Abstract
With the rapid development of nanotechnology, nanomaterials have been widely utilized in many industries and daily life applications due to their unique properties. However, their potential release and the human health/environmental consequences have raised public concern greatly. In this study, we compared the toxic effects of AgNPs and AgNO3 on Skeletonema costatum in 10, 100, and 500 μg·L−1 Ag treatments. In all the AgNP exposure experiments, cell membrane damage and growth inhibition occurred. However, the cellular damage only obviously appears on exposure to a high concentration of AgNO3. The antioxidant enzyme (SOD and CAT) activities and lipid peroxidation in Skeletonema costatum were also induced significantly in the AgNP treatments. In addition, the percentage of Ag release in seawater increased with the increase in AgNP concentrations (13%, 32% for 100 and 500 μg·L−1 AgNPs). Thus, the biotoxic effects of AgNPs were found to be due to a combination of the solubilization of particles into toxic metal ions and the nature of the nanoparticles. It was worth noting that the induction of oxidative stress and damage to the cell membrane comprised the dominant mechanism of toxicity for AgNPs. Therefore, the behavior of nanometals in seawater affects the biotoxic effect on the phytoplankton. These results shed light on the biological toxicity of nanometals and their possible toxicity mechanism.
1. Introduction
In recent years, silver nanoparticles (AgNPs) have received widespread attention [1,2,3]. Due to their stable physical and chemical properties, AgNPs show a better performance than silver in many aspects, and are widely used in textiles, food, water treatment, cosmetics, and other products [4,5]. In nature, many environmental factors affect the formation, aggregation, and dispersion of nanoparticles [6]. The colloidal organic matter is an important regulatory factor for the speciation of trace metals in seawater and even their biogeochemical cycling processes. These colloidal organic compounds can form nanoparticles by joining complexes with metals, thereby affecting the biogeochemical cycle of metals [2,7,8]. Dissolved organic matter in the aquatic environment can adsorb on the surface of nanometals and inhibit the aggregation of nanoparticles. The presence of organic matter affects the bioavailability and toxicity of nanometals [6,9,10], while ions in the water mass compress suspended nanometals and cause them to destabilize and precipitate [11]. Studies [12,13] have shown that a low concentration of humus can stabilize silver nanoparticles, while a high concentration can induce aggregation, thereby changing the bioavailability and biotoxicity of nanoparticles. For AgNPs, the ion release from the nanoparticle is significant (~34% of silver ions resolved from the nanoparticles), resulting in the absorption of metals by organisms exposed to AgNPs in both nano and ionic forms.
After entering the aquatic environment, nanoparticles usually aggregate under physical, chemical, biological, and other conditions, which may cause their aggregates to settle onto the surface of sediment, thereby posing an exposure toxicity risk to organisms living in benthic environments [14,15]. In addition, some active metal nanoparticles may also release metal ions into the water, posing an exposure toxicity risk to plankton that live in the aquatic environment. The physical and chemical properties of nanoparticles (including their size, shape, specific surface area, composition, aggregation, and dissolution) can affect their biotoxicity, especially some nanomaterials containing toxic metals. However, there is controversy as to whether “the biotoxicity is caused by the nanometal itself or the free ions released”. It is generally believed that the biotoxic effect constitutes oxidative stress induced in organisms [16,17,18,19,20]. However, the toxicological mechanism studies in nanometals to date have ignored metal absorption and accumulation in organisms. Ringwood et al. [21] found that low concentrations of AgNPs (<0.16 μg·L−1) did not affect the normal hatching of fertilized eggs in oysters. However, when the exposure concentration exceeded 0.16 μg/L, the content of metallothionein (MT) in the fertilized eggs of oysters increased significantly and reached 80 times the MT contents in the control groups. Thus, it can be seen that the effects of AgNPs on different biological levels may be different [22]. Molecular markers are, undoubtedly, more sensitive indicators. Therefore, it is necessary to study the biological toxic effects of nanometals using different biomarkers in combination with their accumulation in organisms.
Coastal waters are usually considered as the ultimate sink for pollutants, including nanomaterials. In the process of human activities, unintentional and intentional discharge can lead to a large amount of wastewater entering the coastal environment [5,23,24]. Similarly, widely used nanomaterials will inevitably enter the marine environment and cause harm to the marine ecosystem [25,26]. Marine microalgae are highly abundant in coastal waters, so microalgae growth is likely to be affected by anthropogenic pollution. Skeletonema costatum (S. costatum) is a nearshore diatom which is distributed in waters with a wide range of temperatures and salinity levels. Among more than 40 species of bloom algae recorded in Jiaozhou Bay, a region of high urbanization in China, S. costatum is one of the dominant species [27]. As a typical bloom alga with low levels in the food chain, the absorption of nanoparticles in S. costatum directlys affect material transfer throughout the food chain. Therefore, the biotoxic effects of AgNPs on S. costatum may affect the entire coastal ecosystem. Thus, we selected S. costatum as the experimental object that has a certain ecological significance.
As it is an emerging pollutant, the widespread application of AgNPs may cause pollution to the water mass and also induce adverse effects on human health. However, it is difficult to evaluate the ecotoxicity risk of nanomaterials because their toxicity mechanism is still unclear. Understanding the biotoxicity mechanism of AgNPs on marine algae can help us to predict the impact of nanomaterials on the food web and the entire ecosystem in coastal waters. It is unclear how the particle size, shape, and surface properties of NPs are altered after entering the seawater, and how these variations could affect their biotoxicity to marine organisms. Therefore, it is necessary to conduct deeper research to explore the biotoxicity mechanism of AgNPs on marine organisms. Studying the toxic effect of AgNPs on coastal algae is of great scientific value in the protection of coastal ecosystems and provides a scientific basis for further revealing the biotoxic mechanism of nanoparticles.
2. Methods and Materials
2.1. Characteristics of AgNPs
AgNPs (<100 nm, 99.5% trace metal basis) were purchased from Sigma-Aldrich (https://www.sigmaaldrich.cn/CN/zh, accessed on 3 April 2014). The standard solution of Ag (AgNO3) was purchased from China Standard Substance Network (https://www.reagent.com.cn/, accessed on 12 March 2014).
AgNP solutions were prepared with filtered seawater and Milli-Q water, respectively. The concentrations of 10, 100, and 500 μg·L−1 AgNPs/AgNO3 were adapted in this study. The 500 μg·L−1 AgNPs in seawater solution were observed using a scanning electron microscope (SEM, QUANTA200, FEI Company, Hillsboro, OR, USA).
After 24 h of solution preparation in seawater, the AgNP solution was subjected to ultrafiltration using the 3 KD Millipore ultrafiltration centrifuge tube (UFC500396). The Ag+ ion in filtrate, released from the AgNPs, was measured in acidic conditions using a flame atomic absorption spectroscope (MD2134, Thermo Fisher Scientific, Waltham, MA, USA).
2.2. Determination of Cytotoxic Effects
The f/2 medium was prepared with reference to the method in the study by Guillard et al. [28]. To avoid the complexation of Ag ions, Na2EDTA was not added to the culture medium. Thus, it is called “modified f/2 medium”.
The cytotoxic experiments were performed in three groups (control, AgNP exposure, and AgNO3 exposure). In a previous study [29], the concentration of AgNPs in industrial discharge water was found to reach 1–6 mg·L−1. Considering the dilution effect of seawater, we set up 10, 100, and 500 μg·L−1 AgNPs in a seawater culture medium. The AgNO3 exposure experiment also used the concentrations of 10, 100, and 500 μg·L−1 in the culture medium for comparison. Every Ag concentration gradient was performed in triplicate. The alga was cultivated in a constant-temperature incubator and manually shaken three times a day.
Cysteine can be used as a complexing agent to eliminate the role of Ag ions in AgNP toxicity. In our study, cysteine was added to the medium to complex Ag+. In the 500 μg·L−1 AgNO3 treatments, cysteine was added to the medium at a concentration of 500 μg·L−1. In order to avoid the differences in the impact of cysteine, equivalent cysteine also be added to the 500 μg·L−1 AgNP treatment. The Ag-cysteine formed could decrease the toxic effect induced by the Ag+ released from the nanoparticles.
2.2.1. The Relative Growth Rates
The density of algal cells was measured using the microscope at the exposure times of 0, 24, 48, 72, and 96 h, and the relative growth rate was calculated as follows.
where N1, N0 are the density of the algal cell at time T1 and T0, respectively.
Relative growth rate = (ln N1 − ln N0)/(T1 − T0)
2.2.2. Ag Accumulation in the Algal Cell
After 48 h of exposure, 100 mL of algal culture was centrifuged into algal mud and digested with HNO3 and H2SO4 using the microwave digestion method. Metal ligands, like Na2EDTA and cysteine were not added, to avoid the complexation of Ag. The silver content in the algal cell was detected in the digestion solution via flame atomic spectrophotometry.
2.2.3. The Damage to the Cell Membrane
According to Tommaso Pisani’s method [30], the electrolyte flow out after the destruction of the algal cell membrane causes changes in the conductivity of the algal solution. The conductivity E1 and E2 of the algal solution before and after boiling were measured. The degree of the damage to the cell membrane was expressed as E1/E2. The larger the value, the greater the degree of damage.
2.2.4. Determination of the Chl-a
Referring to the method of Porra et al. [31], the absorbance of the supernatant at the wavelengths of 665 nm and 652 nm was measured using a visible spectrophotometer. The Chl-a was calculated as follows.
where A665 and A652 represent the absorbance of the supernatant at the wavelengths of 665 nm and 652 nm.
Chlorophyll a (mg·L−1) = 16.29 × A665 − 8.54 × A652
2.3. Determination of Antioxidant Stress
2.3.1. Preparation of Protease Extract
Firstly, 20 mL algal solution was sampled to centrifuge at 5000 r/min for 10 min. The supernatant was removed. The precipitate was added to an appropriate amount of 0.05 mol·L−1 phosphate buffer (pH = 7.0), then crushed ultrasonically. The liquid was centrifuged at 4 °C, 6500 r/min for 20 min, and the supernatant was the protease extract.
The preparation of the protease extract and the subsequent determination of the protein content, SOD, CAT, and MDA were performed with reference to the instructions in the biological reagent kit purchased from Nanjing Jiancheng Biological Company (http://www.njjcbio.com/, accessed on 15 May 2014).
2.3.2. Determination of Protein Content, SOD, CAT, and MDA
The protein content in the protease extract was determined via the Coomassie brilliant blue method, and the absorbance of the protease extract was measured at the wavelength of 595 nm. The protein content was calculated as follows:
where ODm and ODs are the optical density of the tested sample and standard sample, respectively. ODb is the optical density of the blank samples. Cprot. is the content of the protein, and 0.563 g/L is the protein content of the standard sample.
The SOD content in the protease extract was determined via the hydroxylamine method, and the absorbance of the protease extract was measured at the wavelength of 550 nm. The SOD content was calculated as follows:
where ODc and ODm are the optical density of the control and measured sample, respectively. Vt and Vm are the volume of the total sample and measured sample, respectively, in the unit of mL. The protein content is in the unit of mg prot/mL. The SOD content is in the unit of U/mg prot, representing the SOD unit per mg protein.
The CAT content in the protease extract was determined via the visible light photometric method, and the absorbance of the protease extract was measured at the wavelength of 405 nm. The CAT content was calculated as follows:
where ODc and ODm are the optical density of the control and measured sample, respectively. Vm is the volume of the measured sample, in the unit of mL. The protein content is in the unit of mg prot/mL. The CAT content is in the unit of U/mg prot, representing the CAT unit per mg protein.
The MDA content in the protease extract was determined via the spectrophotometry method, and the absorbance of the protease extract was measured at the wavelength of 532 nm. The MDA content was calculated as follows:
where ODm, ODc, ODs, and ODbl are the optical density of the measured, control, standard, and blank sample, respectively. Con.s is the concentration of MDA in the standard sample (10 nmol/mL). The protein content is in the unit of mg prot/mL. The MDA content is in the unit of U/mg prot, representing the MDA unit per mg protein.
2.4. Statistical Analyses
The data are expressed as mean ± SD. The difference analysis between the control and the exposure experiments is conducted using XLstat and one-way ANOVA. The significant difference is p < 0.05.
3. Results and Discussion
3.1. Characteristics of AgNPs
In an aquatic environment, the physical and chemical conditions could affect the agglomerate and deposition of nanoparticles, thereby altering their toxic effects on organisms. Therefore, it is necessary to consider the impact of the possible flocculation of nanoparticles on their toxicity mechanism.
Although the PVP coated on the surface of AgNPs plays a role in dispersion and stability, the SEM image (Figure 1) shows that AgNPs flocculated to an irregular granular shape in filtered seawater at different magnifications, and particles are produced with a size of micron level. Due to the limitation of the accuracy of the device, the precision of the images in Figure 1 is 10 and 5 μm, respectively. It can be seen from the SEM results that the AgNPs obviously flocculated in seawater, and the size mostly varied between 100 nm and 10 μm.

Figure 1.
SEM images of AgNPs (500 μg·L−1, Sigma, <100 nm) dispersed in seawater.
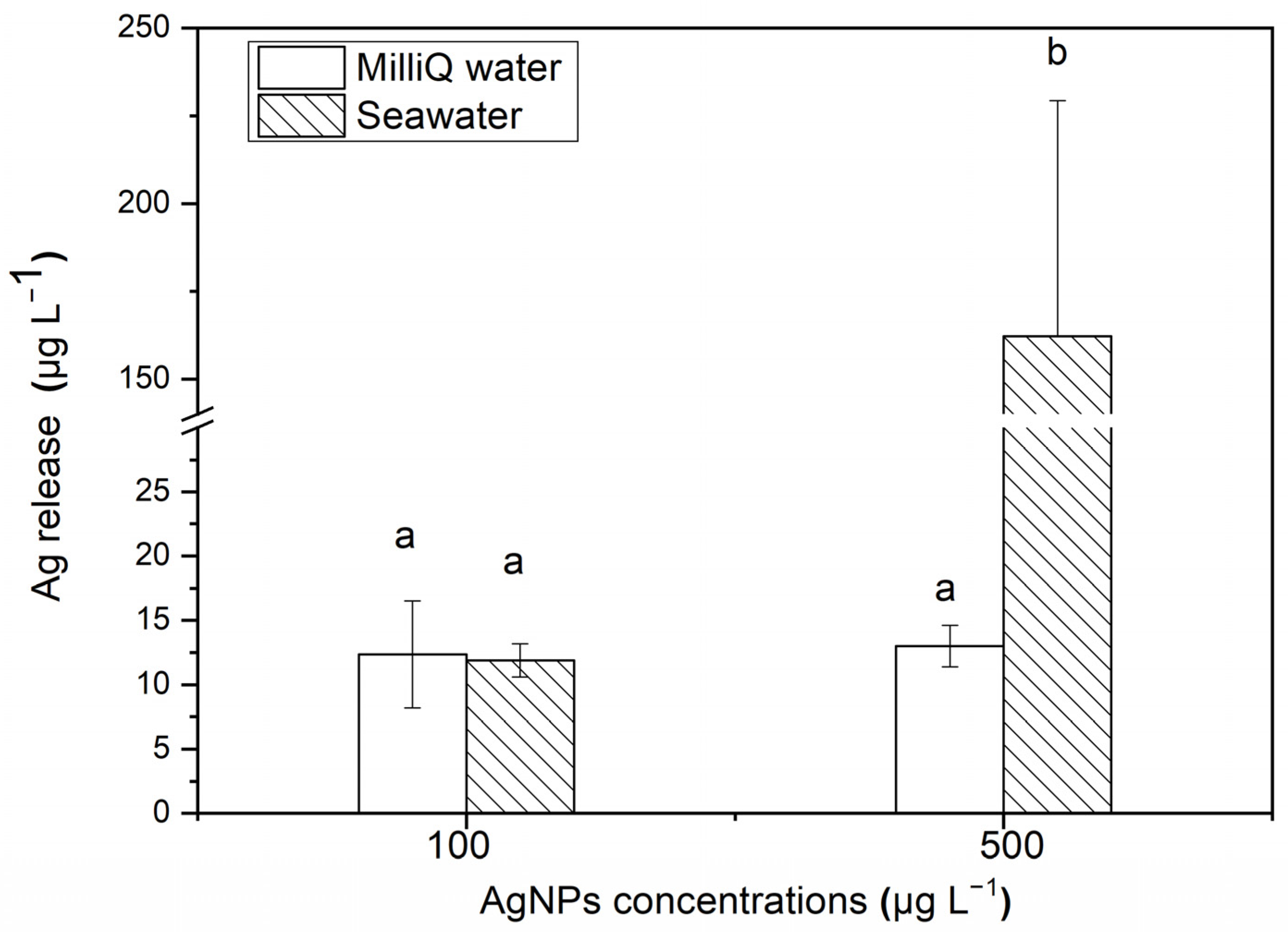
After 24 h, the release of soluble Ag from Milli-Q water and seawater with different concentrations of AgNPs is shown in Figure 2. For 500 μg·L−1 AgNP solutions, ~32.43% Ag is released and exists in the filtered seawater medium as the soluble state (162.17 ± 67.14 μg·L−1), which is significantly higher than the release amount in Milli-Q water medium (11.9 ± 1.29 μg·L−1). However, for 100 μg·L−1 AgNP solutions, there is no significant difference in the released soluble Ag contents between the filtered seawater medium (13.01 ± 3.61 μg·L−1) and Milli-Q water medium (12.35 ± 4.17 μg·L−1). After 48 h, the content of soluble Ag released in both the filtered seawater and Milli-Q water medium were lower than the detection limit of AAS, indicating that the aggregation of AgNPs inhibited the release of soluble Ag from the nanoparticles.
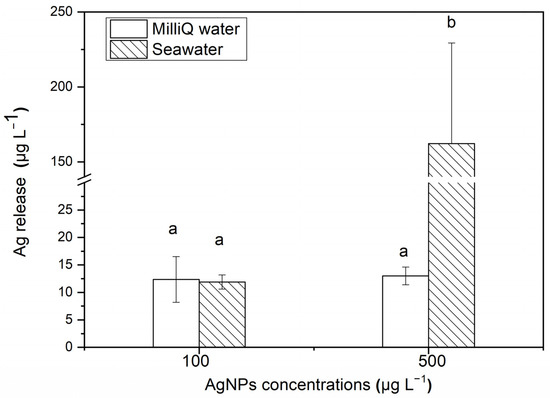
Figure 2.
After 24 h, the release of soluble Ag from AgNPs in Milli-Q water and seawater. The same identification letter (e.g., a and a) represents no significant difference, while different identification letters (e.g., a and b) represent a significant difference.
Many studies have proven that AgNPs easily flocculate in seawater. Gomes et al. [32] showed that AgNPs with the initial particle size of 41.7 ± 9.6 nm flocculated to 144.2 ± 39.2 nm in a seawater suspension. Buffet et al. [13] demonstrated that the particle size of AgNPs would increase from 40 nm to 150–500 nm. The reasons for the change in particle size are summarized as the presence of inorganic ligands in seawater or the large specific surface area of nanoparticles, which may also react with organisms. Therefore, the environmental behavior of AgNPs and the reaction between AgNPs and biological cells are closely related [33,34].
The high ionic strength of seawater has a great influence on the aggregation and dissolution of nanoparticles. In this study, the percentage of released soluble Ag decreased with an increasing AgNP concentration in Milli-Q water (12% and 2% Ag released in 100 and 500 μg·L−1 AgNP solutions, respectively), while the results were the opposite in seawater (13% and 32%, respectively). In comparison, in the study by Zou et al. [35], 18% Ag was released from a 1500 μg·L−1 AgNP solution, which is higher than our results. The AgNPs in the study by Zou et al. [35] were synthesized by AgNO3 and NaBH4 in the presence of ATP-2Na. The research by Burchardt et al. [36] on the impact of AgNPs on diatoms showed that between 68% and 87% Ag was released from a 10 μg·L−1 AgNP solution in artificial seawater, and between 2% and 3% Ag was released in freshwater (BG11 medium). The content of Ag released in seawater was much higher than that in freshwater, which is the same as that in our study. In the study of ZnO NPs by Wang et al. [37], it was found that more than 50% Zn was released in the form of a dissolved state in 102 to 1700 μg·L−1 ZnO NPs, which is higher than the release of Ag from the AgNPs in our study. This showed that the release of ions from the nanoparticles—that is, the stability of the nanoparticle solution—would be affected by the surface wrapping, the synthetic method, the environmental media, the metal type and the environmental conditions (the organic content, pH, etc.). Therefore, understanding the possible morphological changes and dissolution in nanoparticles in different environments is of great significance in studying the toxic effects of nanoparticles on organisms [38,39].
3.2. Biological Effects of AgNPs on S. costatum
3.2.1. The Relative Growth Rate
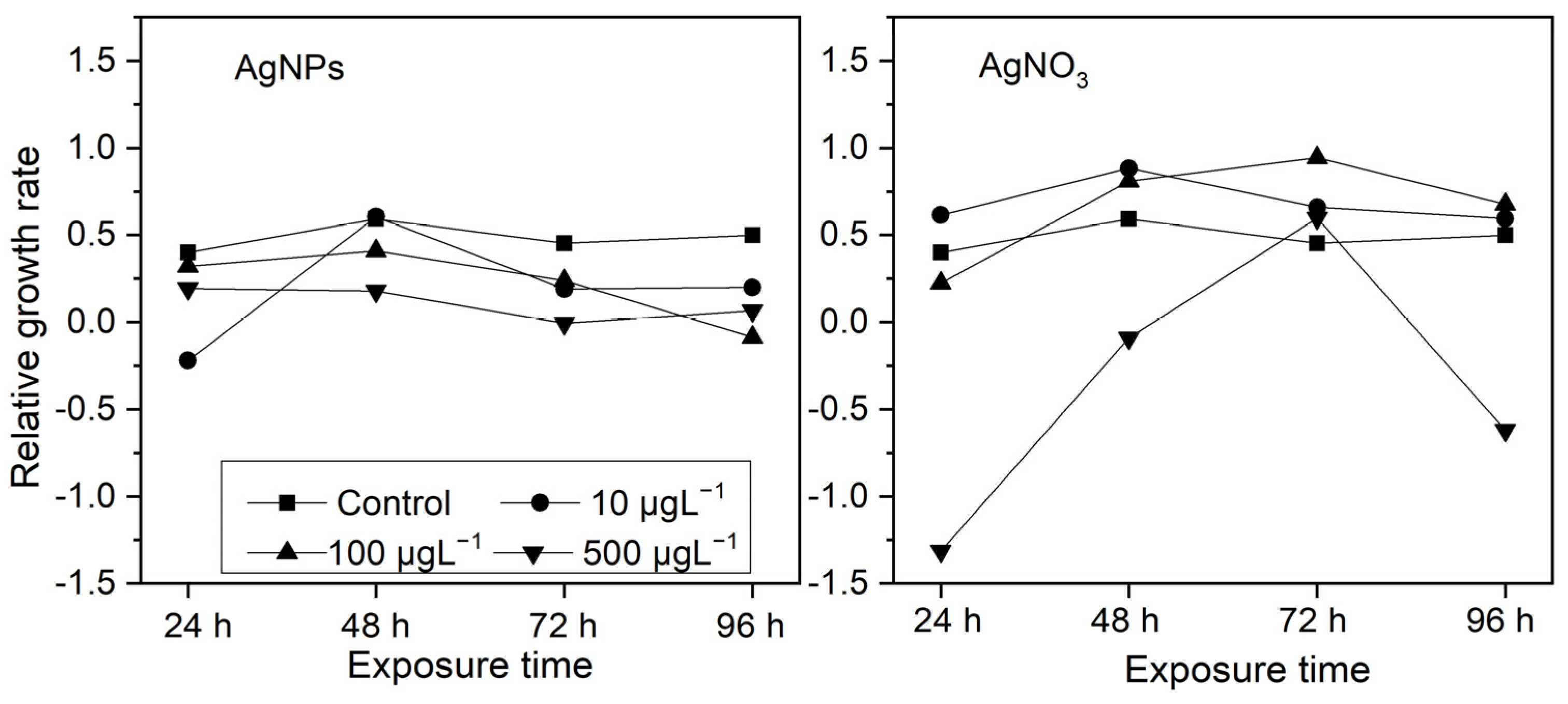
From Figure 3, it can be seen that, after being exposed to different concentrations of AgNPs for 24 h, most of algae exhibited varying degrees of growth inhibition and, as the AgNP concentrations increased, the growth inhibition became apparent, except at 10 μg·L−1AgNP exposure, in which mild growth promotion occurred. In terms of ionic Ag exposure, S. costatum showed significant growth inhibition only at a high concentration (500 μg·L−1). Interestingly, at exposures with a low-concentration ionic Ag level (10 and 100 μg·L−1), the growth in S. costatum was promoted to a certain extent. This may be related to the “low toxicity stimulation” effect.
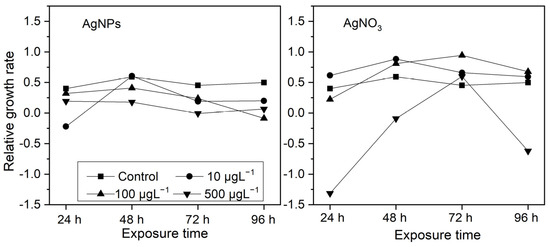
Figure 3.
The effects of different concentrations of AgNPs and soluble Ag concentrations (10, 100, and 500 μg·L−1) on the relative growth rate of S. costatum at 24, 48, 72, and 96 h.
After the initial 24 h, the growth inhibition was more obvious in the 10 μg·L−1 AgNP treatment than in the 100 and 500 μg·L−1 AgNP treatments, while the growth inhibition of algae was more pronounced in the 500 μg·L−1 ionic Ag treatment than in the 10 and 100 μg·L−1 ionic Ag treatments. This may be due to the fact that nanoparticles with a higher concentration are more likely to form aggregates in an algal solution, which temporarily delays the growth inhibition of S. costatum, while a high concentration of ionic Ag has an immediate inhibitory effect on algal growth.
3.2.2. Ag Accumulation
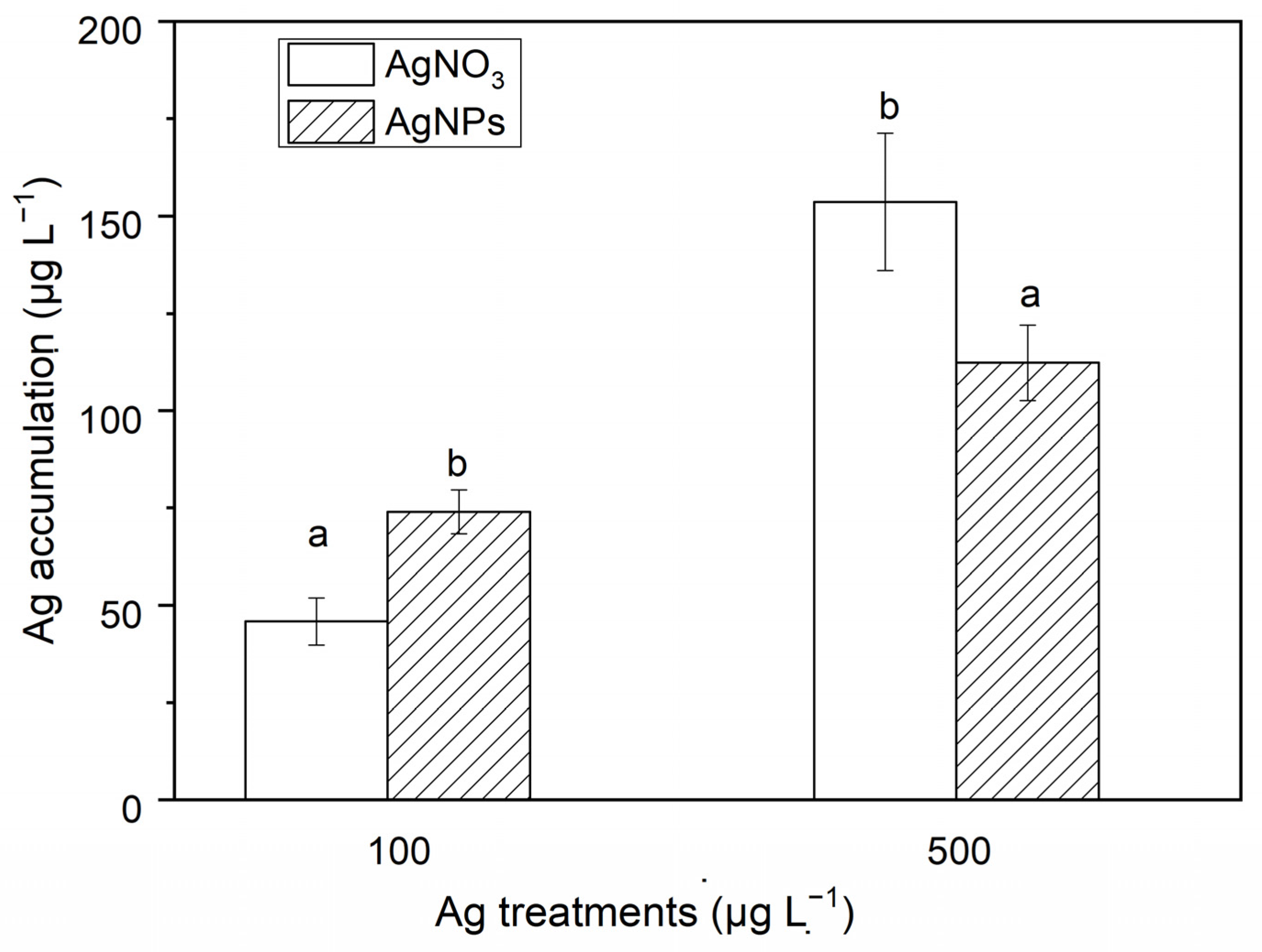
The Ag accumulation in S. costatum at different exposures of AgNPs and ionic Ag is shown in Figure 4. As seen in Figure 4, the Ag accumulation in S. costatum at 48 h of exposure to ionic Ag (500 μg·L−1) was 3.70 times higher than that in the AgNP solution (500 μg·L−1). However, the Ag accumulation for the exposure with 100 μg·L−1 AgNPs was 1.64 times higher than that in the algae exposed to the ionic Ag solution. In addition, a higher concentration of Ag exposure induced a greater Ag accumulation. For example, the Ag accumulation in the algae exposed to 500 μg·L−1 AgNPs was 1.52 times that in the algae exposed to 100 μg·L−1 AgNPs. The Ag accumulation in the 500 μg·L−1 ionic Ag exposure was 3.35 times that in the 100 μg·L−1 ionic Ag exposure experiments. The Ag accumulation levels in the control and the 10 μg·L−1 Ag exposure experiments were all lower than the detection limit. The above results show that AgNPs can be absorbed in the form of particles. High concentrations of ionic Ag and low concentrations of AgNPs are more likely to accumulate in cells, which may be related to the aggregation of nanoparticles. There are more opportunities for high concentrations of AgNPs to form larger-particle-size aggregates through collisions between particles. The particle size of the aggregates exceeds the pore size of the cell membrane, hindering the entry of nanoparticles into the cell.
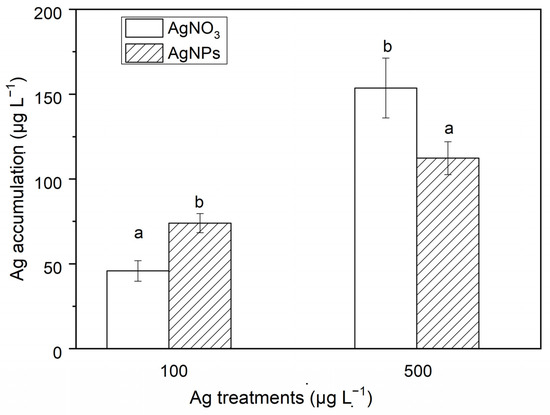
Figure 4.
The Ag accumulation in algae exposed to soluble Ag and AgNPs for 48 h. The same identification letter (e.g., a and a) represents no significant difference, while different identification letters (e.g., a and b) represent a significant difference.
3.2.3. Destruction of Membrane Stability of S. costatum by AgNPs
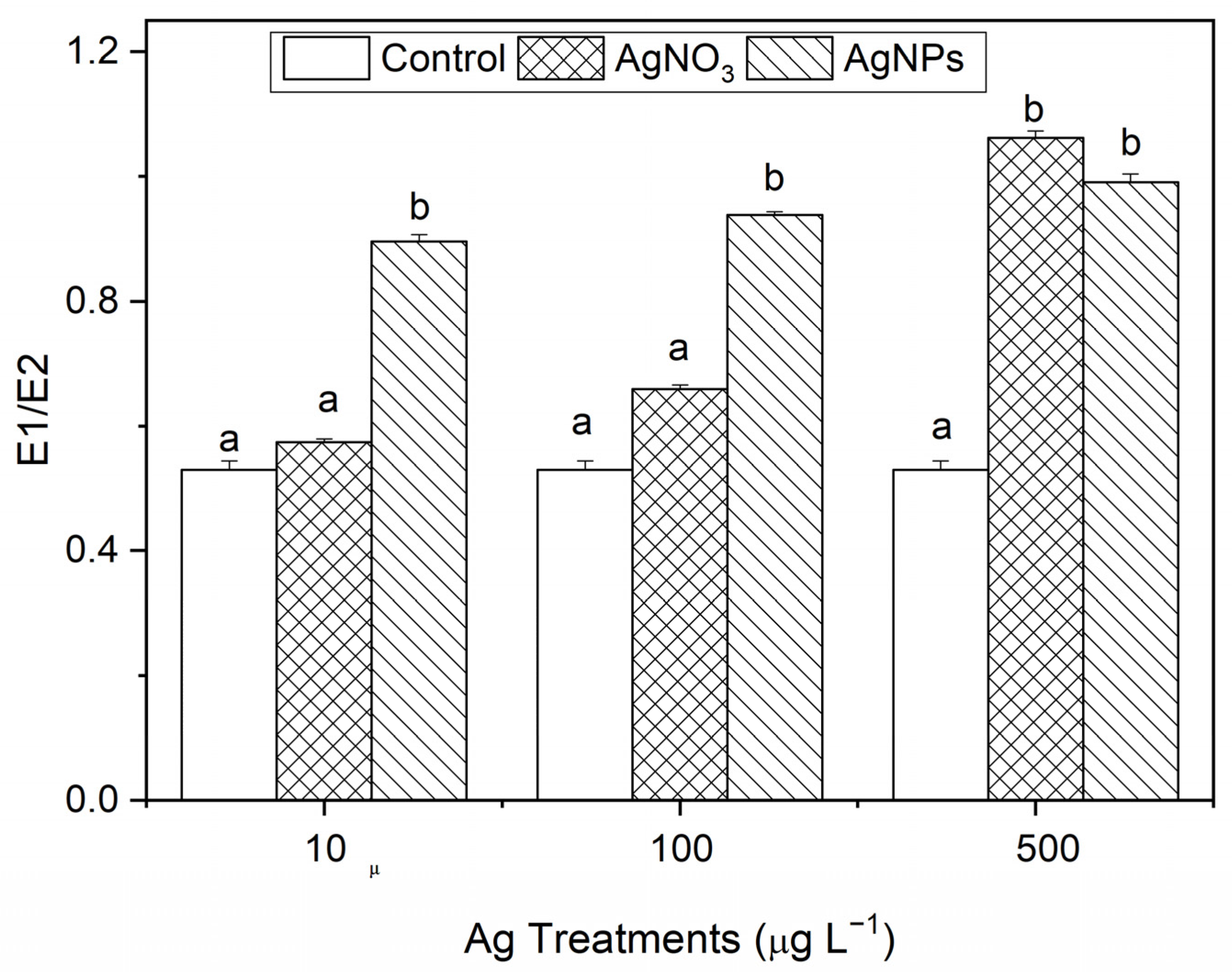
The degree of damage of the cell membrane is represented in E1/E2. The larger the value, the greater the degree of damage. From Figure 5, it can be seen that, compared with the control, the AgNP exposure showed obvious damage to the algal cell membrane. With the ionic Ag exposure, only the 500 μg·L−1 levels showed obvious damage to the algal cell membrane. The results show that AgNPs can induce damage to the stability of algal cell membranes at a relatively low concentration level, which is related to the unique properties of nanoparticles.
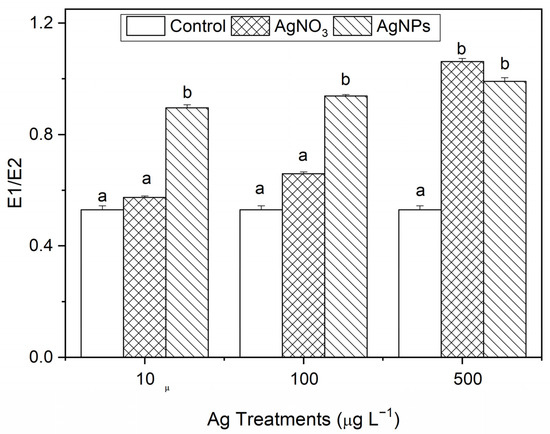
Figure 5.
The stability changes in the cell membrane after S. costatum was exposed to different concentrations of soluble Ag and AgNPs for 48 h. The same identification letter (e.g., a and a) represents no significant difference, while different identification letters (e.g., a and b) represent a significant difference.
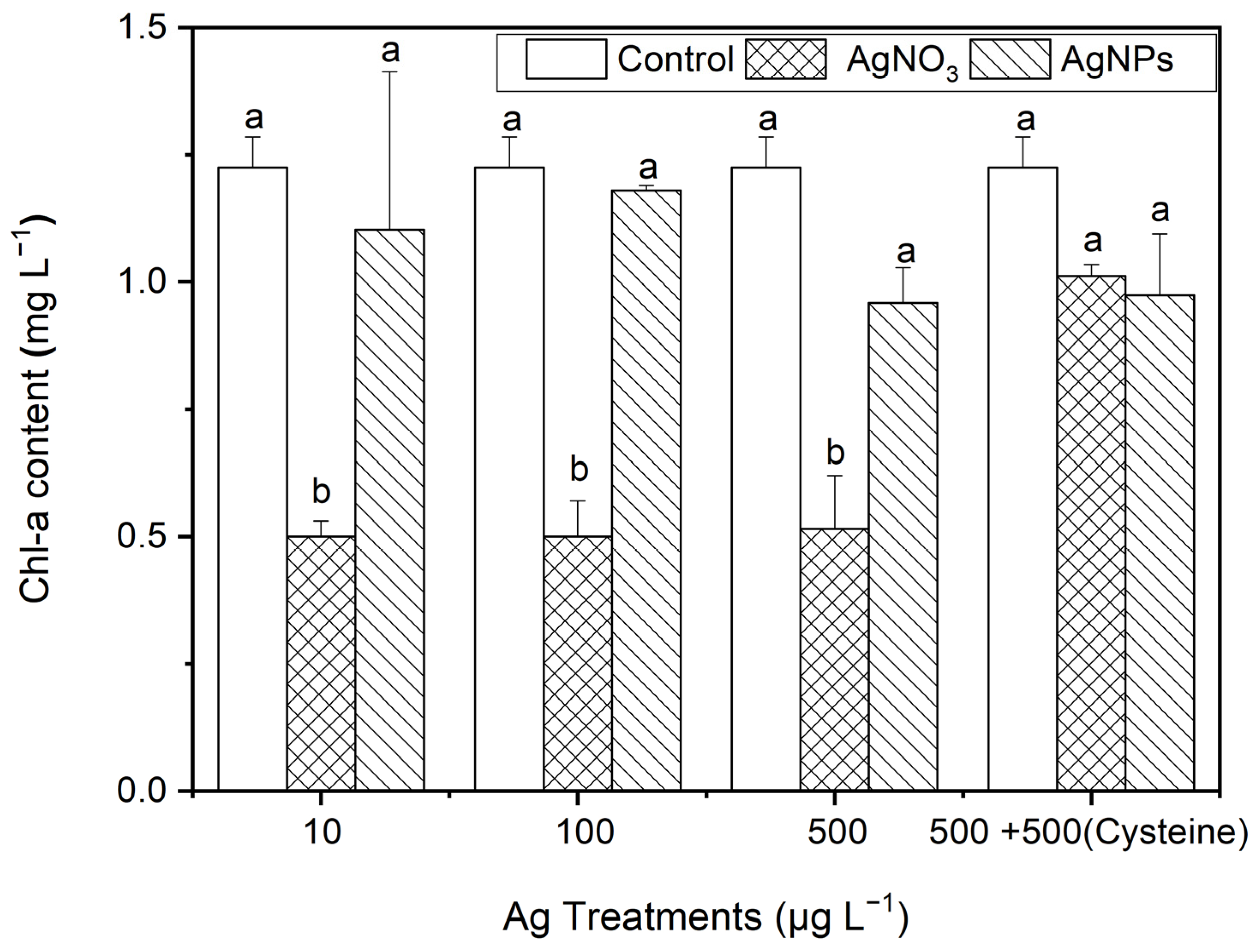
3.2.4. The Effect of AgNPs on Chl-a Contents in S. costatum
After 48 h of exposure to Ag solutions, the Chl-a contents in S. costatum under the AgNP treatments decreased slightly (from 4% to 22%) compared to the control (p > 0.05, Figure 6). Regarding the ionic Ag exposure, the Chl-a contents decreased significantly, by about 60%. This suggests that the ionic Ag was the main cause of the decrease in Chl-a in S. costatum. It is worth noting that, after the addition of cysteine, the Chl-a contents in S. costatum under the AgNO3 treatments showed no significant difference from the control and AgNP exposures. This indicates that cysteine eliminates the inhibitory effect on the Chl-a production from complexing ionic Ag. Furthermore, it could be speculated that the inhibitory effect on Chl-a production was mainly caused by Ag ions, but the soluble Ag released in the AgNPs (100 to 500 μg·L−1) was not enough to induce an obvious inhibitory effect on the Chl-a production in S. costatum.
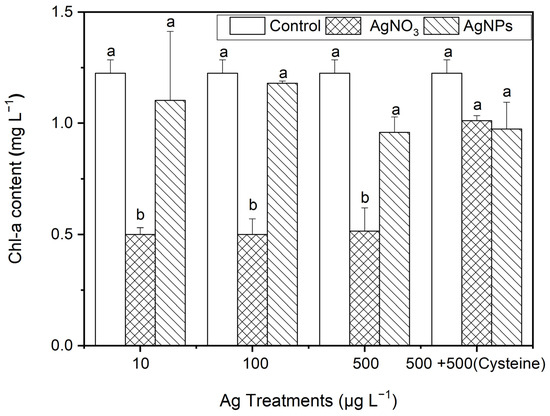
Figure 6.
The evolution of the Chl-a content in algae exposed to different concentrations of ionic Ag and AgNPs for 48 h. The same identification letter (e.g., a and a) represents no significant difference, while different identification letters (e.g., a and b) represent a significant difference.
The adsorption of AgNPs on the surface of microalgae cells will induce obvious toxicity through direct or indirect physical reactions, such as the disintegration of cell walls [40], or the reduction in light required for algal growth (“shading effect”) [41], and limit the utilization of nutrients [42,43]. In the study by Burchardt et al. [36], the same concentration of AgNPs also exhibited varying degrees of growth inhibition on Pseudomonas aeruginosa (inhibition percentage: from 40% to 60%) and Polycystis cyanobacteria (inhibition percentage: 100%). In this study, the growth inhibition percentage of S. costatum was 58%, 70%, and 86% for 10, 100, and 500 μg·L−1 AgNP exposure, respectively. Due to the limited research on the toxicity of AgNPs to algae, it can be inferred that the sensitivity of S. costatum to AgNPs is approximately between those of Pseudomonas aeruginosa and Polycystis cyanobacteria. In addition, low concentrations of AgNPs have a more obvious inhibitory effect on algal growth than relative high concentrations of AgNPs, while a high concentration of ionic Ag has an obvious algal inhibitory effect. This may be due to the fact that relative high concentrations of nanoparticles are more likely to form flocs in the algal medium, which temporarily delays the growth inhibition of S. costatum. The effect of ionic Ag on algal growth can be explained by the “Hormesis effect of toxins”, which can be said to be an adaptive response of organisms to toxins (in the sense of, “What does not kill you makes you stronger”). That is to say, exposure to low-dose toxins will appropriately promote a certain function in the organism (referring to algal growth in this case), while exposure to high concentrations of toxins will have an inhibitory effect on that function.
The accumulation of metals in the organisms indicates that the toxicity of nanometals to organisms is, to some extent, based on the absorption of metals by organisms. Perreault et al. [44] emphasized, in their study on the toxicity of nano copper oxide to the green alga Chlamydomonas reinhardtii, that the exposure to polymer encapsulated nanoparticle solution could increase the absorption of nano copper oxide by algal cells. Hull et al. [45] studied the absorption and retention of nanometals by purple mussels, indicating that, after initial exposure for 8 h, more than 90% of nanometals were absorbed by purple mussels, and almost all nanometals were present in the digestive glands of mussels. In this study, after 48 h of exposure to 500 μg·L−1 AgNPs, approximately 22% of Ag was accumulated in the S. costatum. Therefore, the biological absorption of nanoparticles cannot be ignored when studying the biological toxicity effects of nanoparticles.
As is well known, the cell wall is the first reaction site between nanoparticles and cells, and serves as an important barrier to the absorption of nanoparticles. According to the relevant research [46,47,48], the pore size on the cell wall of diatoms is, approximately, from 3 to 50 nm. That is to say, if the particle size of the nanoparticles is smaller than the pore size, then they can enter the cell through endocytosis, diffusion, or reacting with carrier proteins [47,48]. Moreover, during the regeneration process, the permeability of the cell wall changes. The newly generated cell wall can make it easier for nanoparticles to enter [49]. In addition, the reaction between cells and nanoparticles induces a new pore size that is larger than the normal pore size, which increases the possibility of nanoparticles entering the cell through the cell wall [43,47].
As one of the important producers in the entire marine ecosystem, diatom photosynthesis plays a crucial role in energy flow and nutrient transfer in the food web. Therefore, studying the impact of nanoparticles on alga photosynthesis has a certain scientific significance. Oukarroum et al. [50] studied the inhibitory effect of AgNPs on freshwater algae (Chlorella vulgaris) and marine algae (Dunaliella salina), and the results showed that AgNPs caused a sharp decrease in chlorophyll production in these two algae. Chen et al. [51] studied the effect of nano titanium dioxide on the photosynthetic capacity (Fv/Fm) of Chlamydomonas reinhardtii. The results showed that, when the concentration of nano titanium dioxide was >1 mg·L−1, the photosynthetic capacity of Chlamydomonas reinhardtii decreased rapidly. Perreault et al. [44] studied the toxicity of nano copper oxide particles to the green alga Chlamydomonas reinhardtii, and indicated that the polymer-encapsulated nanoparticle solutions significantly reduced the electron transfer ability of Chlamydomonas reinhardtii photosynthetic system II. Miao et al. [52] studied the toxicity of nano zinc oxide to Thalassiosira pseudonana; it was found that, as the concentration of nanoparticles increased, the photosynthetic capacity (Fv/Fm) almost decreased to 0, and the Chl-a content decreased by 81%. Röhder et al. [53] showed that, when the concentration of nano tin oxide increased to 50 μM, the short-term toxicity of nano tin oxide particles was obviously promoted, and the photosynthesis yield decreased significantly. Therefore, the inhibition of photosynthesis is an important cause of the toxicity of nanoparticles to the aforementioned alga. However, in this study, although the decreases in Chl-a were induced by different concentrations of AgNPs, there was no significant difference compared to the control. This indicates that the inhibition of photosynthesis by AgNPs (within the concentration range from 10 to 500 μg·L−1) was not the main cause of toxicity to S. costatum.
3.3. The Effect of AgNPs on the Antioxidant System of S. costatum
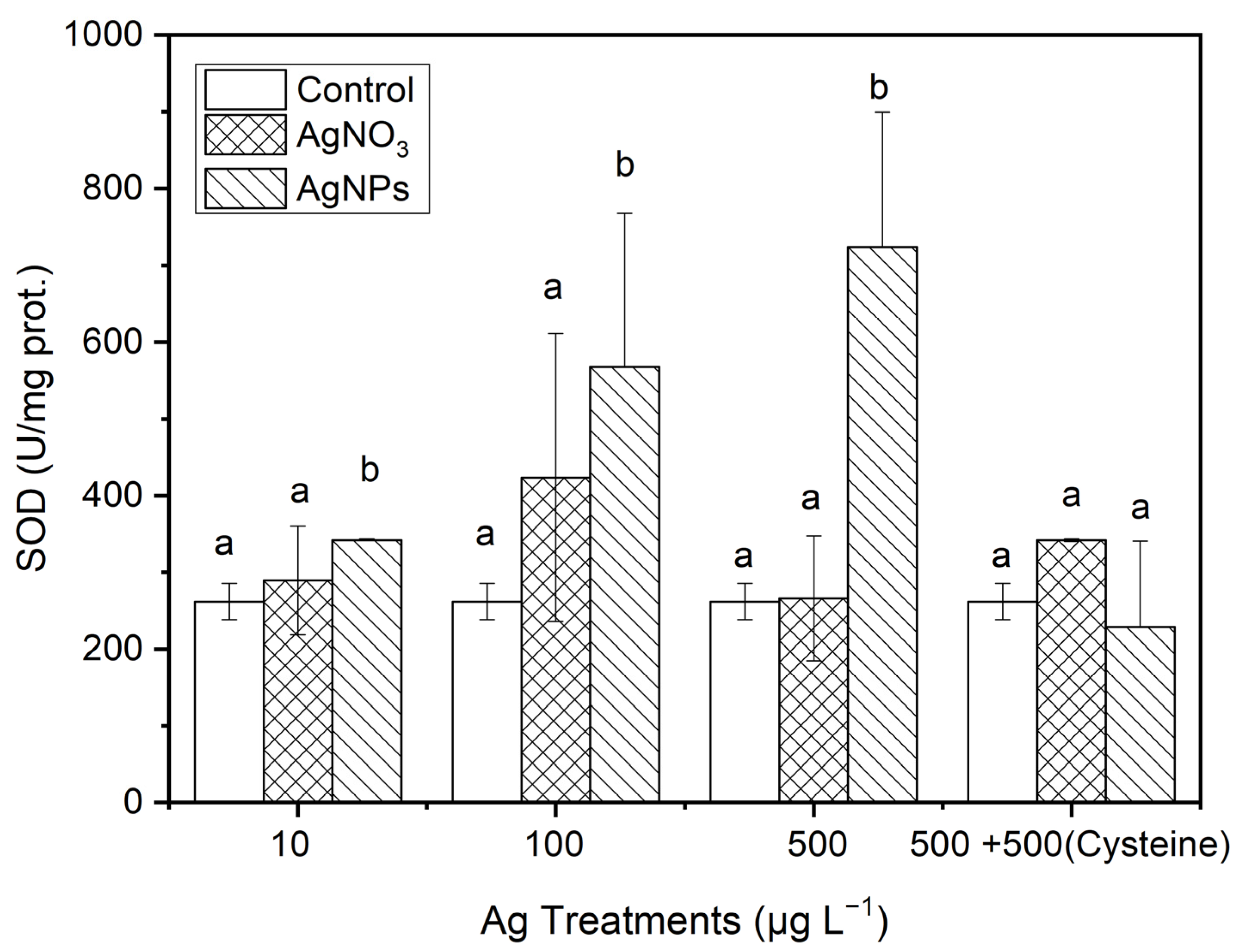
3.3.1. SOD
From Figure 7, it can be seen that, after 48 h of exposure to Ag solutions with different concentration gradients, the SOD contents in the algae in the AgNP treatments were 1.17, 0.34, and 1.72 times higher than those in the ionic Ag treatments for 10, 100, and 500 μg·L−1, respectively. However, the SOD contents in S. costatum in the equal amount of cysteine addition treatments, there was no significant difference between the control and the two forms of Ag treatment.
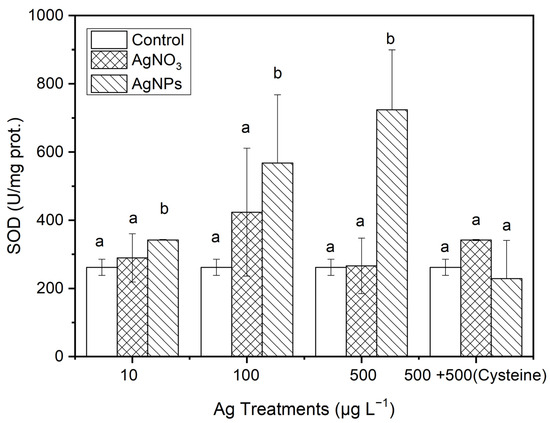
Figure 7.
The evolution of SOD content in algae exposed to different concentrations of soluble Ag and AgNPs for 48 h. The same identification letter (e.g., a and a) represents no significant difference, while different identification letters (e.g., a and b) represent a significant difference.
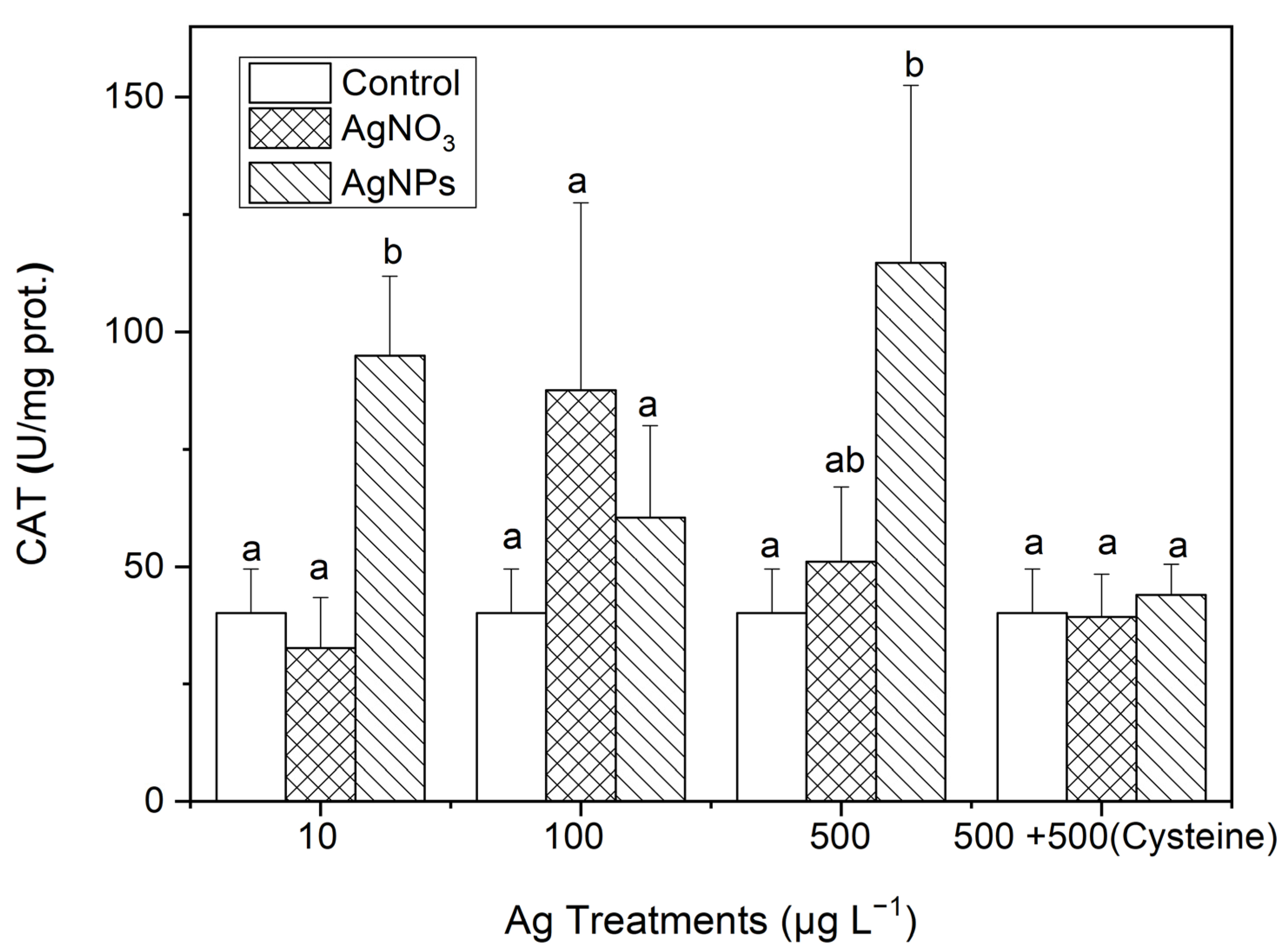
3.3.2. CAT
As can be seen in Figure 8, the CAT contents in S. costatum under AgNP exposure were 2.37, 1.51, and 2.86 times higher than those in the control for the 10, 100, and 500 μg·L−1 treatments, respectively. Additionally, there was a significant difference in the CAT contents only between the 10 μg·L−1 AgNP and ionic Ag treatments. After the addition of an equal amount of cysteine to the 500 μg·L−1 Ag solutions, there was no significant difference in the CAT contents between the control and the two forms of Ag treatments, which was similar to the changes in SOD contents in Section 3.3.1.
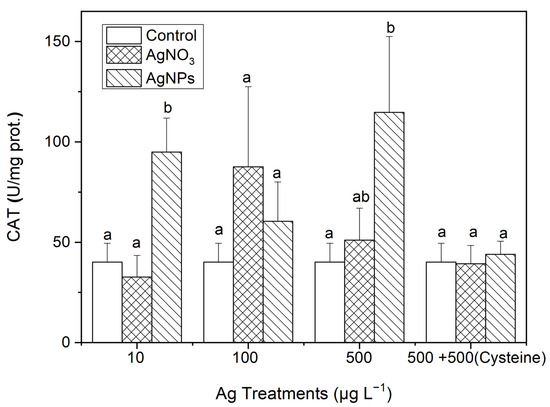
Figure 8.
The evolution of the CAT content in algae exposed to different concentrations of soluble Ag and AgNPs for 48 h. The same identification letter (e.g., a and a) represents no significant difference, while different identification letters (e.g., a and b) represent a significant difference.
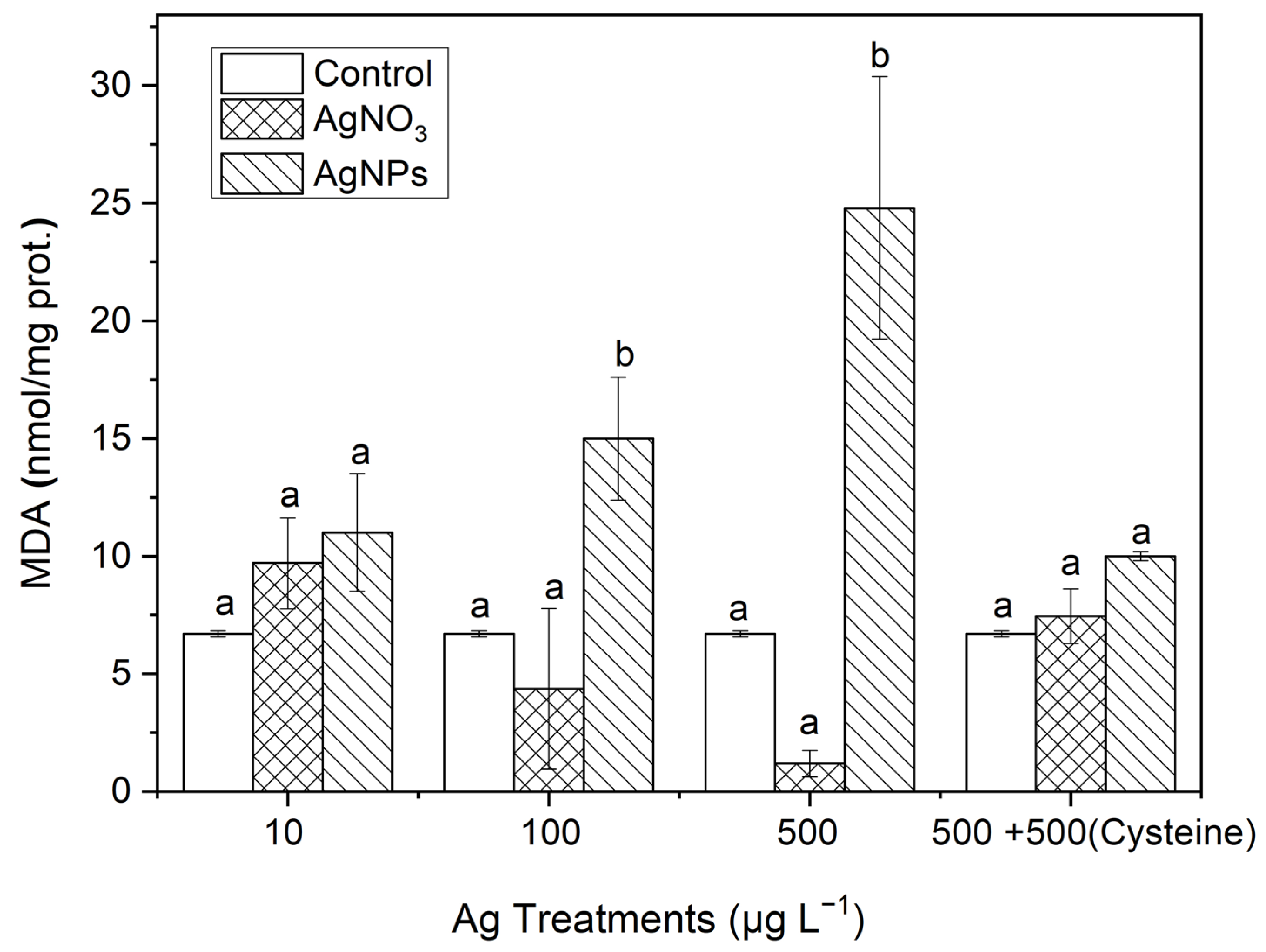
3.3.3. MDA
After organisms are subjected to oxidative stress, the production of MDA can effectively represent the degree of lipid peroxidation. From Figure 9, it can be seen that the MDA contents in S. costatum had also undergone varying degrees of change. In the 100 and 500 μg·L−1 AgNP treatments, the MDA contents were 3.42 and 20.67 times higher than those in the soluble Ag treatments, respectively. There was no significant difference in the MDA contents between the soluble Ag treatments and the control. After an equal amount of cysteine was added to the 500 μg·L−1 Ag solutions, no significant difference in MDA contents occurred between the control and the two forms of Ag treatments, which was similar to the changes in SOD and CAT contents. This indicates that AgNPs could induce an obvious oxidative stress on S. costatum.
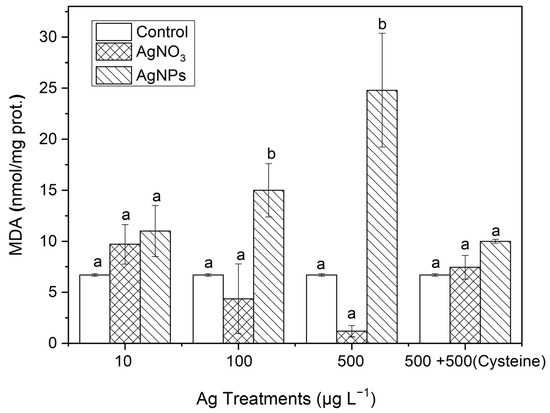
Figure 9.
The evolution of the MDA content in algae exposed to different concentrations of soluble Ag and AgNPs for 48 h. The same identification letter (e.g., a and a) represents no significant difference, while different identification letters (e.g., a and b) represent a significant difference.
The effect of AgNPs on oxidative stress and the antioxidant defense can alter the enzyme activity of SOD and CAT. The induction of these antioxidant enzymes is an effective detoxification mechanism for alga cells to resist exposure to nanoparticles. The production of lipid peroxide MDA indicates that the toxicity of AgNPs is related to oxidative stress. We speculate that, as the concentration of AgNPs continues to increase, the enzyme activity of SOD and CAT will decrease, and the MDA content will continue to increase, because the further increase in the degree of oxidative stress may exceed the ability of antioxidant enzymes to scavenge free radicals.
The study by Oukarroum et al. [50] showed that AgNPs induced lipid peroxidation in two green algae (Chlorella vulgaris and Dunaliella salina). Buffet et al. [54] studied the toxicity of AgNPs to two benthic organisms (the silkworm and bivalve), and the results showed that AgNP and ionic Ag had a significantly enhanced effect on CAT activity, and AgNPs also significantly induced the SOD activity in silkworms. Jiang et al. [55] found the toxic effects of AgNPs on the aquatic plant Spirodela polyrhiza, and that exposure to AgNPs significantly enhanced SOD and CAT activities in the duckweed, while particle Ag on the micrometer level had no obvious effect on SOD and CAT activities. Lin et al. [40] found that nano titanium dioxide could cause an increase in the levels of reactive oxygen species and MDA in Chlorella vulgaris. Chen et al. [51] showed that nano titanium dioxide also increased the MDA in Chlamydomonas reinhardtii to its maximum value after 8 h of exposure. Some studies [48,56] have also studied the toxic effect of AgNPs on large fleas using the ion complexation method with the addition of cysteine, confirming that the biological toxicity of nanoparticles was greatly reduced after cysteine complexed the released ionic Ag. Therefore, nanoparticles can cause oxidative stress in organisms such as S. costatum, leading to toxic effects on organisms [57,58].
4. Conclusions
In this study, the biotoxic effects of AgNPs were found to comprise a combination of the solubilization of particles into toxic metal ions and the nature of the nanoparticles. The ion complexation with cysteine could reduce the biological toxicity of the released ionic Ag from the AgNPs. However, the induction of oxidative stress and the damage to the cell membrane comprised the dominant mechanism of toxicity for AgNPs. Therefore, the nature of nanometals in an aquatic environment is the key factor for understanding their biotoxic effect on phytoplankton. The study into the toxicity mechanism of nanoparticles in depth, and the possible morphological changes and dissolution of nanoparticles in different environments, is of great significance.
Author Contributions
Conceptualization, B.W.; Methodology, K.S. and B.W.; Software, K.S. and B.W.; Validation, D.C. and B.W.; Data curation, D.C., Y.Y. and B.W.; Writing—original draft preparation, K.S.; Writing—review and editing, B.W.; Visualization, Y.Y. and J.X.; Supervision, B.W.; Project administration, B.W.; Funding acquisition, J.X. and B.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Natural Science Foundation of Shandong Province (ZR2021QD051); the scientific research fund project of the National Natural Science Foundation of China (52070123); and Project of Shandong Province Higher Educational Young Innovative Talent Introduction and Cultivation Team [Wastewater Treatment and Resource Innovation Team].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mahjoubian, M.; Naeemi, A.S.; Moradi-Shoeili, Z.; Tyler, C.R.; Mansouri, B. Toxicity of Silver Nanoparticles in the Presence of Zinc Oxide Nanoparticles Differs for Acute and Chronic Exposures in Zebrafish. Arch. Environ. Con. Tox. 2023, 84, 1–17. [Google Scholar] [CrossRef]
- Bakr, Z.; Abdel-Wahab, M.; Thabet, A.A.; Hamed, M.; El-Aal, M.A.; Saad, E.; Faheem, M.; Sayed, A.E.-D.H. Toxicity of silver, copper oxide, and polyethylene nanoparticles on the earthworm Allolobophora caliginosa using multiple biomarkers. Appl. Soil Ecol. 2023, 181, 104681. [Google Scholar] [CrossRef]
- Kose, O.; Mantecca, P.; Costa, A.; Carrière, M. Putative adverse outcome pathways for silver nanoparticle toxicity on mammalian male reproductive system: A literature review. Part. Fibre Toxicol. 2023, 20, 1. [Google Scholar] [CrossRef] [PubMed]
- Lubick, N. Nanosilver toxicity: Ions, nanoparticles—Or both? Environ. Sci. Technol. 2008, 42, 8617. [Google Scholar] [CrossRef]
- Arora, S.; Rajwade, J.M.; Paknikar, K.M. Nanotoxicology and in vitro studies: The need of the hour. Toxicol. Appl. Pharm. 2012, 258, 151–165. [Google Scholar]
- Wang, L.-F.; Habibul, N.; He, D.-Q.; Li, W.-W.; Zhang, X.; Jiang, H.; Yu, H.-Q. Copper release from copper nanoparticles in the presence of natural organic matter. Water Res. 2015, 68, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Dei, R.C.H.; Wang, W.-X.; Guo, L. Marine diatom uptake of iron bound with natural colloids of different origins. Mar. Chem. 2003, 81, 177–189. [Google Scholar] [CrossRef]
- Chen, M.; Wang, W.X. bioavailability of natural colloid-bound iron to marine plankton: Influences of colloidal size and aging. Limnol. Oceanogr. 2001, 46, 1956–1967. [Google Scholar] [CrossRef]
- Peyrot, C.; Wilkinson, K.J.; Desrosiers, M.; Sauve, S. Effects of silver nanoparticles on soil enzyme activities with and without added organic matter. Environ. Toxicol. Chem. 2014, 33, 115–125. [Google Scholar] [CrossRef]
- Sharma, V.K.; Siskova, K.M.; Zboril, R.; Gardea-Torresdey, J.L. Organic-coated silver nanoparticles in biological and environmental conditions: Fate, stability and toxicity. Adv. Colloid Interface Sci. 2013, 204, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Saleh, N.; Kim, H.-J.; Phenrat, T.; Matyjaszewski, K.; Tilton, R.D.; Lowry, G.V. Ionic strength and composition affect the mobility of surface-modified Fe0 nanoparticles in water-saturated sand columns. Environ. Sci. Technol. 2008, 42, 3349–3355. [Google Scholar] [CrossRef]
- Buffet, P.-E.; Pan, J.-F.; Poirier, L.; Amiard-Triquet, C.; Amiard, J.-C.; Gaudin, P.; Faverney, C.R.-d.; Guibbolini, M.; Gilliland, D.; Valsami-Jones, E.; et al. Biochemical and behavioural responses of the endobenthic bivalve Scrobicularia plana to silver nanoparticles in seawater and microalgal food. Ecotox. Environ. Safe 2013, 89, 117–124. [Google Scholar] [CrossRef]
- Buffet, P.E.; Zalouk-Vergnoux, A.; Chatel, A.; Berthet, B.; Metais, I.; Perrein-Ettajani, H.; Poirier, L.; Luna-Acosta, A.; Thomas-Guyon, H.; Risso-de Faverney, C.; et al. A marine mesocosm study on the environmental fate of silver nanoparticles and toxicity effects on two endobenthic species: The ragworm Hediste diversicolor and the bivalve mollusc Scrobicularia plana. Sci. Total Environ. 2014, 470, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Turan, N.B.; Erkan, H.S.; Engin, G.O.; Bilgili, M.S. Nanoparticles in the aquatic environment: Usage, properties, transformation and toxicity—A review. Process Saf. Environ. 2019, 130, 238–249. [Google Scholar] [CrossRef]
- Garcés, M.; Cáceres, L.; Chiappetta, D.; Magnani, N.; Evelson, P. Current understanding of nanoparticle toxicity mechanisms and interactions with biological systems. New J. Chem. 2021, 45, 14328–14344. [Google Scholar] [CrossRef]
- Miao, A.-J.; Schwehr, K.A.; Xu, C.; Zhang, S.-J.; Luo, Z.; Quigg, A.; Santschi, P.H. The algal toxicity of silver engineered nanoparticles and detoxification by exopolymeric substances. Environ. Pollut. 2009, 157, 3034–3041. [Google Scholar] [CrossRef] [PubMed]
- Choi, O.; Hu, Z. Size dependent and reactive oxygen species related nanosilver toxicity to nitrifying bacteria. Environ. Sci. Technol. 2008, 42, 4583–4588. [Google Scholar] [CrossRef]
- Newton, K.M.; Puppala, H.L.; Kitchens, C.L.; Colvin, V.L.; Klaine, S.J. Silver nanoparticle toxicity to Daphnia magna is a function of dissolved silver concentration. Environ. Toxicol. Chem. 2013, 32, 2356–2364. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.X.Z.; Radisic, M. Organ-on-a-chip platforms for evaluation of environmental nanoparticle toxicity. Bioact. Mater. 2021, 6, 2801–2819. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, W.-X. Silver nanoparticle toxicity to the larvae of oyster Crassostrea angulata: Contribution of in vivo dissolution. Sci. Total Environ. 2023, 858, 159965. [Google Scholar] [CrossRef]
- Ringwood, A.H.; McCarthy, M.; Bates, T.C.; Carroll, D.L. The effects of silver nanoparticles on oyster embryos. Mar. Environ. Res. 2010, 69, S49–S51. [Google Scholar] [CrossRef] [PubMed]
- Azadikhah, D.; Yalsuyi, A.M.; Saha, S.; Saha, N.C.; Faggio, C. Biochemical and Pathophysiological Responses in Capoeta capoeta under Lethal and Sub-Lethal Exposures of Silver Nanoparticles. Water 2023, 15, 585. [Google Scholar]
- Liu, Z.; Malinowski, C.R.; Sepúlveda, M.S. Emerging trends in nanoparticle toxicity and the significance of using Daphnia as a model organism. Chemosphere 2022, 291, 132941. [Google Scholar] [CrossRef] [PubMed]
- Padhye, L.P.; Jasemizad, T.; Bolan, S.; Tsyusko, O.V.; Unrine, J.M.; Biswal, B.K.; Balasubramanian, R.; Zhang, Y.; Zhang, T.; Zhao, J.; et al. Silver contamination and its toxicity and risk management in terrestrial and aquatic ecosystems. Sci. Total Environ. 2023, 871, 161926. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Powers, K.; Wang, Y.; Zhou, H.; Roberts, S.M.; Moudgil, B.M.; Koopman, B.; Barber, D.S. Influence of Suwannee River humic acid on particle properties and toxicity of silver nanoparticles. Chemosphere 2012, 89, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Dube, E.; Okuthe, G.E. Engineered nanoparticles in aquatic systems: Toxicity and mechanism of toxicity in fish. Emerg. Contam. 2023, 9, 100212. [Google Scholar] [CrossRef]
- Liu, D.; Sun, J.; Zou, J.; Zhang, J. Phytoplankton succession during a red tide of Skeletonema costatum in Jiaozhou Bay of China. Mar. Pollut. Bull. 2005, 50, 91–94. [Google Scholar] [CrossRef]
- Guillard, R.; Ryther, J. Studies of marine planktonic diatoms I: Cyclotella nanai Hustedt and Detonula confervacea Cleva. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- He, Y.-L.; Bai, J.; Bai, X.-Y.; Li, K.-R. Effect of Nano-Silver on Planktonic-Bacterial Activity in Qingdao Coastal Waters. P. Ocean Univ. China 2020, 50, 76–82. [Google Scholar]
- Pisani, T.; Munzi, S.; Paoli, L.; Bačkor, M.; Loppi, S. Physiological effects of arsenic in the lichen Xanthoria parietina (L.) Th. Fr. Chemosphere 2011, 82, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Gomes, T.; Pereira, C.G.; Cardoso, C.; Pinheiro, J.P.; Cancio, I.; Bebianno, M.J. Accumulation and toxicity of copper oxide nanoparticles in the digestive gland of Mytilus galloprovincialis. Aquat. Toxicol. 2012, 118, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.W.; Song, I.H.; Um, S.H. Role of Physicochemical Properties in Nanoparticle Toxicity. Nanomaterials 2015, 5, 1351–1365. [Google Scholar] [CrossRef]
- Sukhanova, A.; Bozrova, S.; Sokolov, P.; Berestovoy, M.; Karaulov, A.; Nabiev, I. Dependence of Nanoparticle Toxicity on Their Physical and Chemical Properties. Nanoscale Res. Lett. 2018, 13, 44. [Google Scholar] [CrossRef]
- Zou, X.; Shi, J.; Zhang, H. Coexistence of silver and titanium dioxide nanoparticles: Enhancing or reducing environmental risks? Aquat. Toxicol. 2014, 154, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Burchardt, A.D.; Carvalho, R.N.; Valente, A.; Nativo, P.; Gilliland, D.; Garcìa, C.P.; Passarella, R.; Pedroni, V.; Rossi, F.; Lettieri, T. Effects of Silver Nanoparticles in Diatom Thalassiosira pseudonana and Cyanobacterium Synechococcus sp. Environ. Sci. Technol. 2012, 46, 11336–11344. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Gao, Y.; Lin, Z.; Yao, Z.; Zhang, W. The joint effects on Photobacterium phosphoreum of metal oxide nanoparticles and their most likely coexisting chemicals in the environment. Aquat. Toxicol. 2014, 154, 200–206. [Google Scholar] [CrossRef]
- Seitz, F.; Rosenfeldt, R.R.; Storm, K.; Metreveli, G.; Schaumann, G.E.; Schulz, R.; Bundschuh, M. Effects of silver nanoparticle properties, media pH and dissolved organic matter on toxicity to Daphnia magna. Ecotox Environ. Safe 2015, 111, 263–270. [Google Scholar] [CrossRef]
- Buchman, J.T.; Hudson-Smith, N.V.; Landy, K.M.; Haynes, C.L. Understanding Nanoparticle Toxicity Mechanisms to Inform. Redesign Strategies To Reduce Environmental Impact. Acc. Chem. Res. 2019, 52, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Ji, J.; Long, Z.; Yang, K.; Wu, F. The influence of dissolved and surface-bound humic acid on the toxicity of TiO2 nanoparticles to Chlorella sp. Water Res. 2012, 46, 4477–4487. [Google Scholar] [CrossRef]
- Aruoja, V.; Dubourguier, H.-C.; Kasemets, K.; Kahru, A. Toxicity of nanoparticles of CuO, ZnO and TiO2 to microalgae Pseudokirchneriella subcapitata. Sci. Total Environ. 2009, 407, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, N.B.; Von der Kammer, F.; Hofmann, T.; Baalousha, M.; Ottofuelling, S.; Baun, A. Algal testing of titanium dioxide nanoparticles—Testing considerations, inhibitory effects and modification of cadmium bioavailability. Toxicology 2010, 269, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Chen, B.; Sun, X.; Qu, K.; Ma, F.; Du, M. Interaction of TiO2 nanoparticles with the marine microalga Nitzschia closterium: Growth inhibition, oxidative stress and internalization. Sci. Total Environ. 2015, 508, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Perreault, F.; Oukarroum, A.; Melegari, S.P.; Matias, W.G.; Popovic, R. Polymer coating of copper oxide nanoparticles increases nanoparticles uptake and toxicity in the green alga Chlamydomonas reinhardtii. Chemosphere 2012, 87, 1388–1394. [Google Scholar] [CrossRef]
- Hull, M.S.; Vikesland, P.J.; Schultz, I.R. Uptake and retention of metallic nanoparticles in the Mediterranean mussel (Mytilus galloprovincialis). Aquat. Toxicol. 2013, 140–141, 89–97. [Google Scholar] [CrossRef]
- Vrieling, E.G.; Beelen, T.P.M.; van Santen, R.A.; Gieskes, W.W.C. Diatom silicon biomineralization as an inspirational source of new approaches to silica production. J. Biotechnol. 1999, 70, 39–51. [Google Scholar] [CrossRef]
- Navarro, E.; Baun, A.; Behra, R.; Hartmann, N.B.; Filser, J.; Miao, A.J.; Quigg, A.; Santschi, P.H.; Sigg, L. Environmental behavior and ecotoxicity of engineered nanoparticles to algae, plants, and fungi. Ecotoxicology 2008, 17, 372–386. [Google Scholar] [CrossRef]
- Navarro, E.; Piccapietra, F.; Wagner, B.; Marconi, F.; Kaegi, R.; Odzak, N.; Sigg, L.; Behra, R. Toxicity of Silver Nanoparticles to Chlamydomonas reinhardtii. Environ. Sci. Technol. 2008, 42, 8959–8964. [Google Scholar] [CrossRef]
- Ovečka, M.; Lang, I.; Baluška, F.; Ismail, A.; Illeš, P.; Lichtscheidl, I.K. Endocytosis and vesicle trafficking during tip growth of root hairs. Protoplasma 2005, 226, 39–54. [Google Scholar] [CrossRef]
- Oukarroum, A.; Bras, S.; Perreault, F.; Popovic, R. Inhibitory effects of silver nanoparticles in two green algae, Chlorella vulgaris and Dunaliella tertiolecta. Ecotox. Environ. Safe 2012, 78, 80–85. [Google Scholar] [CrossRef]
- Chen, L.; Zhou, L.; Liu, Y.; Deng, S.; Wu, H.; Wang, G. Toxicological effects of nanometer titanium dioxide (nano-TiO2) on Chlamydomonas reinhardtii. Ecotox. Environ. Safe 2012, 84, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Miao, A.J.; Zhang, X.Y.; Luo, Z.; Chen, C.S.; Chin, W.C.; Santschi, P.H.; Quigg, A. Zinc oxide–engineered nanoparticles: Dissolution and toxicity to marine phytoplankton. Environ. Toxicol. Chem. 2010, 29, 2814–2822. [Google Scholar] [CrossRef]
- Röhder, L.A.; Brandt, T.; Sigg, L.; Behra, R. Influence of agglomeration of cerium oxide nanoparticles and speciation of cerium (III) on short term effects to the green algae Chlamydomonas reinhardtii. Aquat. Toxicol. 2014, 152, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Buffet, P.-E.; Poirier, L.; Zalouk-Vergnoux, A.; Lopes, C.; Amiard, J.-C.; Gaudin, P.; Risso-de Faverney, C.; Guibbolini, M.; Gilliland, D.; Perrein-Ettajani, H.; et al. Biochemical and behavioural responses of the marine polychaete Hediste diversicolor to cadmium sulfide quantum dots (CdS QDs): Waterborne and dietary exposure. Chemosphere 2014, 100, 63–70. [Google Scholar] [CrossRef]
- Jiang, H.-S.; Qiu, X.-N.; Li, G.-B.; Li, W.; Yin, L.-Y. Silver nanoparticles induced accumulation of reactive oxygen species and alteration of antioxidant systems in the aquatic plant Spirodela polyrhiza. Environ. Toxicol. Chem. 2014, 33, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.M.; Wang, W.X. Comparison of acute and chronic toxicity of silver nanoparticles and silver nitrate to Daphnia magna. Environ. Toxicol. Chem. 2011, 30, 885–892. [Google Scholar] [CrossRef]
- Yu, Z.; Li, Q.; Wang, J.; Yu, Y.; Wang, Y.; Zhou, Q.; Li, P. Reactive Oxygen Species-Related Nanoparticle Toxicity in the Biomedical Field. Nanoscale Res. Lett. 2020, 15, 115. [Google Scholar] [CrossRef]
- Horie, M.; Tabei, Y. Role of oxidative stress in nanoparticles toxicity. Free Radic. Res. 2021, 55, 331–342. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).