
Ontogenetic Variation in the Trophic and Mercury Levels of Japanese Anchovy in the High Seas of the Northwestern Pacific Ocean

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. Stable Isotope Analysis
2.3. Analysis of Hg Levels
2.4. Data Analysis
3. Results
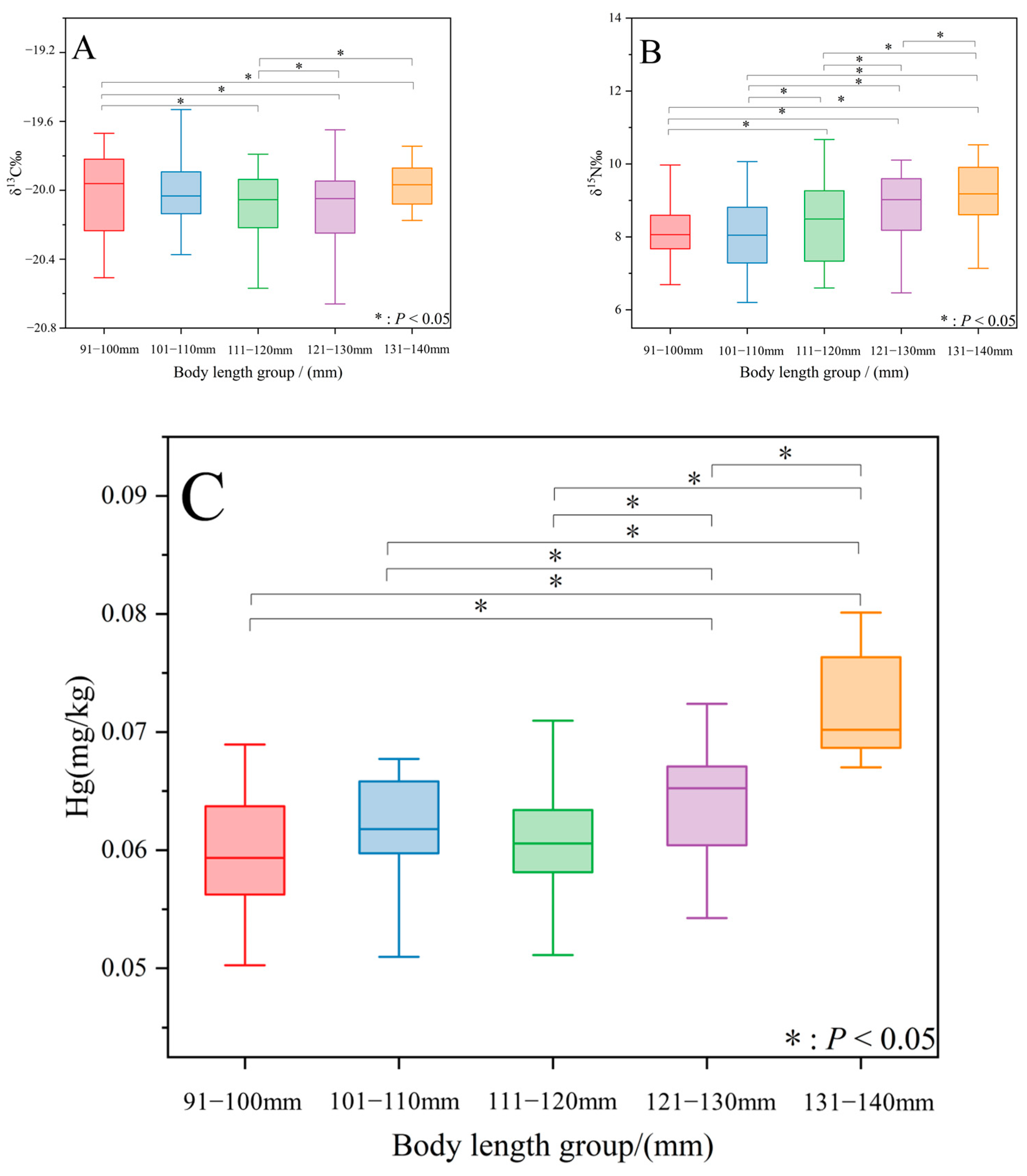
3.1. Differences in Isotopic Values and Hg Levels among Body Length Groups
3.2. Trophic Levels and Trophic Niche
3.3. Correlations between δ15N and δ13C Stable Isotopes and Hg Levels
4. Discussion
4.1. Differences in Isotope Values among Body Length Groups
4.2. Trophic Levels and Trophic Niches
4.3. Factors Explaining the Variation in Hg Levels
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yoshino, K.; Mori, K.; Kanaya, G.; Kojima, S.; Henmi, Y.; Matsuyama, A.; Yamamoto, M. Food sources are more important than biomagnification on mercury bioaccumulation in marine fishes. Environ. Pollut. 2020, 262, 113982. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.Y. A Master’s Thesis on the Process of Small Zooplankton Feeding Rates in Response to Mesoscale Cyclonic Eddies in the Northwestern Pacific Ocean. Master’s Thesis, Xiamen University, Xiamen, China, 2021. Available online: https://link.cnki.net/doi/10.27424/d.cnki.gxmdu.2021.002352 (accessed on 17 June 2024).
- Clayton, S.; Nagai, T.; Follows, M.J. Fine scale phytoplankton community structure across the Kuroshio Front. J. Plankton Res. 2014, 36, 1017–1030. [Google Scholar] [CrossRef]
- Liu, J.X.; Gao, T.X.; Zhuang, Z.M.; Yokogawa, K.; Zhang, Y.P. Late Pleist-ocene divergence and subsequent population expansion of two closely related fish spe-cies, Japanese anchovy (Engraulis japonicus) and Australian anchovy (Engraulis austr-alis). Mol. Phylogenetics Evol. 2006, 40, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Landry, M.R.; Brown, S.L.; Rii, Y.M.; Selph, K.E.; Bidigare, R.R.; Yang, E.J.; Simmons, M.P. Depth-stratified phytoplankton dynamics in Cyclone Opal, a subtropical mesoscale eddy. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 1348–1359. [Google Scholar] [CrossRef]
- Cossa, D.; Coquery, M.; Gobeil, C.; Martin, J.M. Mercury Fluxes at the Ocean Margins. In Global and Regional Mercury Cycles: Sources Fluxes and Mass Balances; 1996; pp. 229–247. Available online: https://www.usualwant.com/chapter/10 (accessed on 5 January 1997).
- Meng, T.X. Studies on the feeding of Japanese anchovy (Engraulis japonicus) post larva in the spawning ground in the Southern waters of Shandongpeninsula. Mar. Fish. Res. 2001, 22, 21–25. [Google Scholar]
- AMAP Assessment. Hg in the Arctic; Arctic Monitoring and Assessment Programme (AMAP): Oslo, Norway, 2011; 193p. [Google Scholar]
- FAO. FAO Yearbook. In Fishery and Aquaculture Statistics 2018/FAO Annuaire; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Streets, D.G.; Devane, M.K.; Lu, Z.; Bond, T.C. All-time releases of mercury to the atmosphere from human activities. Environ. Sci. Technol. 2011, 45, 10485–10491. [Google Scholar] [CrossRef] [PubMed]
- Morel, F.M.; Kraepiel, A.M.; Amyot, M. The chemical cycle and bioaccumulation of mercury. Annu. Rev. Ecol. Syst. 1998, 29, 543–566. [Google Scholar] [CrossRef]
- Takahashi, M.; Watanabe, Y.; Kinoshita, T.; Watanabe, C. Growth of larval and early juvenile Japanese anchovy, Engraulis japonicus, in the Kuroshio Oyashio transition region. Fish. Oceanogr. 2001, 10, 235–247. [Google Scholar] [CrossRef]
- Le Croizier, G.; Schaal, G.; Point, D.; Le Loc’h, F.; Machu, E.; Fall, M. Stable isotope analyses revealed the influence of foraging habitat on mercury accumulation in tropical coastal marine fish. Sci. Total Environ. 2019, 650, 2129–2140. [Google Scholar] [CrossRef] [PubMed]
- McKinney, M.A.; Dean, K.; Hussey, N.E.; Cliff, G.; Wintner, P.; Dudley, S.F. Global versus local causes and health implications of high mercury concentrations in sharks from the east coast of South Africa. Sci. Total Environ. 2016, 541, 176–183. [Google Scholar] [CrossRef]
- Mintzer, V.J.; Gannon, D.P.; Barros, N.B.; Read, A.J. Stomach contents of massstranded short-finned pilot whales (Globicephala macrorhynchus) from North Carolina. Mar. Mammal. Sci. 2008, 24, 290–302. [Google Scholar] [CrossRef]
- Perissinotto, R.; Pakhomov, E.A. Feeding association of the copepod Rhincalanus gigas with the tunicate salp Salpa thompsoni in the southern ocean. Mar. Biol. 1997, 127, 479–483. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, R.J.; Hu, C.L. Feeding ecology of anchovy based on stomach contents analysis and stable isotope techniques. J. Appl. Ecol. 2021, 2035–2044. [Google Scholar]
- Storelli, M.M.; Barone, G.; Piscitelli, G.; Marcotrigiano, G.O. Mercury in fish: Concentration vs. fish size and estimates of mercury intake. Food Addit. Contam. 2007, 24, 1353–1357. [Google Scholar] [CrossRef] [PubMed]
- Phillips, K.L.; Nichols, P.D.; Jackson, G.D. Lipid and fatty acid composition of the mantle and digestive gland of four Southern Ocean squid species: Implications for food-web studies. Antarct. Sci. 2002, 14, 212–220. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Pethybridge, H.R.; Wu, F.; Li, Y. Mercury bioaccumulation in thresher sharks from the eastern tropical Pacific: Influences of body size, maturation stage, and feeding habitat. Sci. Total Environ. 2023, 872, 162248. [Google Scholar] [CrossRef] [PubMed]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Hoboken, NJ, USA, 1999. [Google Scholar]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche-widths among and within communities: SIBER–Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 3.5.0; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Zhang, J.; Liu, B.; Hu, S.; Gong, Y. Trophic structure of micronekton in the Northwest Pacific Ocean. Front. Mar. Sci. 2022, 9, 971649. [Google Scholar] [CrossRef]
- Aita, M.N.; Tadokoro, K.; Ogawa, N.O.; Hyodo, F.; Ishii, R.; Smith, S.L.; Saino, T.; Kishi, M.J.; Saitoh, S.-I.; Wada, E. Linear relationship between carbon and nitrogen isotope ratios along simple food chains in marine environments. J. Plankton Res. 2011, 33, 1629–1642. [Google Scholar] [CrossRef]
- Gorbatenko, K.M.; Lazhentsev, A.E.; Kiyashko, S.I. Seasonal dynamics of the trophic status of zooplankton in the Sea of Okhotsk (based on data from stable carbon and nitrogenisotope analysis). Russ. J. Mar. Biol. 2014, 40, 519–531. [Google Scholar] [CrossRef]
- Zhang, B.; Yan, W.; Dai, W.F. Application of stable isotope techniques to study the feeding ecology of summer fish communities in Laoshan Bay. J. Aquat. Fish. 2016, 40, 585–594. [Google Scholar]
- DeNiro, M.J.; Epstein, S. Influence of diet on the distribution of nitrogen isotopes in animals. Geochim. Cosmochim. Acta 1981, 45, 341–351. [Google Scholar] [CrossRef]
- Cui, P.D.; Zhang, Y.X. Distribution characteristics of anchovy spawning grounds and their influencing factors in the offshore waters of Liaoning in the northern Yellow Sea. Adv. Fish. Sci. 2023, 31–45. [Google Scholar]
- Akahashi, K.; Wada, E.; Sakamoto, M. Relationship between Carbon Isotope Discrimination and the Specific Growth Rate of Green Alga Chlamydomonas reinhardtii. Jpn. J. Limnol. 1991, 52, 105–112. [Google Scholar] [CrossRef]
- Yang, N.; Hua, C.X.; Zhu, Q.C. Carbon and nitrogen stable isotope characterization of fallfish from the high seas of the Northwest Pacific Ocean. Ocean. Lakes 2023, 54, 412–423. [Google Scholar]
- Tanaka, H.; Aoki, I.; Ohshimo, S. Feeding habit and gill raker morphology of three planktivorous pelagic fish species off the coast of northern and western Kyushu in summer. J. Fish Biol. 2006, 68, 1041–1061. [Google Scholar] [CrossRef]
- Meng, T.X. Studies on the feeding of Japanese anchovy (Engraulis japonicus) at different life stages on zooplankton in the Middle and Southern Waters ofthe Yellow Sea. Mar. Fish. Res. 2003, 24, 1–9. [Google Scholar]
- Tanaka, H.; Ohshimo, S.; Takagi, N.; Ichimaru, T. Investigation of the geographical origin and migration of anchovy Engraulis japonicus in Tachibana Bay, Japan: A stable isotope approach. Fish. Res. 2010, 102, 217–220. [Google Scholar] [CrossRef]
- Gong, Y.; Li, Y.; Chen, X.J. Trophic niche and diversity of a pelagic squid (Dosidicus gigas): A comparative study using stable isotope, fatty acid, and feeding apparatuses morphology. Front. Mar. Sci. 2020, 7, 642. [Google Scholar] [CrossRef]
- Han, D.Y.; Xue, Y. Trophic and spatial ecological niche of five goby species in Jiaozhou Bay. Chin. Aquat. Sci. 2013, 20, 148–156. [Google Scholar]
- Zhao, Z.; Hu, G.; Fang, Z.; Li, J.; Liu, B.; Chen, X. Feeding Strategie and Trophic Niche Divergence of Three Groups of Dosidicus gigas off Peru: Based on Stable Carbon and Nitrogen Isotopes and Morphology of Feeding Apparatuses. Mar. Biotechnol. 2023, 25, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Katsumi, M.; Makoto, S.; Yukio, N. Food density and rate of feeding larvae of anchovy and sardine in patchy distribution. Jpn. J. Fish. Sci. 1988, 54, 401–411. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods as-sumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Yasue, N.; Doiuchi, R.; Yoshimoto, Y.; Takeuchi, T. Diet of late larval Japanese anchovy Engraulis japonicus in the Kii Channel, Japan. Fish. Sci. 2010, 76, 63–73. [Google Scholar] [CrossRef]
- Yasunaka, S.; Nojiri, Y.; Nakaoka, S.I.; Ono, T.; Mukai, H.; Usui, N. Monthly maps of sea surface dissolved inorganic carbon in the North Pacific: Basin-wide distribution and seasonal variation. J. Geophys. Res. Ocean. 2013, 118, 3843–3850. [Google Scholar] [CrossRef]
- Loseto, L.L.; Stern, G.A.; Deibel, D.; Connelly, T.L.; Prokopowicz, A.; Lean, D.R.S.; Ferguson, S.H. Linking mercury exposure to habitat and feeding behaviour in Beaufort Sea beluga whales. J. Mar. Syst. 2008, 74, 1012–1024. [Google Scholar] [CrossRef]
- Loseto, L.L.; Stern, G.A.; Ferguson, S.H. Size and biomagnification: How habitat selection explains beluga mercury levels. Environ. Sci. Technol. 2008, 42, 3982–3988. [Google Scholar] [CrossRef]
- Kim, D.I. Metabolic Rates of Japanese Anchovy (Engraulis japonicus) during Early Development Using a Novel Modified Respirometry Method. Animals 2023, 13, 1035. [Google Scholar] [CrossRef]
- Wan, R.; Wu, Y.; Huang, L.; Zhang, J.; Gao, L.; Wang, N. Fatty acids and stable isotopes of a marine ecosystem: Study on the Japanese anchovy (Engraulis japonicus) food web in the Yellow Sea. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 1047–1057. [Google Scholar] [CrossRef]
- Wang, R.; Wang, W.X. Diet-specific trophic transfer of mercury in tilapia (Oreochromis niloticus): Biodynamic perspective. Environ. Pollut. 2018, 234, 288–296. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locations | Date | Number | Min | Max | Mean (SD) |
|---|---|---|---|---|---|
| 39.23° N; 155.66° E | July 2021 | 8 | 121 | 110 | 115.23 ± 4.21 |
| 39.37° N; 154° E | June 2021 | 11 | 93 | 105 | 97.17 ± 2.32 |
| 40.35° N; 156° E | July 2021 | 17 | 96 | 125 | 112.55 ± 5.32 |
| 40.97° N; 155.5° E | July 2021 | 12 | 91 | 115 | 100.73 ± 4.43 |
| 41.00° N; 161.50° E | July 2021 | 22 | 117 | 140 | 132.02 ± 8.43 |
| 41.32° N; 160.03° E | July 2021 | 20 | 105 | 140 | 120.36 ± 8.39 |
| 41.68° N; 156.00° E | July 2021 | 8 | 105 | 131 | 117.22 ± 5.39 |
| 42.00° N; 160.57° E | July 2021 | 11 | 111 | 129 | 119.67 ± 4.93 |
| 42.00° N; 157.00° E | July 2021 | 13 | 98 | 131 | 112.41 ± 3.27 |
| 42.50° N; 155.58° E | July 2021 | 21 | 95 | 134 | 125.83 ± 8.12 |
| Body Length Group | Number | δ15N (‰) | δ13C (‰) | TL | ||||
|---|---|---|---|---|---|---|---|---|
| Max | Min | Mean ± SD | Max | Min | Mean ± SD | |||
| 91–100 mm | 27 | 9.97 | 6.69 | 8.17 ± 0.76 | −19.67 | −20.43 | −19.99 ± 0.21 | 2.85 |
| 101–110 mm | 30 | 10.06 | 6.20 | 8.06 ± 1.04 | −19.53 | −20.37 | −19.97 ± 0.21 | 2.82 |
| 111–120 mm | 30 | 10.67 | 6.60 | 8.44 ± 1.13 | −18.68 | −20.57 | −20.00 ± 0.33 | 2.93 |
| 121–130 mm | 27 | 10.11 | 6.46 | 8.77 ± 0.99 | −19.65 | −20.66 | −20.01 ± 0.21 | 3.03 |
| 131–140 mm | 28 | 10.53 | 6.65 | 9.03 ± 1.06 | −19.16 | −20.40 | −19.92 ± 0.22 | 3.11 |
| Group | 91–100 mm | 101–110 mm | 111–120 mm | 121–130 mm | 131–140 mm | SEAc |
|---|---|---|---|---|---|---|
| 91–100 mm | — | 83.01% | 73.80% | 45.91% | 31.11% | 0.52 |
| 101–110 mm | — | — | 78.81% | 45.14% | 35.94% | 0.68 |
| 111–120 mm | — | — | — | 70.62% | 46.79% | 0.80 |
| 121–130 mm | — | — | — | — | 52.77% | 0.59 |
| 131–140 mm | — | — | — | — | — | 0.69 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Hu, G.; Zhao, Z.; Liu, B.; Chen, X.; Chen, Y. Ontogenetic Variation in the Trophic and Mercury Levels of Japanese Anchovy in the High Seas of the Northwestern Pacific Ocean. J. Mar. Sci. Eng. 2024, 12, 1274. https://doi.org/10.3390/jmse12081274
Chen L, Hu G, Zhao Z, Liu B, Chen X, Chen Y. Ontogenetic Variation in the Trophic and Mercury Levels of Japanese Anchovy in the High Seas of the Northwestern Pacific Ocean. Journal of Marine Science and Engineering. 2024; 12(8):1274. https://doi.org/10.3390/jmse12081274
Chicago/Turabian StyleChen, Long, Guanyu Hu, Zhenfang Zhao, Bilin Liu, Xinjun Chen, and Yingcong Chen. 2024. "Ontogenetic Variation in the Trophic and Mercury Levels of Japanese Anchovy in the High Seas of the Northwestern Pacific Ocean" Journal of Marine Science and Engineering 12, no. 8: 1274. https://doi.org/10.3390/jmse12081274
APA StyleChen, L., Hu, G., Zhao, Z., Liu, B., Chen, X., & Chen, Y. (2024). Ontogenetic Variation in the Trophic and Mercury Levels of Japanese Anchovy in the High Seas of the Northwestern Pacific Ocean. Journal of Marine Science and Engineering, 12(8), 1274. https://doi.org/10.3390/jmse12081274