Abstract
Quorum sensing is a communication system by which bacteria use signal molecules to induce a physiological response. In natural marine environments, quorum sensing is suspected to occur in regions with high cell densities. Free-living bacteria, however, are largely believed to exist at concentrations too low to make use of a density-dependent quorum-sensing system. Due to so-called ‘free-living’ bacteria inhabiting marine gels composed of exopolymeric substances occurring in an operationally defined colloidal fraction, it is possible that quorum-sensing molecules would also occur in this fraction. In this study, possible signaling molecules were collected from marine water, separated, and concentrated as some organic matter. Their identification with an Agrobacterium tumefaciens assay indicated the presence of AHLs. In fall 2002 and spring 2003, free-living (single cells) fraction of marine bacterioplankton was sampled and separated. Various AHLs were added to these disperse populations (109 cells/L). The studied bacterial communities responded with change in the thymidine incorporation. The results are discussed as existence of active reaction of marine free-living bacteria to signaling molecules even in sparse inhabiting marine gels. That there is a bacterial response to signaling molecules in so sparse populations is of great significance for marine and environmental sciences.
1. Introduction
Quorum sensing, QS [1], is the regulation of bacterial gene expression in response to fluctuations in local concentrations of chemical signal molecules. QS was first reported in the marine species Vibrio fischeri [2], which is a symbiont of squid and some fish. Acylated homoserine lactones (AHLs) were the first of several classes of signal molecules to be isolated and characterized [3]. The AHL induced responses observed in V. fisheri and all subsequent species found to utilize quorum sensing were observed to occur only at very high cell densities. This density-dependent response led to the belief that free-living bacteria, such as those found in the water column of natural aquatic systems, existed at densities too low to utilize quorum-sensing systems.
QS, first proposed by Fuqua et al. [4], is understood as communication with regards to space, i.e., bacteria communicate with each other in order to gain the ability to coordinate their activities. However, the sensing phenomenon can only occur when the number of bacteria reaches a certain density. Thus, if QS molecules were to be found in the fraction that does not sediment during ultracentrifugation, bacterial niches must exist in this fraction.
The oceans hold approximately 700 Gt of reduced carbon in different forms, with approximately 660 Gt C in the form of DOC (e.g., [5,6]). A substantial amount of this material is in the form of microscopic gels that are rich in nutrients and trace metals and readily available to bacterial colonization [6]. Marine gels (e.g., [7]) formed in the particulate fraction self-assemble reversibly from EPS precursors in the colloidal, macromolecular fraction. It has been observed that marine gels harbor orders of magnitude of higher concentrations of bacteria [5,6,7,8,9], which can obtain associated nutrients and carbon from these gels, and further modify their composition by degradation and secretion of their own EPS. We propose that QS molecules found after ultracentrifugation are derived from bacteria-inhabiting marine gels. Because gels reversibly assemble and dissemble, it can also be hypothesized that associated QS molecules would appear in the macromolecular fraction that would not be ultracentrifugable under the specified conditions.
Even though we did not directly show the existence of marine gels in the supernatant fraction after ultracentrifugation, it is well established that macromolecules that exist in the supernatant of seawater ultracentrifuged at 16,000 for 15 min (see methods below) are composed of exopolymeric substances, EPS [10,11], that make up the majority of marine gels [10,12]. Marine gels thus occur in both particulate and colloidal high molecular weight (HMW) fractions. About 60–70% of dissolved organic carbon, DOC, in waters of Galveston Bay and coastal sea areas is macromolecular in nature (i.e., colloidal organic carbon, COC ≤ 0.45 µm to ≥1 kDa; as reviewed in [12]). Importantly, the majority, i.e., 54 %, of the COC is identified as EPS (by summing the protein and polysaccharide fraction) in Galveston Bay waters [13]. Furthermore, it has been shown that COC can also contain other low molecular weight, LMW, molecules such as chlorophyll-a and other pigment molecules [14] with molecular weights of 900 Da, again likely contained in marine gels.
QS may control basic and fundamental processes in marine bacteria. Documenting the existence of these controls is crucial to our understanding of bacterial functioning. Potential bacterial processes controlled by quorum sensing may have a large impact on both coastal and open ocean environments. Coastal areas are important because they are more productive and more susceptible to anthropogenic influences than the open ocean. Pollutants may disturb quorum-sensing pathways, which may lead to the release of toxins or changes in bacterial activity at unexpected or unprecedented times. Such effects on bacterial processes may lead to unforeseen changes in water quality and may detrimentally affect finfish, shellfish and even human health. QS in marine environments may be involved in antibiotic production by bacteria. The discovery of quorum sensing in free-living bacteria inhabiting marine gels would open a new avenue in the search for novel antibiotics. The presence of quorum sensing in the macromolecular fraction of DOC, mostly composed of marine gels, may have a significant impact on the biochemical cycling of carbon, including dissolved organic carbon and particulate organic carbon flux. Recognizing that free-living bacteria associated with marine gels respond to quorum-sensing molecules indicates that these bacteria likely utilize QS and would be a large step forward in understanding bacterioplankton function.
QS has been found to regulate a number of behaviors that would be useful to marine bacteria: these include biofilm formation [15], production of antibiotics [16], toxicity ([17]), swarming [18], exopolysaccharide production [19] exoenzyme production [20] and entry into and emergence from stationary phase [21,22]. Thus, marine bacteria that inhabit regions such as particle aggregates where cell densities may be much higher than in the surrounding water column have been suspected to utilize quorum sensing [23,24]. Bacteria isolated from marine snow particles have indeed been shown to produce AHLs in pure laboratory cultures, but the biological relevance has not been documented [25]. Although quorum sensing in environments with low densities of cells has been generally discounted, previous studies have observed a possible role for AHLs in isolation and cultivation of bacterioplankton. Guan and Kamino [26] showed that the growth of filtered marine bacterioplankton communities was enhanced by the addition of AHLs. The authors, however, measured their abundance a full day after inoculation with AHLs. Thus, they were not able to document if an immediate response occurred due to the addition of AHLs. Also, because the only response seen was an increase in abundance, they could not definitively preclude the possibility that the bacteria involved were using the AHLs as substrate. Finally, the study did not address the potential implications or the biological relevance of QS in free-living marine bacteria.
In this study, we show that natural communities of “free-living” marine bacterioplankton in their natural medium do respond to quorum-sensing signal molecules, and they respond in a manner that precludes the possibility that they are using the AHLs as substrate. This discovery of a previously unknown control on free-living heterotrophic bacteria in the water column may allow us to link bacterial function to community composition and may also have large effects on bulk heterotrophic processes such as growth, respiration, and mineralization in marine waters.
2. Material and Methods
2.1. Extraction of AHLs
Surface water samples were collected at Pelican Island at the mouth of Galveston Bay, Texas in November 2002 for AHL extraction. Water samples were ultracentrifuged at 16,000× g for 15 min to remove cells and debris. Ultracentrifugation at 16,000× g for 15 min removes all particulate matter, and even removes HMW glutenin (MW of ~100–200 kDa or more) and other proteins into the pellet ([27]). Furthermore, microsomes (20–200 nm in size. https://www.beckman.com/resources/sample-type/extracellular-vesicles/microsomes (accessed on 15 May 2001)) will still stay in the supernatant after 15,000× g for 20 min. Thus, after 16,000× g for 15 min, the supernatant water will still have nanogels (smaller than 200 nm, i.e., ≤0.22 um filtered seawater) left, including associated QS molecules. With nanogels in the supernatant, these smaller gels can gradually grow into larger size gel particles (becoming microgels in hours or a few days; [5,7], and references therein), relevant for QS observations.
AHLs were then extracted with equal volumes of ethyl acetate (acidified with acetic acid to pH < 7.0). Samples were dried over MgSO4 and concentrated by gentle roto-evaporation [28]. Extracted AHLs were kept in acidified ethyl acetate at –20 °C until use.
2.2. Detection of AHLs—A. tumefaciens Bioassay
The presence of AHLs in water samples was indicated by use of a bacterial reporter strain. This strain (NTL4 (pCF218) (pCF372)) of Agrobacterium tumefaciens indicates the presence of AHLs by a blue colour produced by hydrolysis of 5-bromo-4-chloro-3-indolyl β-D-galactopyranoside (X-gal) by β-galactosidase. This enzyme is produced when NTL4 is grown in the presence of AHLs [29,30]. NTL4 was grown in LB media (Difco) or in AT minimal media [31].
AHLs extracted from 500 mL of bay water and stored in ethyl acetate were dried completely by speed-vac. Dried material with potential AHLs was then brought up in 100 µL of culture media and added to a 5–10 mL culture of A. tumefaciens grown in AT media in the presence of X-Gal. This culture was incubated at ~28 °C. AHLs were deemed present if the sample was blue after incubation and no color formation was observed in negative controls.
2.3. Bacterial Numbers in Marine Water
Bacterial abundance was determined by direct count under microscope on filters with acridine orange staining [32].
2.4. Experimental Protocol
The response of natural communities of free-living bacteria to the addition of AHLs was determined by measuring the incorporation of tritiated thymidine [33]. Seawater samples for experiments were filtered through baked GF/F filters (nominal pore size~0.7 µm) to remove all particles and particle-associated bacteria. After filtration, water samples were placed into sterile flasks and inoculated with commercially purified AHLs (Sigma-Aldrich, St. Louis, MO, USA). Commercially purified AHLs were received as a powder and brought up in acidified ethyl acetate and kept in 10 mM stock solutions at –20 °C. To inoculate samples, appropriate amounts of commercial AHLs from the stock solution were dried in 2.0 mL centrifuge tubes. These AHLs were then resuspended in 100 µL of sample and added back to the samples. Controls were inoculated with equal quantities of dried ethyl acetate. After addition of AHLs or dried ethyl acetate, 1.7 mL of sample were subsampled into 2.0 mL centrifuge tubes containing 25 nM thymidine then incubated at near in situ temperature (~18 °C in the fall, and ~20 °C in the spring) for 1–2 h. Bacterial incorporation rates were determined by addition of tritiated thymidine [34], and processed per the microcentrifugation method of Smith and Azam [34]. Student’s t-test was used to determine the significance of differences in thymidine incorporation between samples and controls.
3. Results and Discussion
3.1. Identification of Origin of Signaling Molecules in the Galveston Bay Water
Incubation of the indicator strain (Agrobacterium tumefaciens) with material extracted from Galveston Bay indicated the presence of AHLs in Galveston Bay water. Although control experiments indicated that impurities in the solvent did not cause a false positive, we cannot preclude the possibility that some other material coextracted from the seawater caused the positive signal. These results, however, did suggest that AHLs may be present in seawater and led us to investigate the effects of AHLs on free-living bacteria.
We suspected that AHLs would affect the endogenous free-living bacterial communities found in Galveston Bay. We elected to examine the growth of bacteria as measured by the incorporation of thymidine because we believed that any significant response to quorum-sensing molecules would have an observable effect on DNA synthesis. Previous works in estuarine and other marine systems have shown a distinct difference in community composition between bacteria associated with particles and free-living bacteria [34,35,36,37]. These studies showed the separation of particle-associated bacteria from “free-living” bacteria by filtration through 3 µm (or larger) filters. The GF/F filters used in this study are 0.7 µm nominal pore size. This very conservative filtration ensured that the remaining bacterial community was composed of non-aggregated, disperse bacteria.
3.2. Response of the Sparse Bacterial Community to AHLs, November 2002
To examine the effect of quorum-sensing signal molecules on free-living marine bacteria, we obtained four commercially available purified AHLs with carbon chain lengths of 4 (butanoyl), 7 (heptanoyl), 8 (octanoyl) and 12 (3-oxo-dodecanoyl) (Sigma). These AHLs were chosen to cover a broad spectrum of AHL sizes. Water was collected from Galveston Bay in the fall of 2002 (November) and again in the spring of 2003 (April) and filtered through a GF/F filter. We then added a cocktail containing all four AHLs at a concentration of 2 µM. Thus, it is unlikely that the overall AHL concentration would reach 2 µM in the water column, and it is possible that individual cells may be exposed to such concentrations of AHLs in a local environment. Thus, we used 2 µM in an attempt to ensure that all the free-living bacteria in the sample would be exposed to a sufficient concentration of AHLs to induce a detectable response. The addition of the purified AHLs caused a significant decrease (p < 0.01) in thymidine incorporation in the free-living bacteria (Figure 1) in the November samples.
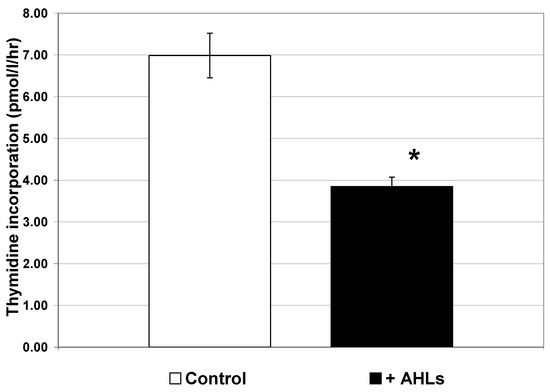
Figure 1.
Thymidine incorporation in free-living bacteria of Galveston Bay collected in November 2002 after addition of purified AHLs. AHLs were added as a 2 µM cocktail of four AHLs with carbon chain lengths of 4 (butyryl), 7 (heptanoyl), 8 (octanoyl) and 12 (3-oxo-dodecanoyl). Error bars represent standard error of the mean. Samples were filtered through a GF/F 0.7 µm filter prior to addition of AHLs. Addition of purified AHL cocktail significantly reduced thymidine incorporation in free-living bacteria (n = 6 for each treatment; * p < 0.01).
Bacteria use AHLs with different carbon chain lengths and substitutions to regulate specific behaviors. Different species of bacteria may use the same AHL to regulate different processes and individual species may utilize more than one AHL [38]. To investigate the specificity of the AHL(s) causing the reduction in thymidine incorporation, we added each of the commercially purified AHLs individually (at 2 µM) to free-living bacteria from Galveston Bay. In November, the only significant response in thymidine incorporation seen in this particular community of bacteria was to octanoyl, an AHL with a carbon chain length of 8 (Figure 2). Octanoyl caused a reduction in thymidine incorporation similar to that seen in the addition of the cocktail of AHL molecules. While these data indicate that octanoyl does cause a decrease in thymidine incorporation in this natural marine bacterial community, they does not indicate that octanoyl is the only AHL capable of invoking a response in free-living bacteria, because we did not test all AHLs. Additionally, the observed response may be specific to one species or several species of free-living bacteria in this particular community. However, the response to octanoyl alone does indicate that the bacterial response observed is to a specific AHL and not to the class of molecules. The lack of a significant change in thymidine incorporation due to addition of any of the other AHLs suggests that no contaminating material from the commercially prepared AHLs is affecting bacterial growth. Finally, the response seen was a decrease in thymidine incorporation indicating that the bacteria were not using the AHL as a substrate. Therefore, these free-living marine bacteria from Galveston Bay are indeed responding specifically to AHLs as signal molecules.
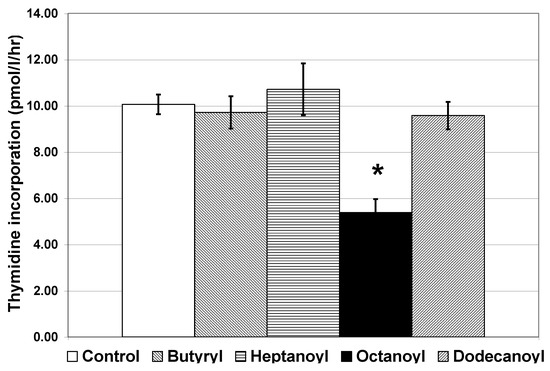
Figure 2.
Thymidine incorporation in free-living bacteria of Galveston Bay collected in November 2002 after addition of individual purified AHLs. AHLs were added separately at a 2 µM concentration. Error bars represent standard error of the mean. Samples were filtered through a GF/F 0.7 µm filter prior to addition of AHLs. Separate addition of purified AHLs had no significant effect on thymidine incorporation except for octanoyl, which significantly reduced thymidine incorporation in free-living bacteria (n = 4 for each treatment; * p < 0.01).
The species composition of the bacterial community present in these marine samples is unknown. The fact that the free-living bacteria respond to quorum-sensing signal molecules, however, is remarkable. Because free-living bacteria in the marine water column exist at cell densities well below those typically found in laboratory cultures or within particles, a density dependent response to this AHL-containing material might not be expected, but the observed response in the filtered fraction to quorum-sensing signal molecules suggests the presence of a quorum-sensing system in the free-living bacterial community.
3.3. Response of the Sparse Bacterial Community to AHLs, April 2003
Bacterial communities in marine systems have been shown to exhibit seasonal changes in both composition [39] and in metabolic capabilities [40]. Therefore, we repeated the AHL addition experiments in April of 2003. We obtained two additional AHLs, hexanoyl (carbon chain length of 6) and decanoyl (carbon chain length of 10), to expand the range of possible responses observed to AHL addition. Water was again collected from Galveston Bay and filtered through a GF/F filter. Each of the six commercially purified AHLs was added individually to the water samples and thymidine incorporation was measured. In these experiments, octanoyl had no significant effect on thymidine incorporation, while both decanoyl and dodecanoyl stimulated an increase in incorporation (Figure 3). The lack of a significant response to octanoyl indicated that the community structure had changed. These results indicate that several free-living species of marine bacteria may utilize quorum-sensing systems. Because the response seen was an increase in thymidine incorporation, we cannot preclude the possibility that the bacteria responding to decanoyl and dodecanoyl are utilizing those particular AHLs as substrate. This is unlikely, however, because these added AHLs constitute only ~1% or less of the total [41] or bioavailable DOC [42] in coastal waters of the Gulf of Mexico.
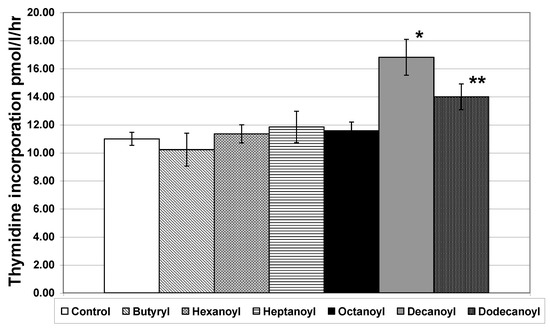
Figure 3.
Thymidine incorporation in free-living bacteria of Galveston Bay collected in April 2003 after addition of individual purified AHLs. AHLs were added separately at a 2 µM concentration. Error bars represent standard error of the mean. Samples were filtered through a GF/F 0.7 µm filter prior to addition of AHLs. Addition of purified AHLs had no significant effect on thymidine incorporation for butyryl, hexanoyl, heptanoyl or octanoyl. Decanoyl and dodecanoyl induced a significant increase in thymidine incorporation in the free-living bacterial community (n = 6 for each treatment; * p < 0.01, ** p < 0.05).
Thus, we have shown that free-living bacteria in Galveston Bay respond to AHLs. The abundance of cells in samples taken from Galveston Bay for the April 2003 experiments was ~1 × 109 cells L−1. This number of cells is several orders of magnitude lower than in most laboratory cultures in which quorum sensing has been observed. For quorum sensing to occur, free-living cells must be exposed to relatively high concentrations of signal molecules (in this case, AHLs). Quorum sensing probably occurs in the water column in microzones of high cell densities such as those that occur in particle or gel aggregates and inside marine biofilms. Bacterioplankton cells are not considered to exist at such high cell concentrations outside of particle aggregates. Other mechanisms may exist, however, to allow free-living cells to reach locally high cell concentrations without attachment. As mentioned before, reversible self-assembly of gels can be assumed to form over a continuum of sizes including the colloidal fraction that would not sink out during ultrafiltration or ultracentrifugation ([12], and references therein). Thus, gels in that fraction would also contain QS molecules. Free-living marine bacteria have been shown to use motility to loosely congregate around point-source particles of amino acids in the lab [43]. These loose congregations of free-living bacteria, if they indeed occur in the water column, may allow these bacteria to reach cell densities high enough to utilize quorum sensing. Alternatively, free-living bacteria may be overproducers of AHLs or they may be very sensitive to the presence of lower concentrations of AHLs. This sensitivity may allow free-living bacteria to react to small shifts in substrate availability or other environmental cues. These models are not mutually exclusive and, in fact, a combination of these models may be most fitting.
In the fall experiments with Galveston Bay bacterial communities, bacteria responded to AHL addition by exhibiting a significant decrease in cell division. We hypothesize that this may be due to the free-living bacteria entering a stationary phase. AHLs and other autoinducer molecules have been shown to be important in the induction of entry into a stationary phase for certain bacteria species [21,22]. In cultures, when cell density becomes high, bacteria may shut down or slow growth processes due to competition for available substrate. A similar process may be occurring in the responding species of free-living bacterial communities in the water column of coastal areas such as Galveston Bay, particularly in the species present late in the year as water temperature and phosphate concentrations decrease [44] and bacterial activity slows. This decrease in resources may also occur in marine gels that will reversibly assemble then disassemble. A cell density high enough for QS in marine gels may mean that nutrient availability and population collapse is imminent and cells may use the signal to stop dividing because lean times are approaching.
In support of our explanation for the November results, [45] showed that Vibrio vulnificus, a gram negative bacterium and AHL producer, shows near minimum abundance in Galveston Bay waters in the fall of 2000, corresponding to lower temperature and salinity of the water. For spring/summer of 2001, they showed near maximum abundance. Furthermore, Santschi [44] showed that in general, phosphate concentrations in Galveston Bay waters are highest in spring/summer, while near a minimum in the fall/winter months. Total nitrogen loadings of Galveston Bay are very high [46], one of the highest of US estuaries, and most likely are not limiting during November 2002 and April 2003.
In April, a different response was seen in response to the addition of AHLs. An increase in thymidine incorporation was observed due to decanoyl and dodecanoyl, while octanoyl did not induce a response. These changes in the behavior of the natural bacterial community may be due to a change in community structure. The mechanism for using quorum sensing between the two communities may be similar, however, with quorum sensing occurring as bacteria congregate about food sources, i.e., marine gels. The response of the species in the April assemblage is entirely different from the response seen in the November community. The responding bacteria in the April community may utilize AHLs to produce exoenzymes or to shift metabolism from cell maintenance to cell growth as the water becomes warmer and nutrients more plentiful. As the responding bacteria are likely different species from the November responders (lower temperature and nutrients), they may utilize QS at a lower cell or molecule density as the gel is aggregating and thus shift to cell growth as nutrients become plentiful. Although these hypotheses are presently merely speculation, investigation of these and other hypotheses may give researchers important insights into the functions of free-living bacterial communities in marine ecosystems.
While there are newer methods and approaches to studying QS using AHLs, our approach is robust for showing the presence of QS bacteria in the water sample. In the absence of a clearly positive response, however, our approach might be more ambiguous.
4. Conclusions
Our work showed that (i) bacteria responded to QS autoinducer molecules in their natural medium. Thus, marine water contained some signaling molecules which provoked bacterial reactions (at least—of A. tumefaciens), (ii) at least some of these agents were AHLs, (iii) dispersed populations of filtered marine bacterioplankton showed response to addition of AHLs, (iv) bacterial populations sampled in 6 months confirmed response but for different AHLs (the reasons of the change are unknown: they may be related both to changes in population composition or in chemical composition of the water); and (v) the concentrations of AHLs that were used were high but permitted to make this investigation as a model. (vi) In summary, the presented work showed bacterial response to signaling molecules in a highly dispersed bacterial community. Natural communities of free-living bacteria are likely living within reversibly forming marine gels, as has been shown previously. These results raise the possibility that quorum-sensing systems may be present throughout aquatic systems as well as in other environments presently discounted as sites for quorum-sensing activity due to low cell densities. Quorum sensing may thus play a vital, and so far unsuspected, role in the carbon cycle and nutrient regeneration in the world’s oceans, as well as in engineered systems (wastewater treatment). Determining the processes controlled by quorum sensing and the extent to which it is utilized in so-called ‘free-living’ marine bacteria, and other free-living bacterial communities, offers an exciting challenge for future endeavors.
Author Contributions
Conceptualization, G.E.S.J. and P.H.S.; methodology, G.E.S.J.; software, G.E.S.J.; validation; formal analysis, G.E.S.J.; investigation, G.E.S.J.; resources, G.E.S.J.; writing—original draft preparation, G.E.S.J.; writing—review and editing, P.H.S.; visualization, G.E.S.J.; supervision, P.H.S.; project administration, P.H.S.; funding acquisition, P.H.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by [Texas Institute of Oceanography].
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors wish to thank C. Fuqua for his kind donation of the A. tumefaciens reporter strain. We thank H. Schultz for comments. This work was supported by the Texas Institute of Oceanography.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hmelo, L.R. Quorum Sensing in Marine Microbial Environments. Annu. Rev. Mar. Sci. 2017, 9, 257–258. [Google Scholar] [CrossRef]
- Nealson, K.H.; Platt, T.; Hastings, J.W. Cellular control of the synthesis and activity of the bacterial luminescence system. J. Bacteriol. 1970, 104, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, A.; Burlingame, A.L.; Eberhard, C.; Kenyon, G.L.; Nealson, K.H.; Oppenheimer, N.J. Structural identification of autoinducer of Photobacterium fischeri. Biochemistry 1981, 20, 2444–2449. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Verdugo, P.; Santschi, P.H. Polymer dynamics of DOC networks and gel formation in seawater. Deep. Sea Res. Part II 2010, 57, 1486–1493. [Google Scholar] [CrossRef]
- Verdugo, P. Marine Microgels. Annu. Rev. Mar. Sci. 2012, 4, 375–400. [Google Scholar] [CrossRef]
- Quigg, A.; Santschi, P.H.; Burd, A.; Chin, W.-C.; Kamalanathan, M.; Xu, C.; Ziervogel, K. From Nano-Gels to Marine Snow: A Synthesis of Gel Formation Processes and Modeling Efforts Involved with Particle Flux in the Ocean. Gels 2021, 7, 114. [Google Scholar] [CrossRef]
- Engel, A.; Endres, S.; Galgani, L.; Schartau, M. Marvelous Marine Microgels: On the Distribution and Impact of Gel-Like Particles in the Oceanic Water-Column. Front. Mar. Sci. 2020, 7, 405. [Google Scholar] [CrossRef]
- Chin, W.-C.; Orellana, M.V.; Verdugo, P. Spontaneous assembly of marine dissolved organic matter into polymer gels. Nature 1998, 391, 568–572. [Google Scholar] [CrossRef]
- Santschi, P.H.; Balnois, E.; Wilkinson, K.; Zhang, J.; Buffle, J.; Guo, L. Fibrillar polysaccharides in marine macromolecular organic matter, as imaged by Atomic Force Microscopy and Transmission Electron Microscopy. Limnol. Oceanogr. 1998, 43, 896–908. [Google Scholar] [CrossRef]
- Verdugo, P.; Alldredge, A.L.; Azam, F.; Kirchman, D.L.; Passow, U.; Santschi, P.H. The oceanic gel phase: A bridge in the DOM–POM continuum. Mar. Chem. 2004, 92, 67–85. [Google Scholar] [CrossRef]
- Guo, L.; Santschi, P.H. Ultrafiltration and its applications to sampling and characterisation of aquatic colloids. IUPAC Ser. Anal. Phys. Chem. Environ. Syst. 2007, 10, 159. [Google Scholar]
- Xu, C.; Chin, W.-C.; Lin, P.; Chen, H.M.; Lin, P.; Chiu, M.-C.; Waggoner, D.C.; Xing, W.; Sun, L.; Schwehr, K.A.; et al. Marine Gels, Extracellular Polymeric Substances (EPS) and Transparent Exopolymeric Particles (TEP) in natural seawater and seawater contaminated with a water accommodated fraction of Macondo oil surrogate. Mar. Chem. 2019, 215, 103667. [Google Scholar] [CrossRef]
- Bianchi, T.S.; Lambert, C.; Santschi, P.H.; Baskaran, M.; Guo, L. Plant pigments as biomarkers of high-molecular-weight dissolved organic carbon. Limnol. Oceanogr. 1995, 40, 422–428. [Google Scholar] [CrossRef]
- Swift, S.; Karlyshev, A.V.; Fish, L.; Durant, E.L.; Winson, M.K.; Chhabra, S.R.; Williams, P.; Macintyre, S.; Stewart, G.S. Quorum sensing in Aeromonas hydrophila and Aeromonas salmonicida: Identification of the LuxRI homologs AhyRI and AsaRI and their cognate N-acylhomoserine lactone signal molecules. J. Bacteriol. 1997, 179, 5271–5281. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.W.; Gong, F.; Daykin, M.M.; Williams, P.; Pierson, L.S. N-acyl-homoserine lactone-mediated regulation of phenazine gene expression by Pseudomonas aereofaciens 30-84 in the wheat rhizosphere. J. Bacteriol. 1997, 179, 7663–7670. [Google Scholar] [CrossRef] [PubMed]
- Dunphy, G.; Miyamoto, C.; Meighen, E. A homoserine lactone autoinducer regulates virulence of an insect-pathogenic bacterium, Xenorhabdus nematophilus (Enterobacteriaceae). J. Bacteriol. 1997, 179, 5288–5291. [Google Scholar] [CrossRef]
- Eberl, L.; Winson, M.K.; Sternberg, C.; Stewart, G.S.; Christiansen, G.; Chharbra, G.; Bycroft, B.W.; Williams, P.; Molin, S.; Givskov, M. Involvement of N-acyl-homoserine lactone autoinducers in controlling the multicellular behavior of Serratia liquefaciens. Mol. Microbiol. 1996, 20, 127–136. [Google Scholar] [CrossRef] [PubMed]
- von Bodman, S.B.; Farrand, S.K. Capsular polysaccharide biosynthesis and pathogenicity in Erwinia stewartii require induction by an N-acylhomoserine lactone autoinducer. J. Bacteriol. 1995, 177, 5000–5008. [Google Scholar] [CrossRef]
- Brint, J.M.; Ohman, D.E. Synthesis of multiple exoproducts in Pseudomonas aeruginosa is under the control of RhlR-RhlI, another set of regulators in strain PAOl with homology to the autoinducer-responsive LuxR-LuxI family. J. Bacteriol. 1995, 177, 7155–7163. [Google Scholar] [CrossRef]
- Gray, K.M.; Pearson, J.P.; Downie, J.A.; Boboye, B.E.A.; Greenberg, E.P. Cell-to-cell signalling in the symbiotic nitrogen-fixing bacterium Rhizobium leguminosarum: Autoinduction of a stationary phase and rhizosphere-expressed genes. J. Bacteriol. 1996, 178, 372–376. [Google Scholar] [CrossRef]
- Lazazzera, B.A. Quorum sensing and starvation: Signals for entry into stationary phase. Curr. Opin. Microbiol. 2000, 3, 177–182. [Google Scholar] [CrossRef]
- Ploug, H.; Grossart, H.P.; Azam, F.; Jørgensen, B.B. Photosynthesis, respiration, and carbon turnover in sinking marine snow from surface waters of Southern California Bight: Implications for the carbon cycle in the ocean. Mar. Ecol. Prog. Ser. 1999, 179, 1–11. [Google Scholar] [CrossRef]
- Kiorboe, T. Colonization of marine snow aggregates by invertebrate zooplankton: Abundance, scaling, and possible role. Limnol. Oceanogr. 2000, 45, 479–484. [Google Scholar] [CrossRef]
- Gram, L.; Grossart, H.-P.; Schlingloff, A.; Kiørboe, T. Possible quorum sensing in marine snow bacteria: Production of acylated homoserine lactones by Roseobacter strains isolated from marine snow. Appl. Environ. Microbiol. 2002, 68, 4111–4116. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.L.; Kamino, K. Bacterial response to siderophore and quorum-sensing chemical signals in the seawater microbial community. BMC Microbiol. 2001, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- van Herpen, T.W.J.M.; Cordewener, J.H.G.; Klok, H.J.; Freeman, J.; America, A.H.P.; Bosch, D.; Smulders, M.J.M.; Gilissen, L.J.W.J.; Shewry, P.R.; Hamer, R.J. The origin and early development of wheat glutenin particles. J. Cereal Sci. 2008, 48, 870–877. [Google Scholar] [CrossRef]
- Shaw, P.D.; Ping, G.; Daly, S.L.; Cha, C.; Cronan, J.E.; Rinehart, K.L.; Farrand, S.K. Detecting and characterizing N-acyl-homoserine lactone signal molecules by thin-layer chromatography. Proc. Natl. Acad. Sci. USA 1997, 94, 6036–6041. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, C.; Winans, S.C. Conserved cis-acting promoter elements are required for density-dependent transcription of Agrobacterium tumefaciens conjugal transfer genes. J. Bacteriol. 1996, 178, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.-Q.; Clemente, T.E.; Farrand, S.K. Construction of a Derivative of Agrobacterium tumefaciens C58 That Does Not Mutate to Tetracycline Resistance. Mol. Plant-Microbe Interact. 2001, 14, 98–103. [Google Scholar] [CrossRef]
- Tempe, J.; Petit, A.; Holsters, M.; Montagu, M.V.; Schell, J. Thermosensitive step associated with transfer of the TI plasmid during conjugation: Possible relation to transformation in crown gall. Proc. Natl. Acad. Sci. USA 1977, 74, 2848–2849. [Google Scholar] [CrossRef] [PubMed]
- DeLong, E.F.; Franks, D.G.; Alldredge, A.L. Phylogenetic diversity of aggregate-attached vs. free-living marine bacterial assemblages. Limnol. Oceanogr. 1993, 38, 924–934. [Google Scholar] [CrossRef]
- Fuhrman, J.A.; Azam, F. Thymidine incorporation as a measure of heterotrophic bacterioplankton production in marine surface waters: Evaluation and field results. Mar. Biol. 1982, 66, 109–120. [Google Scholar] [CrossRef]
- Smith, D.C.; Azam, F. A simple, economical method for measuring bacterial protein synthesis rates in seawater using 3H-leucine. Mar. Microb. Food Webs. 1992, 6, 107–114. [Google Scholar]
- Bidle, K.D.; Fletcher, M. Comparison of free-living and particle-associated bacterial communities in the Chesapeake Bay by stable low-molecular-weight RNA analysis. Appl. Environ. Microbiol. 1995, 61, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Acinas, S.G.; Rodríguez-Valera, F.; Pedrós-Alió, C. Spatial and temporal variation in marine bacterioplankton diversity as shown by RFLP fingerprinting of PCR amplified 16S rDNA. FEMS Microbiol. Ecol. 1997, 24, 27–40. [Google Scholar] [CrossRef]
- Crump, B.C.; Armbrust, E.V.; Baross, J.A. Phylogenetic analysis of particle-attached and free-living bacterial communities in the Columbia River, its estuary, and the adjacent coastal ocean. Appl. Environ. Microbiol. 1999, 65, 3192–3204. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, N.A.; Barnard, A.M.L.; Slater, H.; Simpson, N.J.L.; Salmond, G.P.C. Quorum-sensing in gram negative bacteria. FEMS Microbiol. Rev. 2001, 25, 365–404. [Google Scholar] [CrossRef]
- Kelly, K.M.; Chistoserdov, A.Y. Phylogenetic analysis of the succession of bacterial communities in the Great South Bay (Long Island). FEMS Microbiol. Ecol. 2001, 35, 85–95. [Google Scholar] [CrossRef]
- Schultz, G.E.J.; Ducklow, H.W. Changes in bacterioplankton metabolic capabilities along a salinity gradient in the York River estuary, Virginia, USA. Aquat. Microb. Ecol. 2000, 22, 163–174. [Google Scholar] [CrossRef]
- Guo, L.; Santschi, P.H. Isotopic and elemental characterization of colloidal organic matter from the Chesapeake Bay and Galveston Bay. Mar. Chem. 1997, 59, 1–15. [Google Scholar] [CrossRef]
- Amon, R.M.W.; Benner, R. Rapid cycling of high-molecular-weight dissolved organic matter in the ocean. Nature 1994, 369, 549–552. [Google Scholar] [CrossRef]
- Barbara, G.M.; Mitchell, J.G. Marine bacterial organisation around point-like sources of amino acids. Fems. Microbiol. Ecol. 2003, 43, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Santschi, P.H. Seasonality of nutrient concentrations in Galveston Bay. Mar. Environ. Res. 1995, 40, 337–362. [Google Scholar] [CrossRef]
- Lin, M.; Payne, D.A.; Schwarz, J.R. Intraspecific Diversity of Vibrio vulnificus in Galveston Bay Water and Oysters asDetermined by Randomly Amplified Polymorphic DNA PCR. Appl. Environ. Microbiol. 2003, 69, 3170–3317. [Google Scholar] [CrossRef] [PubMed]
- Warnken, K.W.; Santschi, P.H. Sediment and trace metal delivery from the Trinity River watershed to Galveston Bay and the Gulf of Mexico. Estuaries Coasts 2009, 32, 158–172. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).