
Mediterranean Aquaculture and Genetic Pollution: A Review Combined with Data from a Fish Farm Evaluating the Ecological Risks of Finfish Escapes

 ,
,  ,
, 
 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Evidence for Aquaculture Driven Evolution?
3. Population Genetic Structure of the Main Farmed Mediterranean Marine Fish Species, S. aurata and D. labrax
4. Sea Bream Escapes—A Case Report in Evoikos Gulf, Central Greece
5. Consequences and Proposed Strategies for Risk Reduction of Fish Escapes
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO (Ed.) The State of World Fisheries and Aquaculture—Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar]
- Cossu, P.; Scarpa, F.; Sanna, D.; Lai, T.; Dedola, G.L.; Curini-Galletti, M.; Mura, L.; Fois, N.; Casu, M. Influence of genetic drift on patterns of genetic variation: The footprint of aquaculture practices in Sparus aurata (Teleostei: Sparidae). Mol. Ecol. 2019, 28, 3012–3024. [Google Scholar] [CrossRef] [PubMed]
- European Commission. 2013. Available online: https://cordis.europa.eu/project/id/6540/reporting (accessed on 15 May 2023).
- Dempster, T.; Moe, H.; Fredheim, A.; Sanchez-Jerez, P. Escapes of Marine Fish from Sea-Cage Aquaculture in the Mediterranean Sea: Status and Prevention. CIESM Work. Monogr. 2007, 32, 55–60. Available online: www.ciesm.org/online/monographs/Lisboa.html (accessed on 15 May 2023).
- Christophilogiannis, P.; Theodorou, J. Insurance Requirements of the Greek Mariculture Industry during the period 1986–1994 (1st semester). In Aquaculture Production Economics. Cah. Options Mediterr. 1995, 14, 239–246. [Google Scholar]
- Τheodorou, J.; Τzovenis, I. Aquaculture Insurance. In Encyclopedia of Actuarial Sciences; Teugels, J., Sundt, B., Eds.; John Willey & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- ECASA Project. 2008. Available online: https://cordis.europa.eu/project/id/226885/reporting (accessed on 15 May 2023).
- Theodorou, J.A.; Kotsonis, A.; Sorgeloos, P.; Viaene, J.; Tzovenis, I. Risk assessment of seabass and seabream aquaculture based on insurance claims in Greece. In Economics of Fish Resources and Aquatic Ecosystems: Balancing Uses, Balancing Costs, Proceedings of the Fifteenth Biennial Conference of the International Institute of Fisheries Economics & Trade, Montpellier, France, 13–16 July 2010; Shriver, A.L., Ed.; International Institute of Fisheries Economics & Trade: Corvallis, OR, USA, 2010; ISBN 0-9763432-6-6. [Google Scholar]
- Jackson, D.; Drumm, A.; McEvoy, S.; Jensen, Ø.; Mendiola, D.; Gabiña, G.; Borg, J.A.; Papageorgiou, N.; Karakassis, Y.; Black, K.D. A pan-European valuation of the extent, causes and cost of escape events from sea cage fish farming. Aquaculture 2015, 436, 21–26. [Google Scholar] [CrossRef]
- Holmen, I.M.; Utne, I.B.; Haugen, S. Identification of safety indicators in aquaculture operations based on fish escape report data. Aquaculture 2021, 544, 737143. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Papadakis, I.E.; Papadakis, V.M.; Glaropoulos, A.; Lamprianidou, F.; Kentouri, M. Escape-related behavior of juvenile gilthead sea bream (Sparus aurata) versus rearing density in experimental conditions. J. Biol. Res. 2013, 20, 208–216. [Google Scholar]
- Glaropoulos, A.; Papadakis, V.M.; Papadakis, I.E.; Georgara, A.; Kentouri, M. Sea bream interactions toward the aquaculture net due to the presence of micro-fouling. Aquac. Int. 2014, 22, 1203–1214. [Google Scholar] [CrossRef]
- Papadakis, I.E.; Papadakis, V.M.; Glaropoulos, A.; Petroutsos, S.I.; Gonzalez, L.P.; Kentouri, M. Do the visual conditions at the point of escape affect European sea bass escape behavior? Medit. Mar. Sci. 2013, 14, 172–178. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Izquierdo-Gomez, D.; Uglem, I.; Sanchez-Jerez, P. Aggregations of bluefish Pomatomus saltatrix (L.) at Mediterranean coastal fish farms: Seasonal presence, daily patterns and influence of farming activity. Environ. Biol. Fish. 2015, 98, 499–510. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Borg, J.A.; Segvic-Bubic, T.; Tomas-setti, P.; Ozgu€l, A.; Sanchez-Jerez, P. Aggregations of wild Atlantic Bluefin Tuna (Thunnus thynnus L.) at Mediterranean offshore fish farm sites: Environmental and management considerations. Fish. Res. 2015, 164, 178–184. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Izquierdo-Gomez, D.; Sanchez- Jerez, P. First report of a swordfish (Xiphias gladius Linnaeus, 1758) beneath open-sea farming cages in the Western Mediterranean Sea. Mediterr. Mar. Sci. 2014, 15, 72–73. [Google Scholar] [CrossRef] [Green Version]
- Díaz-López, B. Bottlenose dolphins and aquaculture: Interaction and site fidelity on the north-eastern coast of Sardinia (Italy). Mar. Biol. 2012, 159, 2161–2172. [Google Scholar] [CrossRef]
- Güçlüsoy, H.; Savaş, Y. Interaction between monk seals Monachus monachus (Hermann, 1779) and marine fish farms in the Turkish Aegean and management of the problem. Aquac. Res. 2003, 34, 777–783. [Google Scholar] [CrossRef] [Green Version]
- FEAP. European Aquaculture Production Report 2014–2019. 2020. Available online: https://feap.info/wp-content/uploads/2020/10/20201007_feap-production-report-2020.pdf (accessed on 2 August 2022).
- Moretti, A.; Pedini Fernandez-Criado, M.; Cittolin, G.; Guidastri, R. Manual on Hatchery Production of Seabass and Gilthead Seabream; FAO: Rome, Italy, 1999; pp. 1–194. [Google Scholar]
- Theodorou, J. Current & Future Technological Trends of European Seabass-Seabream Culture. Rev. Fish. Sci. 2002, 10, 529–543. [Google Scholar]
- Theodorou, J.A.; Perdikaris, C.; Filippopoulos, N.G. Evolution through Innovation in Aquaculture: The Case of the Hellenic Mariculture Industry (Greece). J. Appl. Aquac. 2015, 27, 160–181. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Izquierdo-Gomez, D.; Forcada, A.; Fernandez-Jover, D.; Toledo-Guedes, K.; Valle, C.; Sanchez-Jerez, P. Recapturing fish escapes from coastal farms in the western Mediterranean Sea: Insights for potential contingency plans. Ocean Coast. Manag. 2018, 151, 69–76. [Google Scholar] [CrossRef]
- Grigorakis, K.; Rigos, G. Aquaculture effects on environmental and public welfare—The case of Mediterranean mariculture. Chemosphere 2011, 85, 899–919. [Google Scholar] [CrossRef]
- Grant, W.S.; Jasper, J.; Bekkevold, D.; Adkison, M. Responsible genetic approach to stock restoration, sea ranching and stock enhancement of marine fishes and invertebrates. Rev. Fish Biol. Fish. 2017, 27, 615–649. [Google Scholar] [CrossRef]
- Somarakis, S.; Pavlidis, M.; Saapoglou, C.; Tsigenopoulos, C.S.; Dempster, T. Evidence for ‘escape through spawning’ in large gilthead sea bream Sparus aurata reared in com- mercial sea-cages. Aquac. Environ. Inter. 2013, 3, 135–152. [Google Scholar] [CrossRef] [Green Version]
- Theodorou, J.A.; Perdikaris, C.; Venou, B. Origin of broodstock and effects on the deformities of gilthead sea bream (Sparus aurata L. 1758) in a Mediterranean commercial hatchery. Int. Aquat. Res. 2016, 8, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Price, E.O. Behavioral development in animals undergoing domestication. Appl. Anim. Behav. Sci. 1999, 65, 245–271. [Google Scholar] [CrossRef]
- Zeder, M.A. Core questions in domestication research. Proc. Natl. Acad. Sci. USA 2015, 112, 3191–3198. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J. Evolution, consequences and future of plant and animal domestication. Nature 2002, 418, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Balon, E.K. About the oldest domesticates among fishes. J. Fish Biol. 2004, 65, 1–27. [Google Scholar] [CrossRef]
- Houston, R.D.; Bean, T.P.; Macqueen, D.J.; Gundappa, M.K.; Jin, Y.H.; Jenkins, T.L.; Selly, S.L.C.; Martin, S.A.M.; Stevens, J.R.; Santos, E.M.; et al. Harnessing genomics to fast-track genetic improvement in aquaculture. Nat. Rev. Genet. 2020, 21, 389–409. [Google Scholar] [CrossRef]
- Duarte, C.M.; Marbá, N.; Holmer, M. Rapid domestication of marine species. Science 2007, 316, 382–383. [Google Scholar] [CrossRef]
- Teletchea, F. Fish domestication in aquaculture: 10 unanswered questions. Anim. Front. 2021, 11, 87–91. [Google Scholar] [CrossRef]
- Teletchea, F. Domestication of marine fish species: Update and perspectives. J. Mar. Sci. Eng. 2015, 3, 1227–1243. [Google Scholar] [CrossRef] [Green Version]
- Teletchea, F.; Fontaine, P. Levels of domestication in fish: Implications for the sustainable future of aquaculture. Fish Fish. 2014, 15, 181–195. [Google Scholar] [CrossRef]
- FAO. The State of the World’s Aquatic Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2019; Available online: https://www.fao.org/3/CA5256EN/CA5256EN.pdf (accessed on 15 June 2023).
- Janssen, K.; Chavanne, H.; Berentsen, P.; Komen, H. Impact of selective breeding on European aquaculture. Aquaculture 2017, 472, 8–16. [Google Scholar] [CrossRef]
- Lorenzen, K.; Beveridge, M.C.; Mangel, M. Cultured fish: Integrative biology and management of domestication and interactions with wild fish. Biol. Rev. Camb. Philos. Soc. 2012, 87, 639–660. [Google Scholar] [CrossRef]
- Atalah, J.; Sanchez-Jerez, P. Global assessment of ecological risks associated with farmed fish escapes. Glob. Ecol. Conserv. 2020, 21, e00842. [Google Scholar] [CrossRef]
- Sola, L.; Moretti, A.; Crosetti, D.; Karaiskou, N.; Magoulas, A.; Rossi, A.R.; Rye, M.; Triantafyllidis, A.; Tsigenipoulos, C.S. Genetic Effects of Domestification, Culture and Breeding of Fish and Shellfish, and Their Impact on Wild Populations. Gilthead Seabream-Sparus aurata. In Genetic Impact of Aquaculture Activities on Native Populations. Genetic Final Scientific Report (EU Contract No. RICA-CT-2005-022802); Svasand, T., Crosetti, D., Garcia-Vázquez, E., Verspoor, E., Eds.; 2007; pp. 47–54. Available online: https://www.researchgate.net/profile/Philip-Mcginnity/publication/268359165_Research_Priorities_for_Modelling/links/547459fb0cf2778985abd9ef/Research-Priorities-for-Modelling.pdf#page=47 (accessed on 25 August 2022).
- Arabaci, M.; Yilmaz, Y.; Ceyhun, S.B.; Erdogan, O.; Dorlay, H.G.; Diler, I.; Akhan, S.; Kocabas, M.; Ozdemir, K.; Koyun, H.; et al. A Review on Population Characteristics of Gilthead Seabream (Sparus aurata). J. Anim. Vet. Adv. 2010, 9, 976–981. [Google Scholar] [CrossRef] [Green Version]
- Chavanne, H.; Janssen, K.; Hofherr, J.; Contini, F.; Haffray, P.; Aquatrace Consortium; Komen, H.; Nielsen, E.E.; Bargelloni, L. A comprehensive survey on selective breeding programs and seed market in the European aquaculture fish industry. Aquac. Int. 2016, 24, 1287–1307. [Google Scholar] [CrossRef]
- Janssen, K.; Saatkamp, H.; Komen, H. Cost-benefit analysis of aquaculture breeding programs. Genet. Sel. Evol. 2018, 50, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žužul, I.; Šegvić-Bubić, T.; Talijančić, I.; Džoić, T.; Lepen Pleić, I.; Beg Paklar, G.; Ivatek-Šahdan, S.; Katavić, I.; Grubišić, L. Spatial connectivity pattern of expanding gilthead seabream populations and its interactions with aquaculture sites: A combined population genetic and physical modelling approach. Sci. Rep. 2019, 9, 14718. [Google Scholar] [CrossRef] [Green Version]
- Chaoui, L.; Kara, M.H.; Quignard, J.P.; Faure, E.; Bonhomme, F. Strong genetic differentiation of the gilthead sea bream Sparus aurata (L., 1758) between the two western banks of the Mediterranean). C.R. Biol. 2009, 332, 329–335. [Google Scholar] [CrossRef]
- Gkagkavouzis, K.; Karaiskou, N.; Katopodi, T.; Leonardos, I.; Abatzopoulos, T.J.; Triantafyllidis, A. The genetic population structure and temporal genetic stability of gilthead sea bream Sparus aurata populations in the Aegean and Ionian Seas, using microsatellite DNA markers. J. Fish Biol. 2019, 94, 606–613. [Google Scholar] [CrossRef]
- Alarcón, J.A.; Magoulas, A.; Georgakopoulos, T.; Zouros, E.; Alvarez, M.C. Genetic comparison of wild and cultivated European populations of the gilthead sea bream (Sparus aurata). Aquaculture 2004, 230, 65–80. [Google Scholar] [CrossRef]
- Karaiskou, N.; Triantafyllidis, A.; Katsares, V.; Abatzopoulos, T.J.; Triantaphyllidis, C. Microsatellite variability of wild and farmed populations of Sparus aurata. J. Fish Biol. 2009, 74, 1816–1825. [Google Scholar] [CrossRef]
- De Innocentiis, S.; Lesti, A.; Livi, S.; Rossi, A.R.; Crosetti, D.; Sola, L. Microsatellite markers reveal population structure in gilthead sea bream Sparus auratus from the Atlantic Ocean and Mediterranean Sea. Fish. Sci. 2004, 70, 852–859. [Google Scholar] [CrossRef]
- Šegvić-Bubić, T.; Lepen, I.; Trumbić, Ž.; Ljubković, J.; Sutlović, D.; Matić-Skoko, S.; Grubišić, L.; Glamuzina, B.; Mladineo, I. Population genetic structure of reared and wild gilthead sea bream (Sparus aurata) in the Adriatic Sea inferred with microsatellite loci. Agriculture 2011, 318, 309–315. [Google Scholar] [CrossRef]
- Coscia, I.; Vogiatzi, E.; Kotoulas, G.; Tsigenopoulos, C.S.; Mariani, S. Exploring neutral and adaptive processes in expanding populations of gilthead sea bream, Sparus aurata L., in the North-East Atlantic. Heredity 2012, 108, 537–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Celdrán, M.; Ramis, G.; María-Dolores, E.; Peñalver, J.; Borrell, Y.J.; Manchado, M.; Estévez, A.; Afonso, J.M.; Armero, E. Genetic assessment of three gilthead sea bream (Sparus aurata L.) populations along the Spanish coast and of three broodstocks managements. Aquac. Int. 2016, 24, 1409–1420. [Google Scholar] [CrossRef]
- Loukovitis, D.; Sarropoulou, E.; Vogiatzi, E.; Tsigenopoulos, C.S.; Kotoulas, G.; Magoulas, A.; Chatziplis, D. Genetic variation in farmed populations of the gilthead sea bream Sparus aurata in Greece using microsatellite DNA markers. Aquac. Res. 2011, 42, 239–246. [Google Scholar] [CrossRef]
- Polovina, E.S.; Kourkouni, E.; Tsigenopoulos, C.S.; Sanchez-Jerez, P.; Ladoukakis, E.D. Genetic structuring in farmed and wild Gilthead seabream and European seabass in the Mediterranean Sea: Implementations for detection of escapees. Aquat. Liv. Resour. 2020, 33, 7. [Google Scholar] [CrossRef]
- Šegvić-Bubić, T.; Talijančić, I.; Grubišić, L.; Izquierdo-Gomez, D.; Katavić, I. Morphological and molecular differentiation of wild and farmed gilthead sea bream Sparus aurata: Implications for management. Aquac. Environ. Interact. 2014, 6, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Gkagkavouzis, K.; Papakostas, S.; Maroso, F.; Karaiskou, N.; Carr, A.; Nielsen, E.E.; Bargelloni, L.; Triantafyllidis, A. Investigating Genetic Diversity and Genomic Signatures of Hatchery-Induced Evolution in Gilthead Seabream (Sparus aurata) Populations. Diversity 2021, 13, 563. [Google Scholar] [CrossRef]
- Rossi, A.R.; Miggiano, M.; Franchini, P.; Perrone, E.; Crosetti, D.; Sola, L. Genetic comparison of temporal replicates of gilthead sea bream (Sparus aurata) from two Tyrrhenian coastal lagoons characterized by different management. J. Appl. Ichthyol. 2009, 25, 603–605. [Google Scholar] [CrossRef]
- Maroso, F.; Gkagkavouzis, K.; De Innocentiis, S.; Hillen, J.; do Prado, F.; Karaiskou, N.; Taggart, J.B.; Carr, A.; Nielsen, E.; Triantafyllidis, A.; et al. Genome-wide analysis clarifies the population genetic structure of wild gilthead sea bream (Sparus aurata). PLoS ONE 2021, 16, e0236230. [Google Scholar] [CrossRef]
- Giantsis, I.A.; Mucci, N.; Randi, E.; Abatzopoulos, T.J.; Apostolidis, A.P. Microsatellite variation of mussels (Mytilus galloprovincialis) in central and eastern Mediterranean: Genetic panmixia in the Aegean and the Ionian Seas. J. Mar. Biol. Assoc. U. K. 2014, 94, 797–809. [Google Scholar] [CrossRef]
- Väli, Ü.; Einarsson, A.; Waits, L.; Ellegren, H. To what extent do microsatellite markers reflect genome-wide genetic diversity in natural populations? Mol. Ecol. 2008, 17, 3808–3817. [Google Scholar] [CrossRef]
- Ben Slimen, H.; Guebej, H.; Ben Ohmen, A.; Ould Brahim, I.; Blel, H.; Chatti, N. Genetic differentiation between populations of gilthead seabream (Sparus aurata) along the Tunisian coast. Cybium 2004, 28, 45–50. [Google Scholar]
- Franchini, P.; Sola, L.; Crosetti, D.; Milana, V.; Rossi, A.R. Low levels of population genetic structure in the gilthead sea bream, Sparus aurata, along the coast of Italy. ICES J. Mar. Sci. 2012, 69, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Peñaloza, C.; Robledo, D.; Barría, A.; Trịnh, T.Q.; Mahmuddin, M.; Wiener, P.; Benzie, J.A.; Houston, R.D. Development and validation of an open access SNP array for Nile tilapia (Oreochromis niloticus). G3 Genes Genomes Genet. 2020, 10, 2777–2785. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, B.; Fernández, A.; Peiró-Pastor, R.; Penaloza, C.; Houston, R.D.; Sonesson, A.K.; Tsigenopoulos, C.S.; Bargelloni, L.; Gamsız, K.; Karahan, B.; et al. Population structure and genetic variability in wild and farmed Mediterranean populations of gilthead seabream and European seabass inferred from a 60K combined species SNP array. Aquac. Rep. 2022, 24, 101145. [Google Scholar] [CrossRef]
- Allegrucci, G.; Fortunato, C.; Sbordoni, V. Genetic structure and allozyme variation of sea bass (Dicentrarchus labrax and D. punctatus) in the Mediterranean Sea. Mar. Biol. 1997, 128, 347–358. [Google Scholar] [CrossRef]
- Caccone, A.; Allegrucci, G.; Fortunato, C.; Sbordoni, V. Genetic differentiation within the European sea bass (D. labrax) as revealed by RAPD-PCR assays. J. Hered. 1997, 88, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Garcia de Leon, F.J.; Chikhi, L.; Bonhomme, F. Microsatellite polymorphism and population subdivision in natural populations of European sea bass Dicentrarchus labrax (Linnaeus, 1758). Mol. Ecol. 1997, 6, 51–62. [Google Scholar] [CrossRef]
- Naciri, M.; Lemaire, C.; Borsa, P.; Bonhomme, F. Genetic study of the Atlantic/Mediterranean transition in sea bass (Dicentrarchus labrax). J. Hered. 1999, 90, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Bahri-Sfar, L.; Lemaire, C.; Hassine, O.K.B.; Bonhomme, F. Fragmentation of sea bass populations in the western and eastern Mediterranean as revealed by microsatellite polymorphism. Proc. R. Soc. B Biol. Sci. 2000, 267, 929–935. [Google Scholar] [CrossRef]
- Souche, E.L.; Hellemans, B.; Babbucci, M.; MacAoidh, E.; Guinand, B.; Bargelloni, L.; Chistiakov, D.A.; Patarnello, T.; Bonhomme, F.; Martinsohn, J.T.; et al. Range-wide population structure of European sea bass Dicentrarchus labrax. Biol. J. Lin. Soc. 2015, 116, 86–105. [Google Scholar] [CrossRef] [Green Version]
- Bodur, T.; Tsigenopoulos, C.; Cagatay, I.T. Genetic structure of wild european sea bass (Dicentrarchus labrax L, 1758) populations in Aegean and Levantine Sea using microsatellite markers. Tur. J. Fish. Aquat. Sci. 2017, 17, 7–14. [Google Scholar]
- Brown, C.; Miltiadou, D.; Tsigenopoulos, C.S. Prevalence and survival of escaped European seabass Dicentrarchus labrax in Cyprus identified using genetic markers. Aquac. Environ. Interac. 2015, 7, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Brokordt, K.B.; Fernández, M.; Gaymer, C.F. Domestication reduces the capacity to escape from predators. J. Exp. Mar. Biol. Ecol. 2006, 329, 11–19. [Google Scholar] [CrossRef]
- Toledo Guedes, K.; Sánchez-Jerez, P.; González-Lorenzo, G.; Brito Hernández, A. Detecting the degree of establishment of a non-indigenous species in coastal ecosystems: Sea bass Dicentrarchus labrax escapes from sea cages in Canary Islands (Northeastern Central Atlantic). Hydrobiology. 2009, 623, 203–212. [Google Scholar] [CrossRef]
- Toledo-Guedes, K.; Sanchez-Jerez, P.; Mora-Vidal, J.; Girard, D.; Brito, A. Escaped introduced sea bass (Dicentrarchus labrax) infected by Sphaerospora testicularis (Myxozoa) reach maturity in coastal habitats off Canary Islands. Mar. Ecol. 2012, 33, 26–31. [Google Scholar] [CrossRef]
- Valero-Rodriguez, J.M.; Toledo-Guedes, K.; Arechavala-Lopez, P.; Izquierdo-Gomez, D.; Sanchez-Jerez, P. The use of trophic resources by Argyrosomus regius (Asso, 1801) escaped from Mediterranean offshore fish farms. J. Appl. Ichthyol. 2015, 31, 10–15. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Uglem, I.; Izquierdo-Gomez, D.; Fernandez-Jover, D.; Sanchez-Jerez, P. Rapid dispersion of escaped meagre (Argyrosomus regius) from a coastal Mediterranean fish farm. Aquac. Res. 2017, 48, 1502–1512. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Izquierdo-Gomez, D.; Sanchez-Jerez, P.; Bayle-Sempere, J.T. Simulating escapes of farmed sea bass from Mediterranean open sea-cages: Low recaptures by local fishermen. J. Appl. Ichthyol. 2014, 30, 185–188. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Uglem, I.; Fernandez-Jover, D.; Bayle-Sempere, J.T.; Sanchez-Jerez, P. Immediate post-escape behaviour of farmed seabass (Dicentrarchus labrax L.) in the Mediterranean Sea. J. Appl. Ichthyol. 2011, 27, 1375–1378. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Uglem, I.; Fernandez-Jover, D.; Bayle-Sempere, J.T.; Sanchez-Jerez, P. Post-escape dispersion of farmed seabream (Sparus aurata L.) and recaptures by local fisheries in the Western Mediterranean Sea. Fish. Res. 2012, 121, 126–135. [Google Scholar] [CrossRef]
- Šegvić-Bubić, T.; Arechavala-Lopez, P.; Vučić, I.; Talijančić, I.; Grubišić, L.; Žužul, I.; Kovač, Ž. Site fidelity of farmed gilthead seabream Sparus aurata escapees in a coastal environment of the Adriatic Sea. Aquac. Environ. Interac. 2018, 10, 21–34. [Google Scholar] [CrossRef]
- Izquierdo-Gómez, D.; Arechavala-Lopez, P.; Bayle-Sempere, J.T.; Sánchez-Jerez, P. Assessing the influence of gilthead sea bream escapees in landings of Mediterranean fisheries through a scale-based methodology. Fish. Manag. Ecol. 2017, 24, 62–72. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Toledo-Guedes, K.; Izquierdo-Gomez, D.; Šegvić-Bubić, T.; Sanchez-Jerez, P. Implications of sea bream and sea bass escapes for sustainable aquaculture management: A review of interactions, risks and consequences. Rev. Fish. Sci. Aquac. 2018, 26, 214–234. [Google Scholar] [CrossRef]
- Kalantzi, I.; Zeri, C.; Catsiki, V.A.; Tsangaris, C.; Strogyloudi, E.; Kaberi, H.; Vergopoulos, N.; Tsapakis, M. Assessment of the use of copper alloy aquaculture nets: Potential impacts on the marine environment and on the farmed fish. Aquaculture 2016, 465, 209–222. [Google Scholar] [CrossRef]
- Berillis, P.; Mente, E.; Kormas, K.A. The use of copper alloy in aquaculture fish net pens: Mechanical, economic and environmental advantages. J. Fish. 2017, 11, 1–3. [Google Scholar] [CrossRef]
- Dempster, T.; Arechavala-Lopez, P.; Barrett, L.T.; Fleming, I.A.; Sanchez-Jerez, P.; Uglem, I. Recapturing escaped fish from marine aquaculture is largely unsuccessful: Alternatives to reduce the number of escapees in the wild. Rev. Aquac. 2018, 10, 153–167. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Waples, R.S.; Baskett, M.L. Life history and temporal variability of escape events interactively determine the fitness consequences of aquaculture escapees on wild populations. Theor. Popul. Biol. 2019, 129, 93–102. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Cabrera-Álvarez, M.J.; Maia, C.M.; Saraiva, J.L. Environmental enrichment in fish aquaculture: A review of fundamental and practical aspects. Rev. Aquac. 2022, 14, 704–728. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
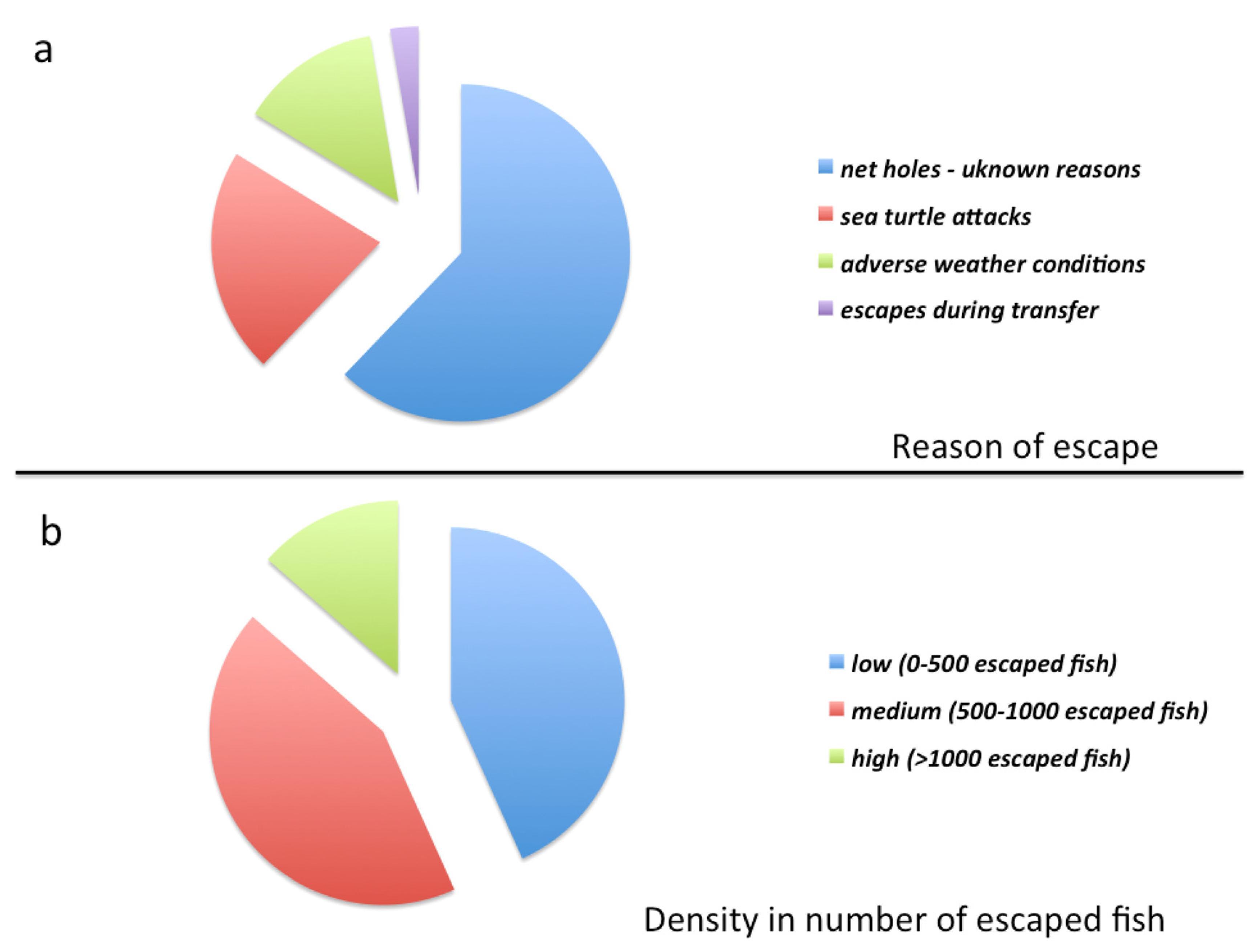{kind=link}
| Major Cause Category | Etiological Factor for the Escape Event |
|---|---|
| Environmental |
|
| Organizational failures |
|
| Operational risks that may cause damages such as holes in fish cages |
|
| Technical |
|
| Location of Populations | FST | Markers Used | Reference |
|---|---|---|---|
| Central Mediterranean | 0.093 | Allozymes | Ben Slimen et al. [63] |
| Western Mediterranean | 0.069 | Microsatellites—RAPDs | Chaoui et al. [47] |
| Central Mediterranean | 0.017 | Microsatellites | Franchini et al. [64] |
| Italian coasts | 0.007 | Microsatellites | Franchini et al. [64] |
| Greece (Aegean vs. Ionian Sea) | 0.002 | Microsatellites | Gkagkavouzis et al. [48] |
| Adriatic | 0.033 | Microsatellites | Šegvić-Bubić et al. [52] |
| Atlantic | 0.068 | Microsatellites—EST—mtDNA dloop | Coscia et al. [53] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvanou, M.V.; Gkagkavouzis, K.; Karaiskou, N.; Feidantsis, K.; Lattos, A.; Michaelidis, B.; Theodorou, J.A.; Batargias, C.; Triantafyllidis, A.; Giantsis, I.A. Mediterranean Aquaculture and Genetic Pollution: A Review Combined with Data from a Fish Farm Evaluating the Ecological Risks of Finfish Escapes. J. Mar. Sci. Eng. 2023, 11, 1405. https://doi.org/10.3390/jmse11071405
Alvanou MV, Gkagkavouzis K, Karaiskou N, Feidantsis K, Lattos A, Michaelidis B, Theodorou JA, Batargias C, Triantafyllidis A, Giantsis IA. Mediterranean Aquaculture and Genetic Pollution: A Review Combined with Data from a Fish Farm Evaluating the Ecological Risks of Finfish Escapes. Journal of Marine Science and Engineering. 2023; 11(7):1405. https://doi.org/10.3390/jmse11071405
Chicago/Turabian StyleAlvanou, Maria V., Konstantinos Gkagkavouzis, Nikoleta Karaiskou, Konstantinos Feidantsis, Athanasios Lattos, Basile Michaelidis, John A. Theodorou, Costas Batargias, Alexandros Triantafyllidis, and Ioannis A. Giantsis. 2023. "Mediterranean Aquaculture and Genetic Pollution: A Review Combined with Data from a Fish Farm Evaluating the Ecological Risks of Finfish Escapes" Journal of Marine Science and Engineering 11, no. 7: 1405. https://doi.org/10.3390/jmse11071405
APA StyleAlvanou, M. V., Gkagkavouzis, K., Karaiskou, N., Feidantsis, K., Lattos, A., Michaelidis, B., Theodorou, J. A., Batargias, C., Triantafyllidis, A., & Giantsis, I. A. (2023). Mediterranean Aquaculture and Genetic Pollution: A Review Combined with Data from a Fish Farm Evaluating the Ecological Risks of Finfish Escapes. Journal of Marine Science and Engineering, 11(7), 1405. https://doi.org/10.3390/jmse11071405