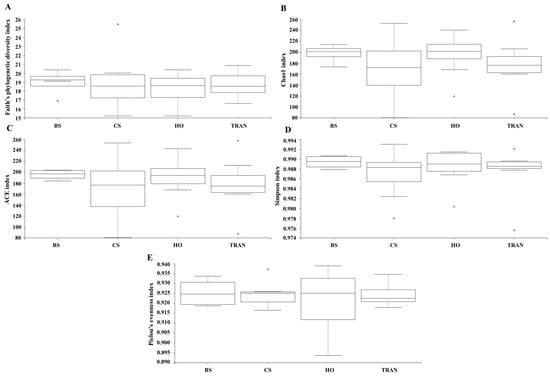
Figure 5.
Species richness (A), species diversity (B–D), and species evenness (E) of 16S rRNA communities in four sites. BS = bare sand; CS = Cymodocea serrulata; HO = Halophila ovalis; TRAN = transplanted seagrass patch. Whiskers indicate the 10th and 90th percentiles.

Table 1.
Output of the pairwise analysis for testing differences in species richness and species evenness of 16S rRNA in four sites. Bare sand; Cymodocea serrulata; Halophila ovalis; transplanted seagrass patch.
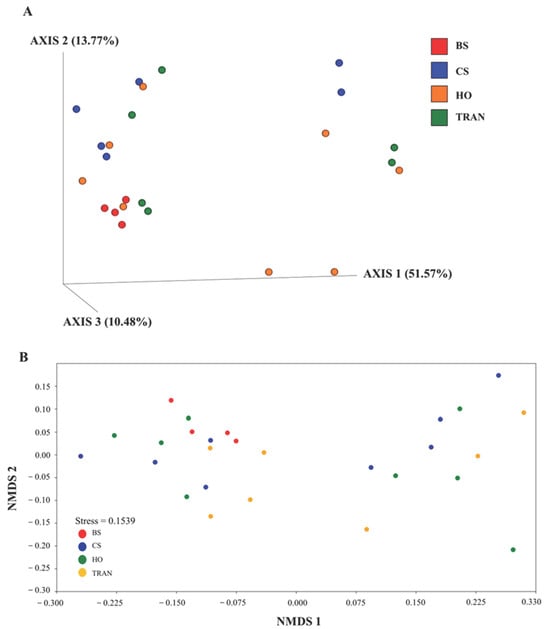
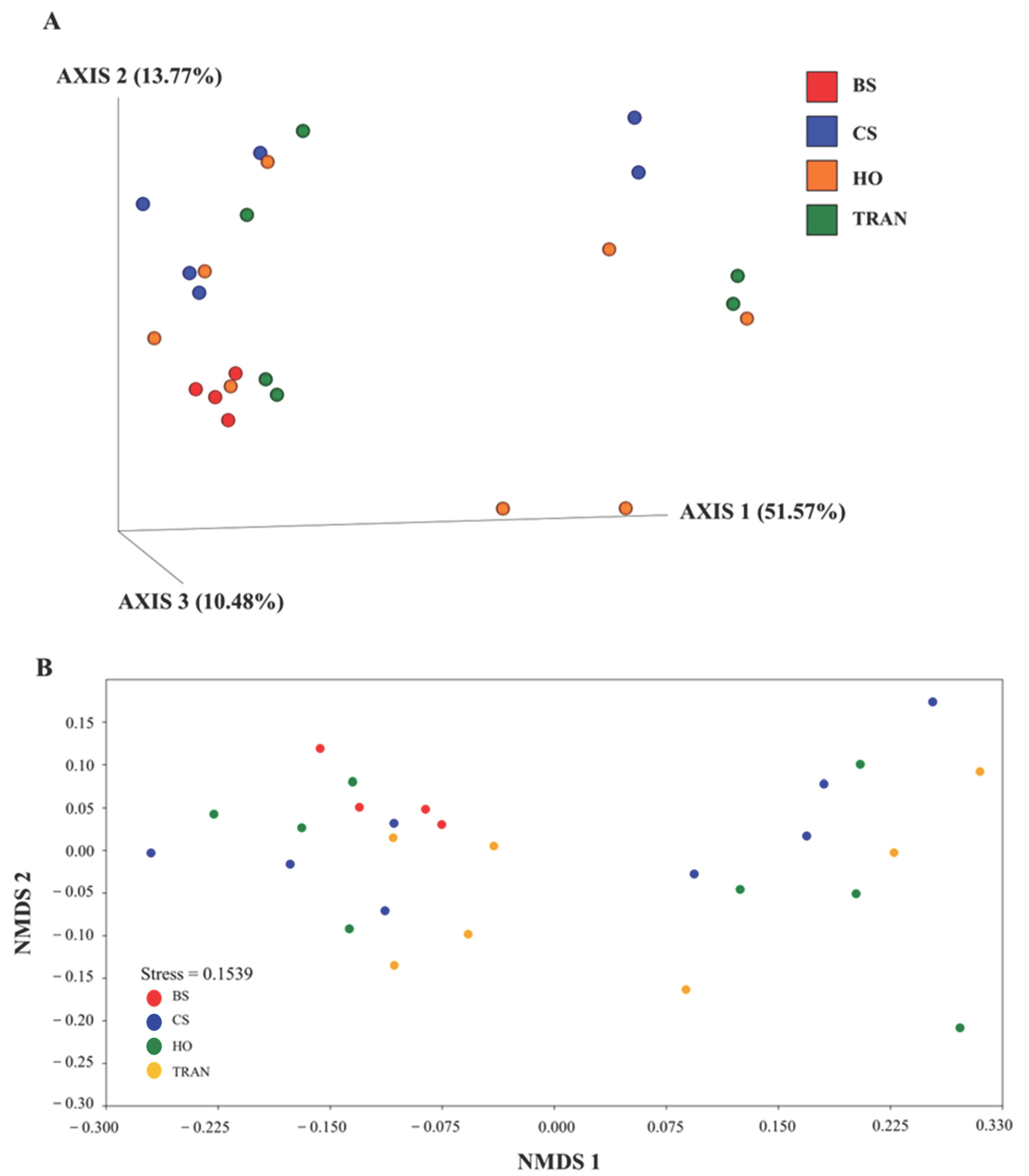
Figure 6.
The PCoA (A) and nMDS (B) of 16S rRNA communities in four sites based on weighted UniFrac distance matrix and Bray–Curtis matrix. BS = bare sand; CS = Cymodocea serrulata; HO = Halophila ovalis; TRAN = transplanted seagrass patch.
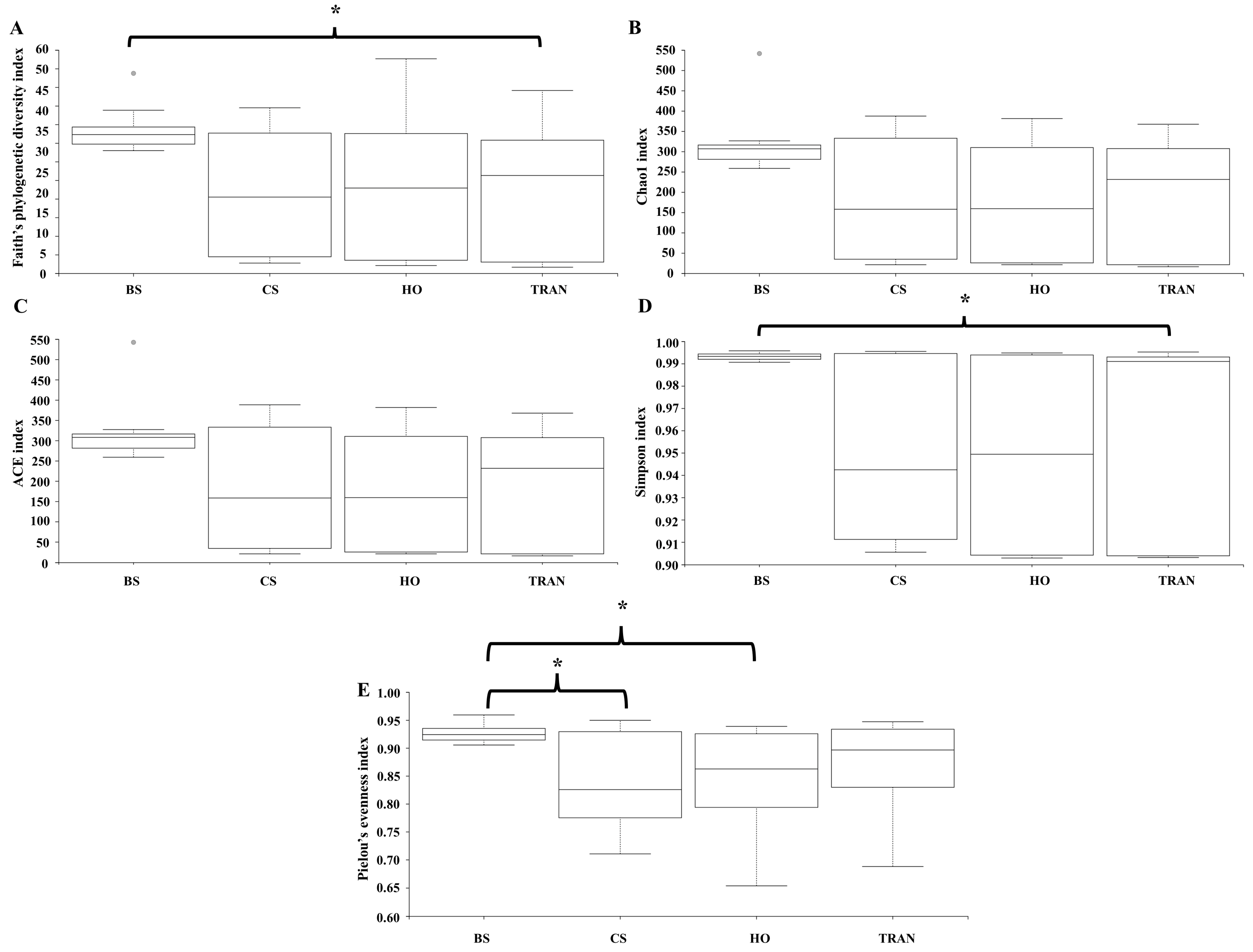
For the community structure of 18S rRNA, Phragmoplastophyta and Cnidaria were the dominant phyla in all conditions with high relative abundances (42.58 ± 5.66% and 34.44 ± 3.40%, respectively) (Figure 3B and Figure 4B). In the bare sand seabed, the dominant phylum in soil samples was Cnidaria at 59.04% ± 2.53% and other Eukaryotes at 19.97% ± 0.07%. For a natural meadow of C. serrulata, soil samples showed a high abundance of Cnidaria at 50.55% ± 3.55% and other Eukaryotes at 18.56% ± 1.95%. Seagrass samples, on the other hand, were dominated by Phragmoplastophyta at 95.95% ± 1.63% and Cnidaria at 3.98% ± 1.63%. For a natural meadow of H. ovalis, soil samples displayed a high abundance of Cnidaria at 50.55% ± 3.55% and other Eukaryotes at 18.56% ± 1.95%. Seagrass samples were dominated by Phragmoplastophyta at 98.60% ± 0.52% and Cnidaria at 0.98% ± 0.46%. For a transplanted meadow, soil samples exhibited high abundances of Cnidaria at 57.54% ± 2.17% and other Eukaryotes at 21.90% ± 2.12%. Seagrass samples were characterized by a high abundance of Phragmoplastophyta at 97.02% ± 1.75% and Cnidaria at 2.82% ± 1.72% (Figure 3B and Figure 4B). Species richness, diversity, and evenness were shown in Figure 7A–E. Our results showed that there was no significant difference in species richness and species evenness among the sites (p = 0.108, 0.068) (Figure 7A,E). There was no significant difference in species richness and evenness between months (p > 0.05). There was, however, a significant difference in species evenness between the bare sand site and the C. serrulata and H. ovalis sites (p = 0.020, 0.039) (Figure 7E). Moreover, the pairwise test showed that there was a significant difference in species diversity between the bare sand site and the transplanted site (p = 0.016) (Table 2). On the H. ovalis, C. serrulata, and transplanted seagrass site, only eight phyla were represented: Phragmoplastophyta, Ascomycota, Cnidaria, Arthropoda, Platyhelminthes, Nematozoa, Ciliophora, Scalidophora, Mollusca, and Tunicata. On the bare sand site, 31 phyla were represented, but the phylum Scalidophora was not among them. Phragmoplastophyta showed a higher relative abundance in all three seagrass sites than in the bare sand site, while Ascomycota and Cnidaria showed a higher relative abundance on bare sand. At the class level, Sordariomycetes was dominant in all conditions, whereas Zygnematophyceae, Intramacronucleata, Trematoda, Arachnida, and Trematoda showed their highest abundances in the bare sand site. Moreover, several classes were represented only in bare sand, including Cryptophyceae, Chrysophyceae, Monogenea, and Perkinsidae. Embryophyta showed the highest abundance in the H. ovalis site, while Hydrozoa showed its highest abundance in the C. serrulata site. Gastropoda and Malacostraca were highly abundant in the transplanted seagrass site.
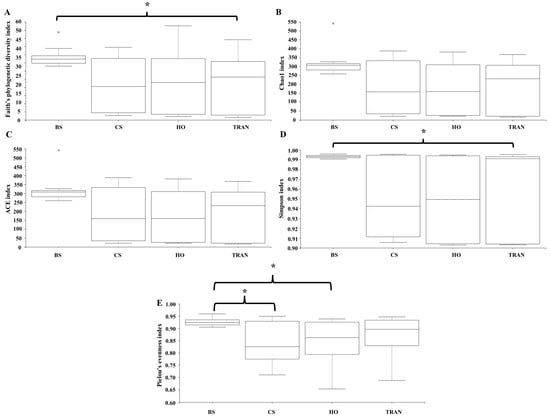
Figure 7.
Species richness (A), species diversity (B–D), and species evenness (E) of 18S rDNA communities in four sites. BS = bare sand; CS = Cymodocea serrulata; HO = Halophila ovalis; TRAN = transplanted seagrass patch. Whiskers indicate the 10th and 90th percentiles. * = p < 0.05.
Table 2.
Output of the pairwise analysis for testing differences in species richness and species evenness of 18S rRNA in four sites. Bare sand; Cymodocea serrulata; Halophila ovalis; transplanted seagrass patch.
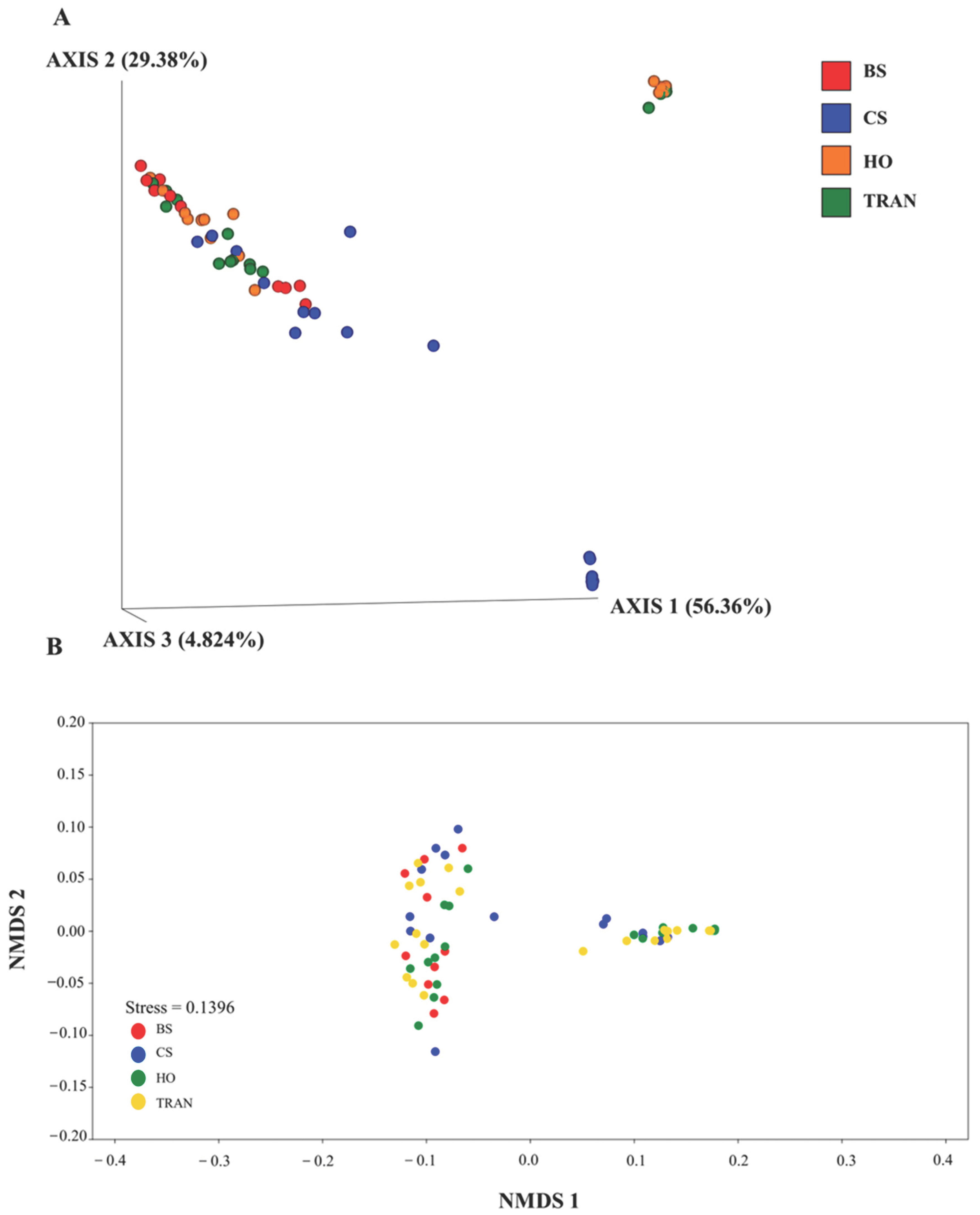
PCoA illustrated that there was significant difference in species composition of the 18S rRNA community among the four sites (p = 0.0289) (Figure 8). Xylariaceae spp. were dominant in all sites. However, many species showed similar abundances in the natural and transplanted seagrass sites. These species included Priapulus caudatus, Obelia dichotoma, Leptothecata, Phyllidiella pustulosa, Stauridiosarsia ophiogaster, Paraberrapex manifestus, Plagiopyla plagiopylid, and Trichostomatia. Meanwhile, some species were present only in bare sand. These species included Cosmarium protractum, Cryptomonas, Lacinularia flosculosa, and Pterocystis tropica.
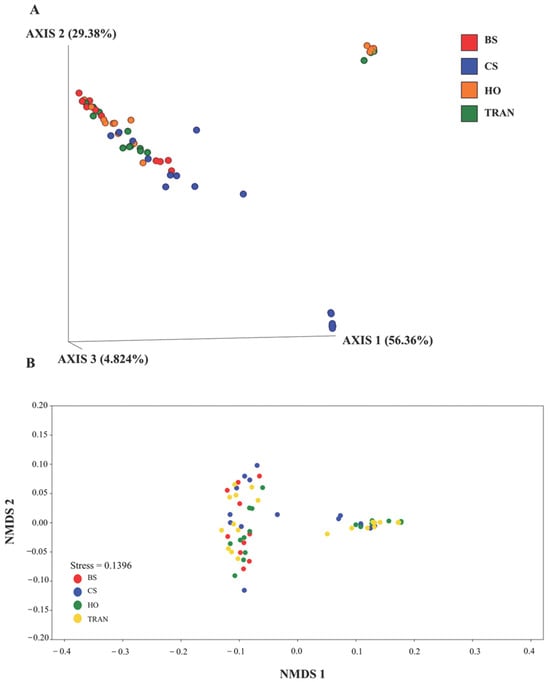
Figure 8.
The PCoA (A) and nMDS (B) of 18S rRNA communities in four sites based on weighted UniFrac distance matrix and Bray–Curtis matrix. BS = bare sand; CS = Cymodocea serrulata; HO = Halophila ovalis; TRAN = transplanted seagrass patch.
In this study, dugong trails and commercially valuable species such as sea cucumber, Holuthuria (Metriatyla) scabra Jaejer, 1833, Dog conch, Laevistrombus canarium (Linnaeus, 1758), and swimming crab, Portunus sp. have been observed in the natural and transplanted seagrass sites.
4. Discussion
Using eDNA, we assessed and compared the diversity of marine organisms among four sites: (1) a bare sand seabed, (2) a natural meadow of Cymodocea serrulata, (3) a natural meadow of Halophila ovalis, and (4) a patch of transplanted seagrass.
For the community structure of 16S rRNA, Proteobacteria, Bacteroidetes, Actinobacteria, and Cyanobacteria were dominant bacterial groups at all sites and months. Similar results have been reported in that they are widespread across marine habitats such as mangrove forests, coral reefs, seawater, and seagrass meadows [39,40,41,42,43,44,45,46]. In addition, these phyla are common and associated with epilithic algae or turf algae [47]. Moreover, many studies reported that these four dominant phyla were also important groups in other seagrass and seaweed habitats, protecting the host from pathogens and inhibiting fungal infection by facilitating immune responses [48,49,50,51,52]. Additionally, microbes can supply hosts with nitrogen sources and protect host from herbivory by secreting chemical defenses against herbivores [44,45,46,47,48,49]. In this study, we found that the domain Archaea, which is normally absent or present with low abundance in marine environments [53,54], was present with high abundance in bare sand. Our results were similar to the study of Zheng et al. [55], who found that Archaea is dominant in the seagrass system and bare sediments. Their work showed that the different dominant phyla of Archaea prefer different environments. Some phylum of Archaea such as phylum Bathyarchaeota was enriched in seagrass-colonized sediments, while Woesearchaeota showed similar relative proportions in colonized seagrass and bare sediments. Additionally, it has been reported that Archaea can be detected in shallow or anaerobic sediments. It has been suggested that Archaea contribute to carbon and anaerobic nitrogen cycling [55].
The dominant bacterial classes in our study were Gammaproteobacteria and Alphaproteobacteria. These two classes were recently reported to be well represented in the sediment of bare sand, in seagrass-colonized areas, and on healthy seagrass leaves [56,57]. Moreover, these classes of bacteria were common in all parts of seagrass from root to leaf, fruit, and flower [58]. They were widely distributed and abundant because of their high abundance in seawater and their resistance to environmental changes [59]. These two classes can enhance seagrass growth by providing nutrients and play a role in nutrient cycling. At the species level, Haliea mediterranea, Desulfopila aestuarii, Thioalkalivibrio, and Spongiibacter tropicus were dominant in all sites. These four dominant organic-matter cycling bacterial groups have been reported as common bacteria distributed in the marine environment [60,61,62,63]. Rabbani et al. [64] reported that bacterial genera can be an indicator of healthy and stressed seagrass meadows, observing that nutrient-cycling bacteria are frequently found and are abundant in healthy seagrass meadows, while sulfur-cycling bacteria are more detected in stressed seagrass meadows.
There were no significant differences in bacterial species evenness and diversity among all three seagrass patches and the bare sand site. This might be because of the suitable site and technique used. In our selected seagrass transplantation site, seagrass species had previously been present. Additionally, this site was close to the donor site, which had similar environmental conditions such as water depth, wave current, sediment type, and light intensity. For the transplantation technique, we used the plug method that can extract seagrasses with sediment and rhizomes intact. This attempt can help seagrasses, benthic fauna, and microbial communities adapt to the new environments of the transplantation site. So, their communities did not differ between the donor and the transplanted sites. However, it has been reported that microbial communities are complex and vary over spatial and temporal scales. Rabbani et al. [64] mentioned that the microbial community structures can differ among sampling sites, with living seagrass parts showing that below-ground parts (rhizome and root) and sediment had more similar bacterial diversity than those above ground. It has been recommended that microbial community structure and dynamics in seagrass restoration should be considered as important keys for seagrass restoration and management success. However, caution must be applied since relic DNA from dead microorganisms that can persist in the environment [65] could be responsible for mis-estimations of up to 40%.
For the community structure of 18S rRNA, Ascomycota, Cnidaria, Phragmoplastophyta/Charophyta, and Arthropoda were the dominant eukaryotic phyla. Ascomycota was found in seagrass roots and marine sediment [66,67,68]. Several studies reported that Ascomycota was an important fungal phylum that plays a key role in the biogeochemical cycles and the decomposition of organic substrates in the ecosystem [66,69]. The phylum Cnidaria was also well represented in all sites. However, there was a higher abundance of Cnidaria in bare sand compared with the seagrass sites. This finding was different from that of Wesselmann et al. [70] that there was a high abundance of Cnidaria in seagrass meadows because of the complex structural habitat that seagrass provides. Barnes [71] also showed that abundance and richness of macrofauna were higher in seagrass meadows than in bare sand. Phragmoplastophyta or Charophyta were the dominant phyla in the seagrass sites in the present study. Since Phragmoplastophyta are green epiphytic algae that normally attach to seagrasses [72,73], there was a high abundance of Phragmoplastophyta in the seagrass samples that we collected. Malacostraca in phylum Arthropoda had high abundance in the transplanted seagrass site, which was an important habitat, nursery ground, and refuge for marine fauna.
Our results revealed that relative abundance among the four sites was significantly different. Species richness and evenness of fauna between the natural and transplantation meadows were not different. Their abundance, richness, and evenness in transplantation meadows might recover and become level with the natural meadows. It seems that fauna are not limited in their capacity to move between the natural and transplanted habitats. From our observation, the transplanted meadows could expand the area from around 2.12 m2 of total transplanted meadows to around 55.33 m2 after 9 months of restoration. We found that these transplanted meadows might attract and provide ecosystem services (such as food, habitat, and refuge) for marine organisms similar to those of natural seagrass meadows, increasing diversity [17,18]. These results are similar to the results of Gagnon et al. [74], showing that faunal density and biodiversity of planted eelgrass (Zostera marina) plots were similar to the reference meadow after 15 months of restoration. Restored seagrass meadows rapidly become habitat and can provide important services such as provisioning, regulating, and supporting services to diverse fauna.
In this study, dugong trails and commercially valuable species such as sea cucumber, Holuthuria (Metriatyla) scabra Jaejer, 1833, dog conch, Laevistrombus canarium (Linnaeus, 1758), and swimming crab, Portunus sp. have been observed in the natural and transplanted seagrass sites. Unfortunately, the DNA of these fauna were not detected. The diversity we reported in this study might be underestimated. It could be because DNA samples were easily degraded or the DNA of these fauna was not detectable because they may not frequently visit these sites. Additionally, the successful identification of eDNA is limited. The classification relies on reference databases, some of which may not be in reference libraries [70]. So, complete and comprehensive reference databases are important and needed. In addition, the combination of eDNA, underwater video, and visual site survey during low tide for biodiversity assessment may provide a more holistic view of marine fauna and flora.
This study provides an overall assessment of the biodiversity of a restored seagrass area compared with donor areas and a bare sand area. Restored seagrass meadow can be successfully established, create habitat, and provide food for fauna after restoration. The plug method was a successful method for our restoration. It is easy in soft substrates with small and thin-leaved seagrass species. The plug method seems suitable for a small area where the donor site is close to the transplanted site. For larger transplantations, other methods could be considered.
To get a better understanding of seagrass ecosystem services after restoration, long-term monitoring and more comprehensive data from combining eDNA and traditional assessments such as visual survey and underwater video are carefully considered to assess the local diversity of marine organisms and the success of restoration programs. Additionally, carbon sequestration and organic matter cycling by the restored seagrass meadow are needed to evaluate comparisons with the donor meadows and the bare sand area.
Author Contributions
Conceptualization, J.M., P.T. and E.R.; methodology, J.M., K.S., P.T. and E.R.; software, K.S., J.T. and J.M.; formal analysis, J.T. and K.S.; investigation, J.M., P.T., E.R. and J.T.; data curation, K.S.; writing—original draft preparation, J.T. and J.M.; writing—review and editing, J.T. and J.M.; visualization, J.T. and K.S.; supervision, J.M.; project administration, J.M., P.T. and E.R.; funding acquisition, J.M., P.T. and E.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Research Council of Thailand (NRCT) (No. 306/2021).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available on request.
Acknowledgments
Many thanks to the Seaweed and Seagrass Research Unit (SSRU) of Prince of Songkla University (PSU) for help with field work, as well as to Thomas Duncan Coyne for assistance with the English text.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cullen-Unsworth, L.; Unsworth, R. Seagrass meadows, ecosystem services, and sustainability. Environ. Sci. Policy 2013, 55, 14–28. [Google Scholar] [CrossRef]
- Ruiz-Frau, A.; Gelcich, S.; Hendriks, I.E.; Duarte, C.M.; Marbà, N. Current state of seagrass ecosystem services: Research and policy integration. Ocean. Coast. Manag. 2017, 149, 107–115. [Google Scholar] [CrossRef]
- Reusch, T.B.; Schubert, P.R.; Marten, S.M.; Gill, D.; Karez, R.; Busch, K.; Hentschel, U. Lower Vibrio spp. abundances in Zostera marina leaf canopies suggest a novel ecosystem function for temperate seagrass beds. Mar. Biol. 2021, 168, 149. [Google Scholar] [CrossRef]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K., Jr.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe Threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef]
- Robert, J.; Wong, P.P. Coastal systems and low-lying areas. In Climate Change 2007: Impacts, Adaptation and Vulnerability; Parry, M., Canziani, O., Eds.; Cambridge University Press: New York, NY, USA, 2007; pp. 326–367. [Google Scholar]
- Sala, E.; Knowlton, N. Global marine biodiversity trends. Annu. Rev. Environ. Resour. 2006, 31, 93–122. [Google Scholar] [CrossRef]
- Dunlop, M. Biodiversity: Strategy conservation. Nat. Clim. Chang. 2013, 3, 1019–1020. [Google Scholar] [CrossRef]
- Koch, M.; Bowes, G.; Ross, C.; Zhang, X.H. Climate change and ocean acidification effects on seagrasses and marine macroalgae. Glob. Chang. Biol. 2013, 19, 103–132. [Google Scholar] [CrossRef]
- Rasheed, M.; McKenna, S.A.; Carter, A.B.; Coles, R. Contrasting recovery of shallow and deep-water seagrass communities following climate associated losses in tropical north Queensland, Australia. Mar. Pollut. Bull. 2014, 83, 491–499. [Google Scholar] [CrossRef]
- Reed, B.J.; Hovel, K.A. Seagrass habitat disturbance: How loss and fragmentation of, eelgrass Zostera marina influences epifaunal abundance and diversity. Mar. Ecol. Prog. Ser. 2006, 326, 133–143. [Google Scholar] [CrossRef]
- Irlandi, E.A.; Orlando, B.A.; Ambrose, G.C., Jr. Influence of seagrass habitat patch size on growth and survival of juvenile bay scallops, Argopecten irradians concentricus (Say). J. Exp. Mar. Biol. Ecol. 1999, 235, 21–43. [Google Scholar] [CrossRef]
- Spivak, A.C.; Canuel, E.A.; Duffy, J.E.; Richardson, J.P. Top-down and bottom-up controls on sediment organic matter composition in an experimental seagrass ecosystem. Limnol. Oceanogr. 2007, 52, 2595–2607. [Google Scholar] [CrossRef]
- Bryars, C.; Neverauskas, V. Natural recolonization of seagrasses at a disused sewage sludge outfall. Aqua. Bot. 2004, 80, 283–289. [Google Scholar] [CrossRef]
- Riemann, B.; Carstensen, J.; Dahl, K.; Fossing, H.; Hansen, J.W.; Jakobsen, H.H.; Josefson, A.B.; Krause-Jensen, D.; Markager, S.; Stæhr, P.A.; et al. Recovery of Danish coastal ecosystems after reductions in nutrient loading: A holistic ecosystem approach. Estuaries Coasts 2016, 39, 82–97. [Google Scholar] [CrossRef]
- Orth, R.J.; McGlathery, J.K. Eelgrass recovery in the coastal bays of the Virginia Coast Reserve, USA. Mar. Ecol. Prog. Ser. 2012, 448, 173–176. [Google Scholar] [CrossRef]
- Matheson, F.E.; Reed, J.; Dos Santos, V.M.; Mackay, G.; Cummings, V.J. Seagrass rehabilitation: Successful transplants and evaluation of methods at different spatial scales. N. Z. J. Mar. Freshw. Res. 2016, 51, 96–109. [Google Scholar] [CrossRef]
- Blandon, A.; Zu Ermgassen, P.S. Quantitative estimate of commercial fish enhancement by seagrass habitat in southern Australia. Coast. Shelf Sci. 2014, 141, 1–8. [Google Scholar] [CrossRef]
- Lefcheck, J.S.; Marion, S.R.; Orth, R.J. Restored Eelgrass (Zostera marina L.) as a refuge for epifaunal biodiversity in mid-western Atlantic coastal bays. Estuaries Coast 2017, 40, 200–212. [Google Scholar] [CrossRef]
- Ondiviela, B.; Losada, I.J.; Lara, J.L.; Maza, M.; Galván, C.; Bouma, T.I.; van Belzen, J. The role of seagrasses in coastal protection in a changing climate. Coast. Eng. 2014, 87, 158–168. [Google Scholar] [CrossRef]
- Reynolds, L.K.; Waycott, M.; McGlathery, K.J.; Orth, R.J. Ecosystem services returned through seagrass restoration. Restor. Ecol. 2016, 24, 583–588. [Google Scholar] [CrossRef]
- Orth, R.J.; Lefcheck, J.S.; McGlathery, K.S.; Aoki, L.; Luckenbach, M.W.; Moore, K.A.; Oreska, M.; Snyder, R.; Wilcox, D.J.; Lusk, B. Restoration of seagrass habitat leads to rapid recovery of coastal ecosystem services. Sci. Adv. 2020, 6, eabc6434. [Google Scholar] [CrossRef] [PubMed]
- van Katwijk, M.M.; Thorhaug, A.; Marbà, N.; Orth, R.J.; Duarte, C.M.; Kendrick, G.A.; Althuizen, I.H.; Balestri, E.; Bernard, G.; Cambridge, M.L.; et al. Global analysis of seagrass restoration: The importance of large-scale planting. J. Appl. Ecol. 2016, 53, 567–578. [Google Scholar] [CrossRef]
- Valdemarsen, T.; Canal-Vergés, P.; Kristensen, E.; Holmer, M.; Kristiansen, M.; Flindt, M.V.R. Vulnerability of Zostera marina seedlings to physical stress. Mar. Ecol. Prog. Ser. 2010, 418, 119–130. [Google Scholar] [CrossRef]
- Carstensen, J.; Krause-Jensen, D.; Markager, S.; Timmermann, K.; Windolf, J. Water clarity and eelgrass responses to nitrogen reductions in the eutrophic Skive Fjord, Denmark. Hydrobiologia 2013, 704, 293–309. [Google Scholar] [CrossRef]
- He, H.; Stanley, R.R.E.; Rubidge, E.M.; Jeffery, N.W.; Hamilton, L.C.; Westfall, K.M.; Gilmore, S.R.; Roux, L.M.D.; Gale, K.S.P.; Heaslip, S.G.; et al. Fish community surveys in eelgrass beds using both eDNA metabarcoding and seining: Implications for biodiversity monitoring in the coastal zone. Can. J. Fish. Aquat. Sci. 2022, 79, 1335–1346. [Google Scholar] [CrossRef]
- Stankovic, M.; Hayashizaki, K.I.; Tuntiprapas, P.; Rattanachot, E.; Prathep, A. Two decades of seagrass area change: Organic carbon sources and stock. Mar. Pollut. Bull. 2021, 163, 111913. [Google Scholar] [CrossRef]
- Kaewsrikhaw, R.; Upanoi, T.; Prathep, A. Ecosystem services and vulnerability assessments of seagrass ecosystems: Basic tools for prioritizing conservation management actions using an example from Thailand. Water 2022, 14, 3650. [Google Scholar] [CrossRef]
- Tongnunui, P.; Tarangkoon, W.; Hukiew, P.; Kaeoprakan, P.; Horinouchi, M.; Rojchanaprasart, N.; Ponpai, W. Seagrass restoration: An update from Trang province, Southwestern Thailand. In Proceedings of the International Conference “Managinag Risks to Coastal Regions and Communities in a Changinag World” (EMECS’11—SeaCoasts XXVI), Saint-Petersburg, Russia, 22–27 August 2017; Academus Publishing: Redwood City, CA, USA, 2017; pp. 10–12. [Google Scholar]
- Thomsen, P.F.; Willerslev, E. Environmental DNA—An emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 2015, 183, 4–18. [Google Scholar] [CrossRef]
- Guardiola, M.; Uriz, M.J.; Taberlet, P.; Coissac, E.; Wangensteen, O.S.; Turon, X. Correction: Deep-sea, deep-sequencing: Metabarcoding extracellular DNA from sediments of marine canyons. PLoS ONE 2016, 11, e0153836. [Google Scholar] [CrossRef]
- Ortega, A.; Geraldi, N.R.; Duarte, C.M. Environmental DNA identifies marine macrophyte contributions to blue carbon sediments. Limnol. Oceanogr. 2013, 65, 3139–3149. [Google Scholar] [CrossRef]
- Closek, C.J.; Santora, J.A.; Starks, H.A.; Schroeder, I.D.; Andruszkiewicz, E.A.; Sakuma, K.M.; Bograd, S.J.; Hazen, E.L.; Field, J.C.; Boehm, A.B. Marine vertebrate biodiversity and distribution within the Central California current using environmental DNA (eDNA) metabarcoding and ecosystem surveys. Front. Mar. Sci. 2019, 6, 732. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucl. Acids. Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Pielou, E.C. The Measurement of diversity in different types of biological collections. J. Theor. Biol. 1996, 13, 131–144. [Google Scholar] [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef]
- Somboonna, N.; Assawamakin, A.; Wilantho, A.; Tangphatsornruang, S.; Tongsima, S. Structural and functional diversity of free-living microorganisms in reef surface, Kra island, Thailand. BMC Genom. 2014, 15, 607. [Google Scholar] [CrossRef]
- Liang, J.; Yu, K.; Wang, Y.; Huang, X.; Huang, W.; Qin, Z.; Pan, Z.; Yao, Q.; Wang, W.; Wu, Z. Distinct bacterial communities associated with massive and branching Scleractinian corals and potential linkages to coral susceptibility to thermal or cold stress. Front. Microbiol. 2017, 8, 979. [Google Scholar] [CrossRef]
- Pootakham, W.; Mhuantong, W.; Yoocha, T.; Putchim, L.; Sonthirod, C.; Naktang, C.; Thongtham, N.; Tangphatsornruang, S. High resolution profiling of coral-associated bacterial communities using full-length 16S rRNA sequence data from PacBio SMRT sequencing system. Sci. Rep. 2017, 7, 2774. [Google Scholar] [CrossRef]
- Somboonna, N.; Wilantho, A.; Monanunsap, S.; Chavanich, S.; Tangphatsornruang, S.; Tongsima, S. Microbial communities in the reef water at Kham Island, lower Gulf of Thailand. PeerJ 2017, 5, e3625. [Google Scholar] [CrossRef]
- Mhuantong, W.; Nuryadi, H.; Trianto, A.; Sabdono, A.; Tangphatsornruang, S.; Eurwilaichitr, L.; Kanokkratana, P.; Champreda, V. Comparative analysis of bacterial communities associated with healthy and diseased corals in the Indonesian sea. PeerJ 2019, 7, e8137. [Google Scholar] [CrossRef]
- Pollock, F.J.; Lamb, J.B.; van de Water, J.A.; Smith, H.A.; Schaffelke, B.; Willis, B.L.; Bourne, D.G. Reduced diversity and stability of coral-associated bacterial communities and suppressed immune function precedes disease onset in corals. R. Soc. Open Sci. 2019, 6, 190355. [Google Scholar] [CrossRef]
- Nimnoi, P.; Pongsilp, N. Marine bacterial communities in the upper gulf of Thailand assessed by Illumina next-generation sequencing platform. BMC Microbiol. 2020, 20, 19. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Q.; Ling, J.; Long, L.; Huang, H.; Yin, J.; Wu, M.; Tang, X.; Lin, X.; Zhang, Y.; et al. Shifting the microbiome of a coral holobiont and improving host physiology by inoculation with a potentially beneficial bacterial consortium. BMC Microbiol. 2021, 21, 130. [Google Scholar] [CrossRef]
- Titioatchasai, J.; Surachat, K.; Kim, J.H.; Mayakun, J. Diversity of microbial communities associated with epilithic macroalgae in different coral reef conditions and damselfish territories of the Gulf of Thailand. J. Mar. Sci. Eng. 2023, 11, 514. [Google Scholar] [CrossRef]
- Jensen, S.I.; Kühl, M.; Priemé, A. Different bacterial communities associated with the roots and bulk sediment of the seagrass Zostera marina. FEMS Microbiol. Ecol. 2007, 62, 108–117. [Google Scholar] [CrossRef]
- Marhaeni, B.; Radjasa, O.K.; Bengen, D.G.; Kaswadji, R.F. Screening of bacterial symbionts of seagrass Enhalus sp. against biofilm-forming bacteria. J. Coast. Dev. 2010, 13, 126–132. [Google Scholar]
- Ugarelli, K.; Laas, P.; Stingl, U. The microbial communities of leaves and roots associated with turtle Grass (Thalassia testudinum) and manatee grass (Syringodium filliforme) are distinct from seawater and sediment communities, but are similar between species and sampling sites. Microorganisms 2018, 7, 4. [Google Scholar] [CrossRef]
- Martin, B.C.; Alarcon, M.S.; Gleeson, D.; Middleton, J.A.; Fraser, M.W.; Ryan, M.H.; Holmer, M.; Kendrick, G.A.; Kilminster, K. Root microbiomes as indicators of seagrass health. FEMS Microbiol. Ecol. 2020, 96, fiz201. [Google Scholar] [CrossRef]
- Iqbal, M.M.; Nishimura, M.; Haider, M.N.; Sano, M.; Ijichi, M.; Kogure, K.; Yoshizawa, S. Diversity and composition of microbial communities in an eelgrass (Zostera marina) bed in Tokyo Bay, Japan. Microbes Environ. 2021, 36, ME21037. [Google Scholar] [CrossRef]
- Wegley, L.; Edwards, R.; Rodriguez-Brito, B.; Liu, H.; Rohwer, F. Metagenomic analysis of the microbial community associated with the coral Porites astreoides. Environ. Microbiol. 2007, 9, 2707–2719. [Google Scholar] [CrossRef] [PubMed]
- Bier, E.J.; Sun, S.; Howard, E.C. Prokaryotic genomes and diversity in surface ocean waters: Interrogating the Global ocean sampling metagenome. Appl. Environ. Microbiol. 2009, 75, 2221–2229. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Wang, C.; Zhang, X.; Gong, J. Community structure and abundance of Archaea in a Zostera marina Meadow: A Comparison between seagrass-colonized and bare sediment sites. Archaea 2019, 2019, 5108012. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.L.; Wang, X.L.; Li, Y.C. The influence of eefgrass (Zostera marina L.) on the environmental factors and the microbial communities in sediment from Shuangdao bay of China. Appl. Ecol. Environ. Res. 2019, 9, 10767–11077. [Google Scholar]
- Banister, R.B.; Schwarz, M.T.; Fine, M.; Ritchie, K.B.; Muller, E.M. Instability and stasis among the microbiome of seagrass leaves, roots and rhizomes, and nearby sediments within a natural pH gradient. Microb. Ecol. 2022, 84, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Tarquinio, F.; Attlan, O.; Vanderklift, M.A.; Berry, O.; Bissett, A. Distinct endophytic bacterial communities inhabiting seagrass seeds. Front. Microbiol. 2021, 12, 703014. [Google Scholar] [CrossRef] [PubMed]
- Conte, C.; Rotini, A.; Manfra, L.; D’Andrea, M.M.; Winters, G.; Migliore, L. The seagrass holobiont: What we know and what we still need to disclose for its possible use as an ecological indicator. Water 2021, 13, 406. [Google Scholar] [CrossRef]
- Suzuki, D.; Ueki, A.; Amaishi, A.; Ueki, K. Desulfopila aestuarii gen. nov., sp. nov., a Gram-negative, rod-like, sulfate-reducing bacterium isolated from an estuarine sediment in Japan. Int. J. Syst. Evol. Microbiol. 2007, 57, 520–526. [Google Scholar] [CrossRef]
- Sundarakrishnan, B.; Pushpanathan, M.; Jayashree, S.; Rajendhran, J.; Sakthivel, N.; Jayachandran, S.; Gunasekaran, P. Assessment of microbial richness in pelagic sediment of Andaman Sea by bacterial Tag Encoded FLX Titanium Amplicon Pyrosequencing (bTEFAP). Indian J. Microbiol. 2012, 52, 544–550. [Google Scholar] [CrossRef]
- Guevara, R.; Ikenaga, M.; Dean, A.L.; Pisani, C.; Boyer, J.N. Changes in sediment bacterial community in response to long-term nutrient enrichment in a subtropical seagrass-dominated estuary. Microb. Ecol. 2014, 68, 427–440. [Google Scholar] [CrossRef]
- Spring, S.; Scheuner, C.; Göker, M.; Klenk, H.P. A taxonomic framework for emerging groups of ecologically important marine gammaproteobacteria based on the reconstruction of evolutionary relationships using genome-scale data. Front. Microbiol. 2015, 6, 281. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, G.; Yan, B.C.; Lee, N.L.Y.; Ooi, J.L.S.; Lee, J.N.; Huang, D.; Wainwright, B.J. Spatial and structural factors shape seagrass-associated bacterial communities in Singapore and Peninsular Malaysia. Front. Mar. Sci. 2021, 8, 659180. [Google Scholar] [CrossRef]
- Carini, P.; Marsden, P.J.; Leff, J.W.; Morgan, E.E.; Strickland, M.S.; Fierer, N. Relic DNA is abundant in soil and obscures estimates of soil microbial diversity. Nat. Microbiol. 2017, 2, 16242. [Google Scholar] [CrossRef] [PubMed]
- Panno, L.; Bruno, M.; Voyron, S.; Anastasi, A.; Gnavi, G.; Miserere, L.; Varese, G.C. Diversity, ecological role and potential biotechnological applications of marine fungi associated to the seagrass Posidonia oceanica. New Biotechnol. 2013, 30, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Vohník, M.; Borovec, O.; Kolarík, M. Communities of cultivable root mycobionts of the seagrass Posidonia oceanica in the northwest Mediterranean Sea are dominated by a hitherto undescribed pleosporalean dark septate endophyte. Microb. Ecol. 2016, 71, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Orfali, R.; Perveen, S.; Khan, M.F.; Ahmed, A.F.; Wadaan, M.A.; Al-Taweel, A.M.; Alqahtani, A.S.; Nasr, F.A.; Tabassum, S.; Luciano, P.; et al. Antiproliferative Illudalane Sesquiterpenes from the Marine Sediment Ascomycete Aspergillus oryzae. Mar. Drugs 2021, 19, 333. [Google Scholar] [CrossRef] [PubMed]
- Zuccaro, A.; Schoch, C.L.; Spatafora, J.W.; Kohlmeyer, J.; Draeger, S.; Mitchell, J.I. Detection and identification of fungi intimately associated with the brown seaweed Fucus serratus. Appl. Environ. Microbiol. 2008, 74, 931–941. [Google Scholar] [CrossRef]
- Wesselmann, M.; Geraldi, N.R.; Marbà, N.; Hendriks, I.E.; Díaz-Rúa, R.; Duarte, C.M. eDNA reveals the associated metazoan diversity of mediterranean seagrass sediments. Diversity 2022, 14, 549. [Google Scholar] [CrossRef]
- Barnes, R.F. Patterns of benthic invertebrate biodiversity in intertidal seagrass in Moreton Bay, Queensland. Reg. Stud. Mar. Sci. 2017, 15, 17–25. [Google Scholar] [CrossRef]
- Hassenrück, C.; Hofmann, L.C.; Bischof, K.; Ramette, A. Seagrass biofilm communities at a naturally CO2-rich vent. Environ. Microbiol. Rep. 2015, 7, 516–525. [Google Scholar] [CrossRef]
- Tsioli, S.; Vasillis, P.; Anastasia, R.; Maria, K.; Christos, K.; Eva, P.; Küpper, F.C.; Sotiris, O. Diversity and composition of algal epiphytes on the Mediterranean seagrass Cymodocea nodosa: A scale-based study. Bot. Mar. 2021, 64, 101–118. [Google Scholar] [CrossRef]
- Gagnon, K.; Bocoum, E.H.; Chen, C.Y.; Baden, S.P.; Moksnes, P.O.; Infantes, E. Rapdi faunal colonization and recovery of biodiversity and functional diversity following eelgrass restoration. Restor. Ecol. 2023, 21, e13887. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).