
Elucidating Temporal Patterns in Coral Health and Assemblage Structure in Papahānaumokuākea

Abstract
:1. Introduction
2. Materials and Methods
2.1. Survey Design
2.2. Coral Assemblage Analyses
2.3. Coral Health Condition Analyses
3. Results
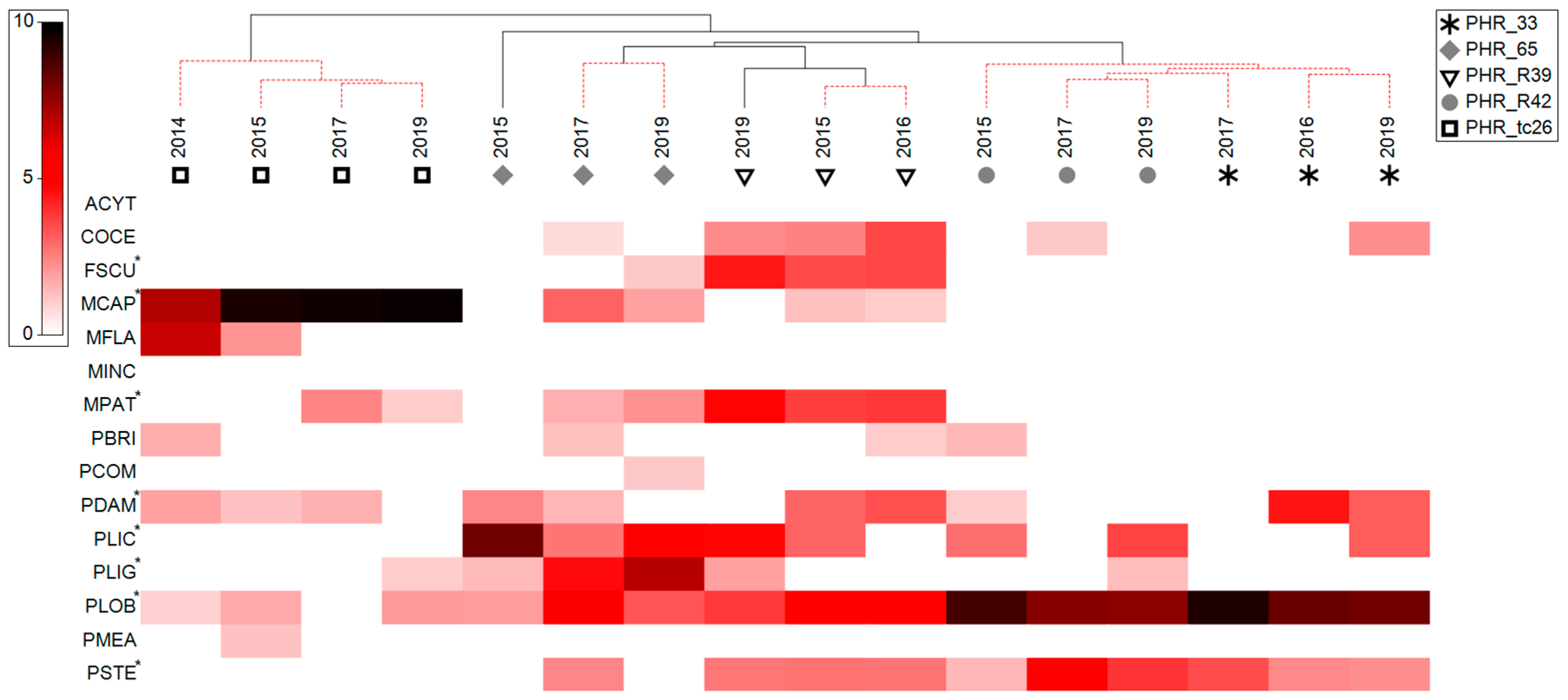
3.1. Coral Assemblage Composition—Percent Colony Count
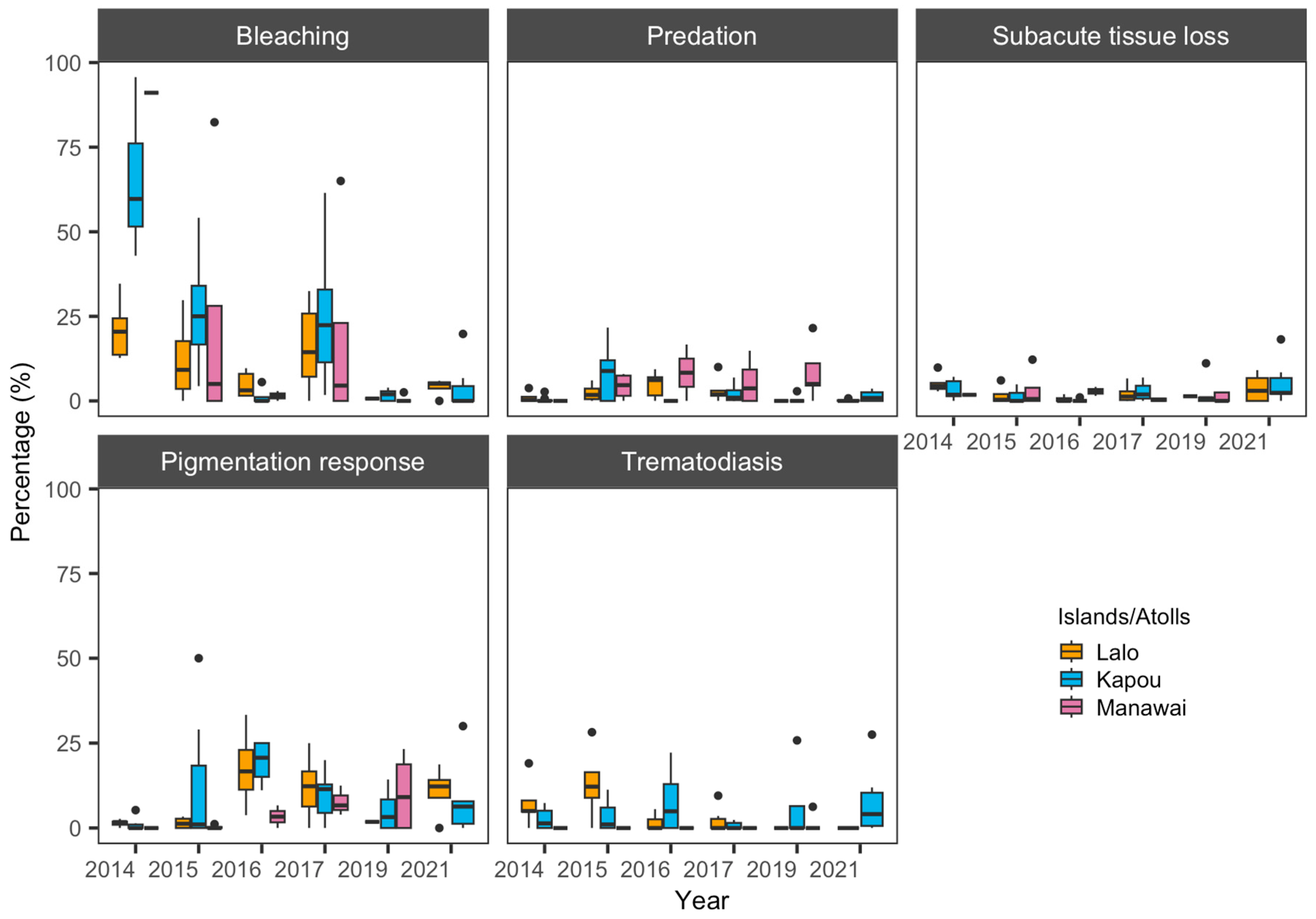
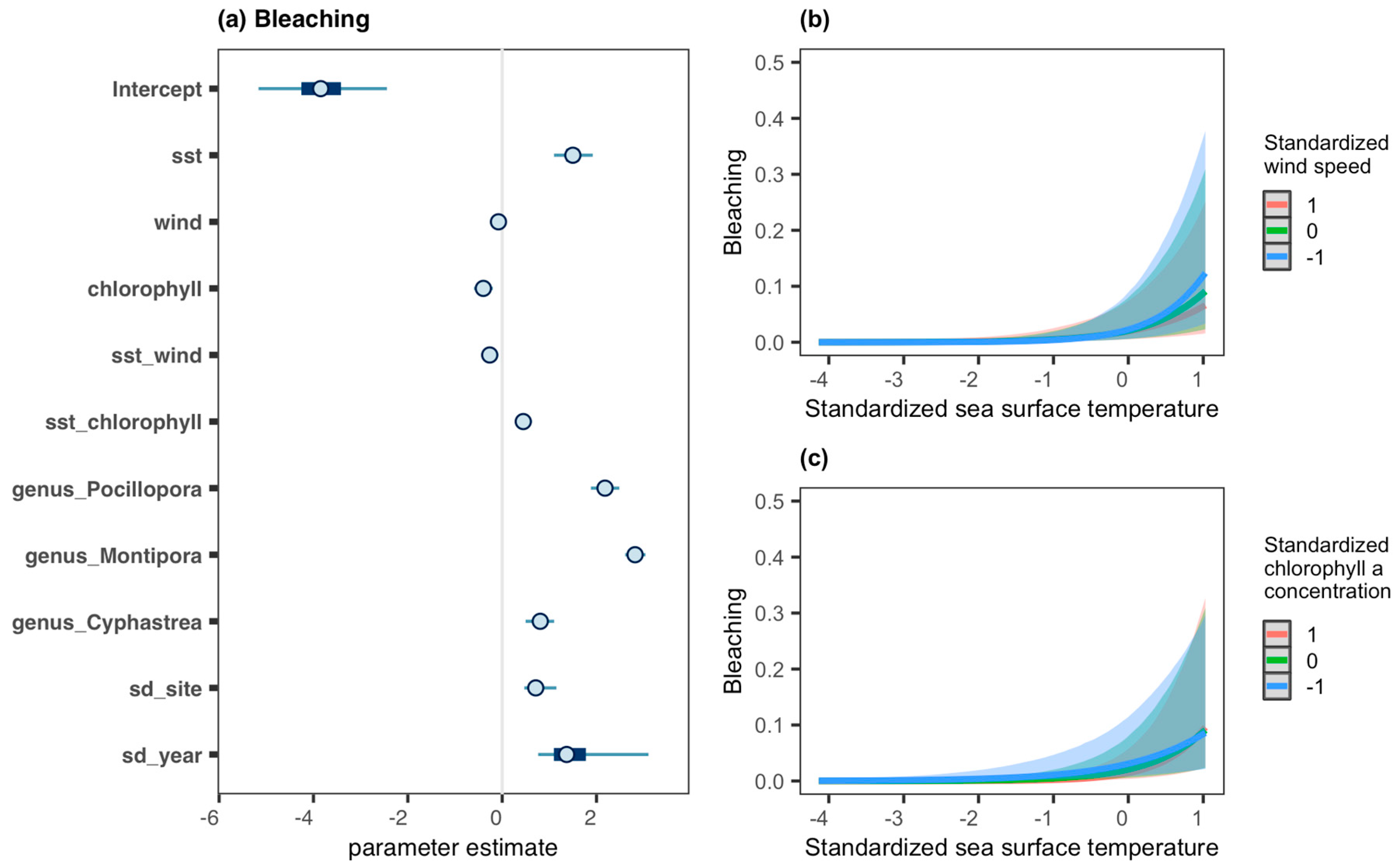
3.2. Coral Health Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Quataert, E.; Storlazzi, C.; van Rooijen, A.; Cheriton, O.; van Dongeren, A. The influence of coral reefs and climate change on wave-driven flooding of tropical coastlines. Geophys. Res. Lett. 2015, 42, 6407–6415. [Google Scholar] [CrossRef]
- Perry, C.T.; Edinger, E.N.; Kench, P.S.; Murphy, G.N.; Smithers, S.G.; Steneck, R.S.; Mumby, P.J. Estimating rates of biologically driven coral reef framework production and erosion: A new census-based carbonate budget methodology and applications to the reefs of Bonaire. Coral Reefs 2012, 31, 853–868. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Pendleton, L.; Kaup, A. People and the changing nature of coral reefs. Reg. Stud. Mar. Sci. 2019, 30, 100699. [Google Scholar] [CrossRef]
- Moritz, C.; Brandl, S.J.; Rouzé, H.; Vii, J.; Pérez-Rosales, G.; Bosserelle, P.; Chancerelle, Y.; Galzin, R.; Liao, V.; Siu, G.; et al. Long-term monitoring of benthic communities reveals spatial determinants of disturbance and recovery dynamics on coral reefs. Mar. Ecol. Prog. Ser. 2021, 672, 141–152. [Google Scholar] [CrossRef]
- Couch, C.S.; Burns, J.H.R.; Liu, G.; Steward, K.; Gutlay, T.N.; Kenyon, J.; Eakin, C.M.; Kosaki, R.K. Mass coral bleaching due to unprecedented marine heatwave in Papahānaumokuākea Marine National Monument (Northwestern Hawaiian Islands). PLoS ONE 2017, 12, e0185121. [Google Scholar] [CrossRef]
- Pascoe, K.H.; Fukunaga, A.; Kosaki, R.K.; Burns, J.H.R. 3D assessment of a coral reef at Lalo Atoll reveals varying responses of habitat metrics following a catastrophic hurricane. Sci. Rep. 2021, 11, 12050. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.F.; Petes, L.E.; Harvell, C.D.; Hettinger, A. Nutrient enrichment can increase the severity of coral diseases. Ecol. Lett. 2003, 6, 1056–1061. [Google Scholar] [CrossRef]
- Walton, C.J.; Hayes, N.K.; Gilliam, D.S. Impacts of a regional, multi-year, multi-species coral disease outbreak in southeast Florida. Front. Mar. Sci. 2018, 5, 323. [Google Scholar] [CrossRef]
- Burke, S.; Pottier, P.; Lagisz, M.; Macartney, E.L.; Ainsworth, T.; Drobniak, S.M.; Nakagawa, S. The impact of rising temperatures on the prevalence of coral diseases and its predictability: A global meta-analysis. Ecol. Lett. 2023, 26, 1466–1481. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Filip, L.; Estrada-Saldívar, N.; Pérez-Cervantes, E.; Molina-Hernández, A.; González-Barrios, F.J. A rapid spread of the stony coral tissue loss disease outbreak in the Mexican Caribbean. PeerJ 2019, 7, e8069. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, A.; Pascoe, K.H.; Pugh, A.R.; Kosaki, R.K.; Burns, J.H.R. Underwater photogrammetry captures the initial recovery of a coral reef at Lalo Atoll. Diversity 2022, 14, 39. [Google Scholar] [CrossRef]
- Sherwood, A.R.; Huisman, J.M.; Paiano, M.O.; Williams, T.M.; Kosaki, R.K.; Smith, C.M.; Giuseffi, L.; Spalding, H.L. Taxonomic determination of the cryptogenic red alga, Chondria tumulosa sp. nov., (Rhodomelaceae, Rhodophyta) from Papahānaumokuākea Marine National Monument, Hawai‘i, USA: A new species displaying invasive characteristics. PLoS ONE 2022, 15, e0234358. [Google Scholar] [CrossRef] [PubMed]
- Bythell, J.C.; Bythell, M.; Gladfelter, E.H. Initial results of a long-term coral reef monitoring program: Impact of Hurricane Hugo at Buck Island Reef National Monument, St. Croix, US. Virgin Islands. J. Exp. Mar. Biol. Ecol. 1993, 172, 171–183. [Google Scholar] [CrossRef]
- Adjeroud, M.; Chancerelle, Y.; Schrimm, M.; Perez, T.; Lecchini, D.; Galzin, R.; Salvat, B. Detecting the effects of natural disturbances on coral assemblages in French Polynesia: A decade survey at multiple scales. Aquat. Living Resour. 2005, 18, 111–123. [Google Scholar] [CrossRef]
- Obura, D.; Gudka, M.; Samoilys, M.; Osuka, K.; Mbugua, J.; Keith, D.A.; Porter, S.; Roche, R.; van Hooidonk, R.; Ahamada, S.; et al. Vulnerability to collapse of coral reef ecosystems in the Western Indian Ocean. Nat. Sustain. 2022, 5, 104–113. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Clarke, K.R.; Somerfield, P.J.; Gorley, R.N. Testing of null hypotheses in exploratory community analyses: Similarity profiles and biota-environment linkage. J. Exp. Mar. Biol. Ecol. 2008, 366, 56–69. [Google Scholar] [CrossRef]
- Somerfield, P.J.; Clarke, K.R. Inverse analysis in non-parametric multivariate analyses: Distinguishing groups of associated species which covary coherently across samples. J. Exp. Mar. Biol. Ecol. 2013, 449, 261–273. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Bürkner, P.-C. Brms: An R package for Bayesian multilevel models using Stan. J. Stat. Softw. 2017, 80, 1–28. [Google Scholar] [CrossRef]
- Bürkner, P.-C. Advanced Bayesian multilevel modeling with the R package brms. R J. 2018, 10, 395–411. [Google Scholar] [CrossRef]
- Bürkner, P.-C. Bayesian item response modeling in R with brms and Stan. J. Stat. Softw. 2021, 100, 1–54. [Google Scholar] [CrossRef]
- Eddelbuettel, D.; François, R. Rcpp: Seamless R and C++ integration. J. Stat. Softw. 2011, 40, 1–18. [Google Scholar] [CrossRef]
- Eddelbuettel, D. Seamless R and C++ Integration with Rcpp; Springer: New York, NY, USA, 2013; p. 220. [Google Scholar]
- Eddelbuettel, D.; Balamuta, J.J. Extending R with C++: A brief introduction to Rcpp. Am. Stat. 2018, 72, 28–36. [Google Scholar] [CrossRef]
- Eakin, C.M.; Sweatman, H.P.A.; Brainard, R.E. The 2014–2017 global-scale coral bleaching event: Insights and impacts. Coral Reefs 2019, 38, 539–545. [Google Scholar] [CrossRef]
- Fukunaga, A.; Burns, J.H.R.; Pascoe, K.H.; Kosaki, R.K. A remote coral reef shows macroalgal succession following a mass bleaching event. Ecol. Indic. 2022, 142, 109175. [Google Scholar] [CrossRef]
- Paparella, F.; Xu, C.; Vaughan, G.O.; Burt, J.A. Coral Bleaching in the Persian/Arabian Gulf is modulated by summer winds. Front. Mar. Sci. 2019, 6, 205. [Google Scholar] [CrossRef]
- Kenyon, J.C.; Aeby, G.S.; Brainard, R.E.; Chojnacki, J.D.; Dunlap, M.J.; Wilkinson, C.B. Mass coral bleaching on high-latitude reef in the Hawaiian Archipelago. In Proceedings of the 10th International Coral Reef Symposium, Okinawa, Japan, 28 June–3 July 2006; Japanese Coral Reef Society: Okinawa, Japan, 2006; pp. 631–643. [Google Scholar]
- Bahr, K.D.; Jokiel, P.L.; Rodgers, K.S. Relative sensitivity of five Hawaiian coral species to high temperature under high-pCO2 conditions. Coral Reefs 2016, 35, 729–738. [Google Scholar] [CrossRef]
- Grottoli, A.G.; Rodrigues, L.J.; Palardy, J.E. Heterotrophic plasticity and resilience in bleached corals. Nature 2006, 440, 1186–1189. [Google Scholar] [CrossRef] [PubMed]
- Tricas, T.C. Prey selection by coral-feeding butterflyfishes: Strategies to maximize the profit. Environ. Biol. Fish. 1989, 25, 171–185. [Google Scholar] [CrossRef]
- Jayewardene, D.; Donahue, M.J.; Birkeland, C. Effects of frequent fish predation on corals in Hawaii. Coral Reefs 2009, 28, 499–506. [Google Scholar] [CrossRef]
- Maragos, J.E.; Potts, D.C.; Aeby, G.S.; Gulko, D.; Kenyon, J.; Siciliano, D.; VanRavenswaay, D. 2000–2002 Rapid Ecological Assessment of corals (Anthozoa) on shallow reefs of the Northwestern Hawaiian Islands. Part 1: Species and distribution. Pac. Sci. 2004, 58, 211–230. [Google Scholar] [CrossRef]
- D’Angelo, C.; Smith, E.G.; Oswald, F.; Burt, J.; Tchernov, D.; Wiedenmann, J. Locally accelerated growth is part of the innate immune response and repair mechanisms in reef-building corals as detected by green fluorescent protein (GFP)-like pigments. Coral Reefs 2012, 31, 1045–1056. [Google Scholar] [CrossRef]
- Palmer, C.V.; Roth, M.S.; Gates, R.D. Red fluorescent protein responsible for pigmentation in trematode-infected Porites compressa tissues. Biol. Bull. 2009, 216, 68–74. [Google Scholar] [CrossRef]
- Kubomura, T.; Wee, H.B.; Reimer, J.D. Investigating incidence and possible causes of pink and purple pigmentation response in hard coral genus Porites around Okinawajima Island, Japan. Reg. Stud. Mar. Sci. 2021, 41, 101569. [Google Scholar] [CrossRef]
- Benzoni, F.; Galli, P.; Pichon, M. Pink spots on Porites: Not always a coral disease. Coral Reefs 2010, 29, 153. [Google Scholar] [CrossRef]
- Ravindran, J.; Raghukumar, C. Pink line syndrome (PLS) in the scleractinian coral Porites lutea. Coral Reefs 2002, 21, 252. [Google Scholar] [CrossRef]
- Bruno, J.F.; Selig, E.R.; Casey, K.S.; Page, C.A.; Willis, B.L.; Harvell, C.D.; Sweatman, H.; Melendy, A.M. Thermal stress and coral cover as drivers of coral disease outbreaks. PLoS Biol. 2007, 5, e124. [Google Scholar] [CrossRef]
- Kuta, K.; Richardson, L. Ecological aspects of black band disease of corals: Relationships between disease incidence and environmental factors. Coral Reefs 2002, 21, 393–398. [Google Scholar] [CrossRef]
- Rooney, J.J.; Wessel, P.; Hoeke, R.; Weiss, J.; Baker, J.; Parrish, F.; Fletcher, C.H.; Chojnacki, J.; Garcia, M.; Brainard, R.; et al. Geology and geomorphology of coral reefs in the Northwestern Hawaiian Islands. In Coral Reefs of the USA; Riegl, B.M., Dodge, R.E., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 519–571. [Google Scholar]
- Aeby, G.S.; Williams, G.J.; Franklin, E.C.; Kenyon, J.; Cox, E.F.; Coles, S.; Work, T.M. Patterns of coral disease across the Hawaiian Archipelago: Relating disease to environment. PLoS ONE 2011, 6, e20370. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Depths (m) | Pins | 2014 | 2015 | 2016 | 2017 | 2019 | 2021 |
| FFS 30 | 6–8 | x | x | x | x | x | ||
| FFS 32 | 6–8 | x | x | x | x | |||
| FFS 33 | 9–10 | x | x | x | x | x | ||
| FFS 34 | 9–10 | x | x | x | x | x | ||
| FFS R29 | 10–11 | x | x | x | x | |||
| FFS Rapture Reef | 23–26 | x | x | x | ||||
| LIS 10 | 8–9 | x | x | x | x | x | ||
| LIS 12 | 7–10 | x | x | x | x | |||
| LIS 18 | 7–9 | x | x | x | x | x | ||
| LIS 4008 | 7–8 | ? | x | x | x | x | ||
| LIS 4019 | 14–17 | ? | x | x | x | x | ||
| LIS 4067 | 1–5 | x | x | x | x | x | x | |
| LIS Court | 4–7 | x | x | x | x | x | x | |
| LIS R7 | 10–11 | x | x | x | ||||
| LIS R10 | 12–14 | x | x | x | x | |||
| PHR 33 | 12 | x | x | x | ||||
| PHR 65 | 13–21 | ? | x | x | x | |||
| PHR R39 | 11–14 | x | x | x | ||||
| PHR R42 | 14–15 | x | x | x | ||||
| PHR tc26 | 2–3 | x | x | x | x |
| Source | df | SS | MS | Pseudo-F | P |
|---|---|---|---|---|---|
| Region | 2 | 26,975 | 13,487 | 2.90 | 0.0050 |
| Year | 5 | 8470 | 1694 | 3.05 | 0.0002 |
| Site (Region) | 17 | 97,132 | 5714 | 10.29 | 0.0002 |
| Region:Year | 9 | 9605 | 1067 | 1.92 | 0.0026 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukunaga, A.; Pascoe, K.H.; Kosaki, R.K.; Burns, J.H.R. Elucidating Temporal Patterns in Coral Health and Assemblage Structure in Papahānaumokuākea. J. Mar. Sci. Eng. 2024, 12, 1267. https://doi.org/10.3390/jmse12081267
Fukunaga A, Pascoe KH, Kosaki RK, Burns JHR. Elucidating Temporal Patterns in Coral Health and Assemblage Structure in Papahānaumokuākea. Journal of Marine Science and Engineering. 2024; 12(8):1267. https://doi.org/10.3390/jmse12081267
Chicago/Turabian StyleFukunaga, Atsuko, Kailey H. Pascoe, Randall K. Kosaki, and John H. R. Burns. 2024. "Elucidating Temporal Patterns in Coral Health and Assemblage Structure in Papahānaumokuākea" Journal of Marine Science and Engineering 12, no. 8: 1267. https://doi.org/10.3390/jmse12081267