
Genotype by Environment Interaction for Growth in Atlantic Cod (Gadus morhua L.) in Four Farms of Norway

Abstract
:1. Introduction
2. Materials and Methods
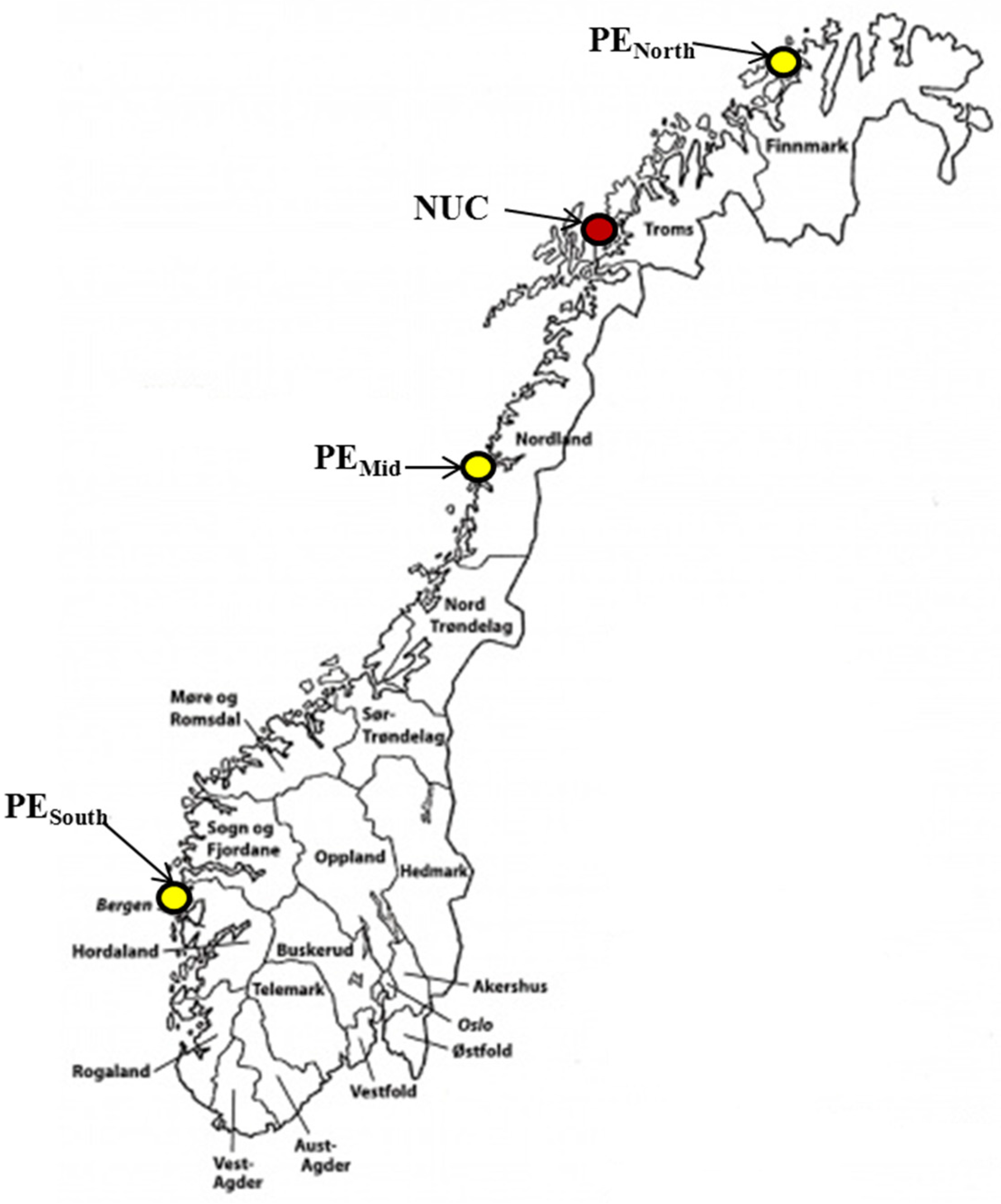
2.1. Fish Materials

{kind=link}
{kind=link}
| Year class | Farms | Fish, no | Family, no | Sires, no | Dams, no | Fish per Family | TWT, g | TAG, day | AG, day | BW, g | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Min | Max | Mean | s.d. | Mean | s.d. | Mean | s.d. | Mean | s.d. | ||||||
| 2007 | NUC | 10,685 | 185 | 140 | 185 | 57 | 4 | 112 | 27 | 9 | 205 | 3 | 977 | 9 | 2431 | 600 |
| PENorth | 924 | 155 | 125 | 155 | 6 | 1 | 15 | 27 | 9 | 205 | 2 | 959 | 6 | 2414 | 582 | |
| PESouth | 2202 | 141 | 116 | 141 | 15 | 5 | 45 | 28 | 9 | 205 | 2 | 966 | 7 | 1880 | 442 | |
| 2010 | NUC | 6977 | 193 | 132 | 180 | 36 | 12 | 79 | 26 | 8 | 202 | 2 | 945 | 10 | 2715 | 569 |
| PEMid | 2172 | 178 | 127 | 167 | 12 | 1 | 21 | 26 | 8 | 202 | 2 | 998 | 9 | 3365 | 673 | |
| All farms | 22,960 | 379 | 273 | 367 | 28 | 9 | 204 | 3 | 966 | 19 | 2552 | 680 | ||||
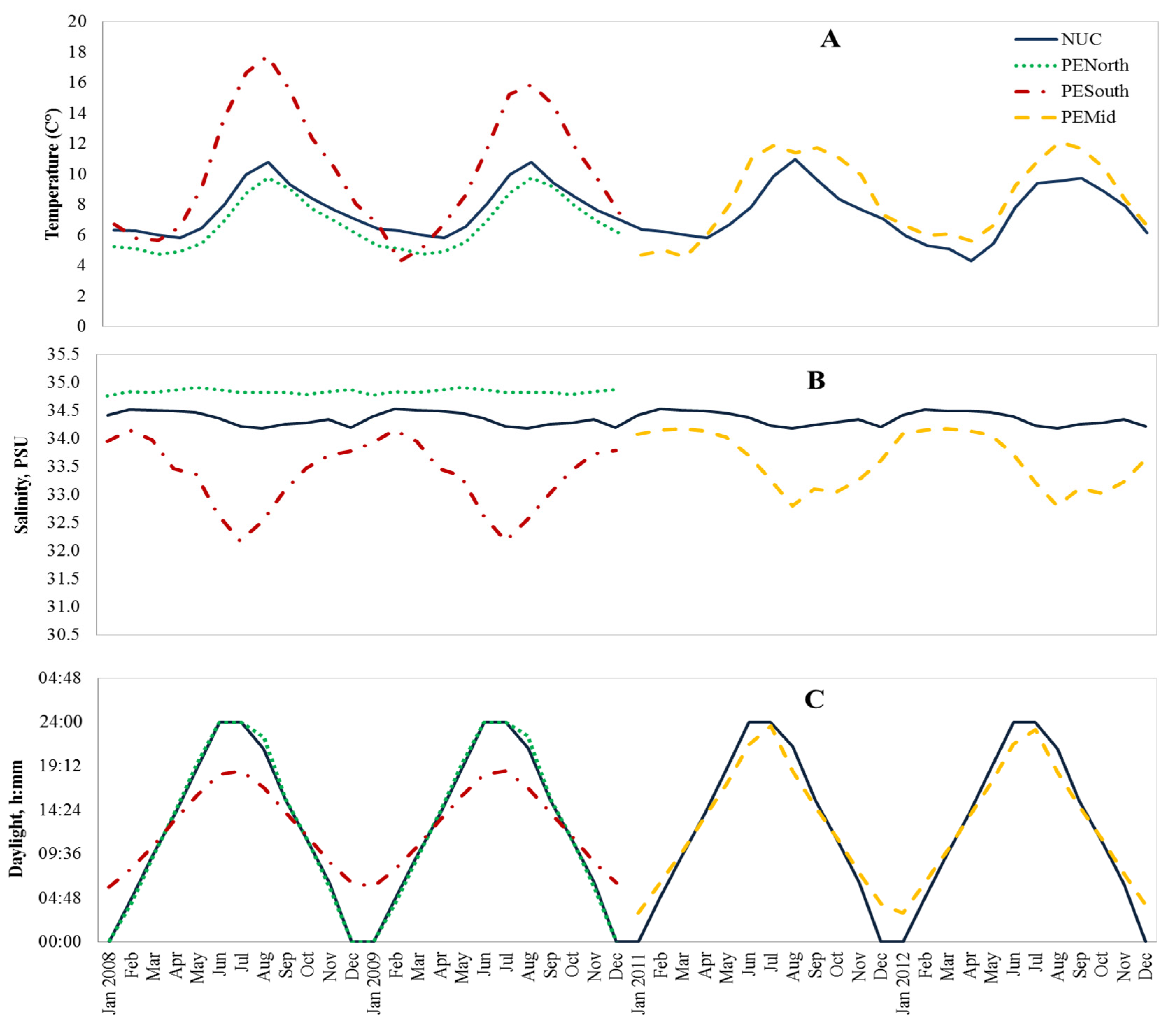
2.2. Test Environments


2.3. Data Collection
2.4. Statistical Analyses
2.4.1. Univariate Analysis
2.4.2. Multivariate Analysis
2.4.3. Calculation of Genetic Parameters
3. Results
3.1. Descriptive Statistics
3.2. Heterogeneity of Genetic Variation
| Year-class | Farm | ± s.e. | ± s.e.* | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2007 | NUC | 318,420 | 50,498 | 247,060 | 23.21 | 9.24 | 20.45 | 0.16 ± 0.06 | 0.07 ± 0.02 |
| PENorth | 311,780 | 31,634 | 249,250 | 23.13 | 7.37 | 20.68 | 0.10 ± 0.10 | 0.10 ± 0.05 | |
| PESouth | 175,840 | 21,675 | 134,460 | 22.30 | 7.83 | 19.50 | 0.12 ± 0.09 | 0.11 ± 0.04 | |
| 2010 | NUC | 305,970 | 78,629 | 205,300 | 20.37 | 10.33 | 16.69 | 0.26 ± 0.07 | 0.07 ± 0.03 |
| PEMid | 444,830 | 34,401 | 375,410 | 19.82 | 5.51 | 18.21 | 0.08 ± 0.06 | 0.08 ± 0.03 |
3.3. Genetic Correlations
| Full-model a | Reduced-model a | ||||||
|---|---|---|---|---|---|---|---|
| Farms | NUC | PENorth | PESouth | PEMid | PENorth | PESouth | PEMid |
| NUC | 0.92 ± 0.38 | 0.96 ± 0.17 | 0.81 ± 0.19 | 0.81 ± 0.08 | 0.86 ± 0.04 | 0.84 ± 0.05 | |
| PENorth | 0.68 ± 0.18 | 0.07 ± 0.65 | - | 0.63 ± 0.11 | - | ||
| PESouth | 0.77 ± 0.12 | 0.84 ± 0.15 | - | - | |||
| PEMid | 0.69 ± 0.23 | - | - | ||||
4. Discussion
4.1. Genotype by Environment Interaction
4.2. Implications for Breeding
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sae-Lim, P.; Komen, H.; Kause, A.; Mulder, H.A. Identifying environmental variables explaining genotype-by-environment interaction for body weight of rainbow trout (Onchorynchus mykiss): Reaction norm and factor analytic models. Genet. Sel. Evol. 2014, 46, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, M.; Walsh, B. Genetics and Analysis of Quantitative Traits; Sinauer Associates: Sunderland, MA, USA, 1998. [Google Scholar]
- Falconer, D.S.; Mackay, T.F. Introduction to Quantitative Genetics; Longman: New York, NY, USA, 1996. [Google Scholar]
- Mulder, H.; Bijma, P. Effects of genotype × environment interaction on genetic gain in breeding programs. J. Anim. Sci. 2005, 83, 49–61. [Google Scholar] [PubMed]
- Przystalski, M.; Osman, A.; Thiemt, E.; Rolland, B.; Ericson, L.; Østergård, H.; Levy, L.; Wolfe, M.; Büchse, A.; Piepho, H.-P. Comparing the performance of cereal varieties in organic and non-organic cropping systems in different european countries. Euphytica 2008, 163, 417–433. [Google Scholar] [CrossRef]
- Charmantier, A.; Garant, D. Environmental quality and evolutionary potential: Lessons from wild populations. Proc. R. Soc. Lond. B Biol. Sci. 2005, 272, 1415–1425. [Google Scholar] [CrossRef] [PubMed]
- Kolstad, K.; Thorland, I.; Refstie, T.; Gjerde, B. Genetic variation and genotype by location interaction in body weight, spinal deformity and sexual maturity in atlantic cod (Gadus morhua) reared at different locations off norway. Aquaculture 2006, 259, 66–73. [Google Scholar] [CrossRef]
- Bangera, R.; Ødegård, J.; Præbel, A.K.; Mortensen, A.; Nielsen, H.M. Genetic correlations between growth rate and resistance to vibriosis and viral nervous necrosis in atlantic cod (Gadus morhua L.). Aquaculture 2011, 317, 67–73. [Google Scholar] [CrossRef]
- Karlsen, Ø.; Holm, J.C.; Kjesbu, O.S. Effects of periodic starvation on reproductive investment in first-time spawning atlantic cod (Gadus morhua L.). Aquaculture 1995, 133, 159–170. [Google Scholar]
- Iwamoto, R.; Myers, J.; Hershberger, W. Genotype-environment interactions for growth of rainbow trout, Salmo gairdneri. Aquaculture 1986, 57, 153–161. [Google Scholar] [CrossRef]
- Dupont-Nivet, M.; Karahan-Nomm, B.; Vergnet, A.; Merdy, O.; Haffray, P.; Chavanne, H.; Chatain, B.; Vandeputte, M. Genotype by environment interactions for growth in european seabass (Dicentrarchus labrax L.) are large when growth rate rather than weight is considered. Aquaculture 2010, 306, 365–368. [Google Scholar]
- Fevolden, S.; Pogson, G. Genetic divergence at the synaptophysin (Syp I) locus among norwegian coastal and north-east arctic populations of atlantic cod. J. Fish. Biol. 1997, 51, 895–908. [Google Scholar]
- Pogson, G.H.; Mesa, K.A.; Boutilier, R.G. Genetic population structure and gene flow in the atlantic cod Gadus morhua: A comparison of allozyme and nuclear rflp loci. Genetics 1995, 139, 375–385. [Google Scholar] [PubMed]
- Ødegård, J.; Sommer, A.I.; Præbel, A.K. Heritability of resistance to viral nervous necrosis in atlantic cod (Gadus morhua L.). Aquaculture. 2010, 300, 59–64. [Google Scholar] [CrossRef]
- United states naval observatory (USNO). Available online: http://aa.Usno.Navy.Mil (accessed on 12 December 2014).
- Holm, J. Ultrasonography, a non–invasive method for sex determination in cod (Gadus morhua). J. Fish. Biol. 1994, 44, 965–971. [Google Scholar]
- SAS Institute Inc. SAS 9.3 Output Delivery System: User’s Guide.; SAS Institute: Cary, NC, USA, 2011. [Google Scholar]
- Gilmour, A.R.; Gogel, B.; Cullis, B.; Thompson, R. Asreml User Guide Release 3.0; VSN International Ltd.: Hemel Hempstead, UK, 2009. [Google Scholar]
- Stram, D.O.; Lee, J.W. Variance components testing in the longitudinal mixed effects model. Biometrics 1994, 50, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Sae-Lim, P.; Kause, A.; Mulder, H.; Martin, K.; Barfoot, A.; Parsons, J.; Davidson, J.; Rexroad, C.; van Arendonk, J.; Komen, H. Genotype-by-environment interaction of growth traits in rainbow trout (Oncorhynchus mykiss): A continental scale study. J. Anim. Sci. 2013, 91, 5572–5581. [Google Scholar] [CrossRef] [PubMed]
- Gjedrem, T.; Baranski, M. Selective Breeding in Aquaculture: An. Introduction; Springer Verlag: Berlin, Germany, 2009; Volume 10. [Google Scholar]
- Gjerde, B.; Simianer, H.; Refstie, T. Estimates of genetic and phenotypic parameters for body weight, growth rate and sexual maturity in atlantic salmon. Livest. Prod. Sci. 1994, 38, 133–143. [Google Scholar] [CrossRef]
- Sylvén, S.; Rye, M.; Simianer, H. Interaction of genotype with production system for slaughter weight in rainbow trout (Oncorhynchus mykiss). Livest. Prod. Sci. 1991, 28, 253–263. [Google Scholar] [CrossRef]
- Khaw, H.L.; Ponzoni, R.W.; Hamzah, A.; Abu-Bakar, K.R.; Bijma, P. Genotype by production environment interaction in the gift strain of nile tilapia (Oreochromis niloticus). Aquaculture 2012, 326, 53–60. [Google Scholar] [CrossRef]
- Maluwa, A.O.; Gjerde, B.; Ponzoni, R.W. Genetic parameters and genotype by environment interaction for body weight of Oreochromis shiranus. Aquaculture 2006, 259, 47–55. [Google Scholar] [CrossRef]
- Gjedrem, T. Selection and Breeding Programs in Aquaculture; Springer Verlag: Berlin, Germany, 2005. [Google Scholar]
- Dupont-Nivet, M.; Vandeputte, M.; Vergnet, A.; Merdy, O.; Haffray, P.; Chavanne, H.; Chatain, B. Heritabilities and gxe interactions for growth in the european sea bass (Dicentrarchus labrax L.) using a marker-based pedigree. Aquaculture 2008, 275, 81–87. [Google Scholar] [CrossRef]
- Imsland, A.K.; Foss, A.; Folkvord, A.; Stefansson, S.O.; Jonassen, T.M. Genotypic response to photoperiod treatment in atlantic cod (Gadus morhua). Aquaculture 2005, 250, 525–532. [Google Scholar] [CrossRef]
- Otterlei, E.; Nyhammer, G.; Folkvord, A.; Stefansson, S.O. Temperature-and size-dependent growth of larval and early juvenile atlantic cod (Gadus morhua): A comparative study of norwegian coastal cod and northeast arctic cod. Can. J. Fish. Aquat. Sci. 1999, 56, 2099–2111. [Google Scholar] [CrossRef]
- Svåsand, T.; Jørstad, K.; Otterå, H.; Kjesbu, O. Differences in growth performance between arcto-norwegian and norwegian coastal cod reared under identical conditions. J. Fish. Biol. 1996, 49, 108–119. [Google Scholar] [CrossRef]
- Kause, A.; Ritola, O.; Paananen, T.; Wahlroos, H.; Mäntysaari, E.A. Genetic trends in growth, sexual maturity and skeletal deformations, and rate of inbreeding in a breeding programme for rainbow trout (Oncorhynchus mykiss). Aquaculture 2005, 247, 177–187. [Google Scholar] [CrossRef]
- Mulder, H.A.; Bijma, P.; Hill, W.G. Prediction of breeding values and selection responses with genetic heterogeneity of environmental variance. Genetics 2007, 175, 1895–1910. [Google Scholar] [CrossRef] [PubMed]
- Sonesson, A.K.; Ødegård, J.; Rönnegård, L. Genetic heterogeneity of within-family variance of body weight in atlantic salmon (Salmo salar). Genet. Sel. Evol. GSE 2013, 45, 41. [Google Scholar] [CrossRef] [PubMed]
- Janhunen, M.; Kause, A.; Vehviläinen, H.; Järvisalo, O. Genetics of microenvironmental sensitivity of body weight in rainbow trout (Oncorhynchus mykiss) selected for improved growth. PLoS ONE 2012, 7, e38766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visscher, P.M.; Hill, W.G.; Wray, N.R. Heritability in the genomics era—Concepts and misconceptions. Nat. Rev. Genet. 2008, 9, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Kettunen, A.; Serenius, T.; Fjalestad, K. Three statistical approaches for genetic analysis of disease resistance to vibriosis in atlantic cod (Gadus morhua L.). J. Anim. Sci. 2007, 85, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Cameron, N. Methodologies for estimation of genotype with environment interaction. Livest. Prod. Sci. 1993, 35, 237–249. [Google Scholar] [CrossRef]
- Tosh, J.J.; Garber, A.F.; Trippel, E.A.; Robinson, J.A.B. Genetic, maternal, and environmental variance components for body weight and length of atlantic cod at 2 points in life. J. Anim. Sci. 2010, 88, 3513–3521. [Google Scholar] [CrossRef] [PubMed]
- Sae-Lim, P.; Komen, H.; Kause, A. Bias and precision of estimates of genotype-by-environment interaction: A simulation study. Aquaculture 2010, 310, 66–73. [Google Scholar] [CrossRef]
- Arnold, S.J. Multivariate Inheritance and Evolution: A Review of Concepts. In Quantitative Genetic Studies of Behavioral Evolution; University of Chicago Press: Chicago, IL, USA, 1994; pp. 17–48. [Google Scholar]
- Gjerde, B. Design of Breeding Programs. In Selection and Breeding Programs in Aquaculture; Springer: Berlin, Germany, 2005; pp. 173–195. [Google Scholar]
- Martinez, V.; Kause, A.; Mäntysaari, E.; Mäki-Tanila, A. The use of alternative breeding schemes to enhance genetic improvement in rainbow trout (Oncorhynchus mykiss): I. One-stage selection. Aquaculture 2006, 254, 182–194. [Google Scholar] [CrossRef]
- Bourdon, R.M. Understanding Animal Breeding; Prentice Hall: Upper Saddle River, NJ, USA, 2000; Volume 2. [Google Scholar]
- Meuwissen, T.; De Jong, G.; Engel, B. Joint estimation of breeding values and heterogeneous variances of large data files. J. Dairy Sci. 1996, 79, 310–316. [Google Scholar] [CrossRef]
- Namkoong, G. The influence of composite traits on genotype by environment relations. Theor. Appl. Genet. 1985, 70, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Rutten, M.; Bijma, P.; Woolliams, J.; Van Arendonk, J. Selaction: Software to predict selection response and rate of inbreeding in livestock breeding programs. J. Hered. 2002, 93, 456–458. [Google Scholar] [CrossRef] [PubMed]
- Mulder, H.; Veerkamp, R.; Ducro, B.; van Arendonk, J.; Bijma, P. Optimization of dairy cattle breeding programs for different environments with genotype by environment interaction. J. Dairy Sci. 2006, 89, 1740–1752. [Google Scholar] [CrossRef]
- James, J.W. Selection in two environments. Heredity 1961, 16, 145–152. [Google Scholar] [CrossRef]
- Sae-Lim, P. One size fits all?: Optimization of Rainbow Trout Breeding Program under Diverse Preferences and Genotype-By-Environment Interaction. Doctoral Thesis, The Wageningen University, Wageningen, The Nethelands, 2013. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bangera, R.; Drangsholt, T.M.K.; Nielsen, H.M.; Sae-Lim, P.; Ødegård, J.; Puvanendran, V.; Hansen, Ø.J.; Mortensen, A. Genotype by Environment Interaction for Growth in Atlantic Cod (Gadus morhua L.) in Four Farms of Norway. J. Mar. Sci. Eng. 2015, 3, 412-427. https://doi.org/10.3390/jmse3020412
Bangera R, Drangsholt TMK, Nielsen HM, Sae-Lim P, Ødegård J, Puvanendran V, Hansen ØJ, Mortensen A. Genotype by Environment Interaction for Growth in Atlantic Cod (Gadus morhua L.) in Four Farms of Norway. Journal of Marine Science and Engineering. 2015; 3(2):412-427. https://doi.org/10.3390/jmse3020412
Chicago/Turabian StyleBangera, Rama, Tale M. K. Drangsholt, Hanne Marie Nielsen, Panya Sae-Lim, Jørgen Ødegård, Velmurugu Puvanendran, Øyvind J. Hansen, and Atle Mortensen. 2015. "Genotype by Environment Interaction for Growth in Atlantic Cod (Gadus morhua L.) in Four Farms of Norway" Journal of Marine Science and Engineering 3, no. 2: 412-427. https://doi.org/10.3390/jmse3020412