1. Introduction
Water cooling using natural waters is typically utilized for cooling large industrial facilities such as power plants. Once-through systems are a common choice, when abundant water resources such as lakes, rivers, or seas are accessible. Industrial coolers are systems in which the excess heat is transferred through metal surfaces to colder, circulating water [
1]. Cooling systems utilizing natural water are susceptible to biofouling, which refers to the undesirable accumulation of biotic deposits on a surface [
2]. There is a known sequence of biofouling formation: first the surfaces are colonized by microorganisms that prepare the surface conditions suitable for settlement of other biofouling species [
2,
3]. Microfouling, i.e., microbial biofilm formation, on the surfaces of materials in the water cycles can enhance corrosion of these materials. In the industrial systems, the biofilm formation together with inorganic fouling and scaling can cause problems such as an increase in the flow resistance of pipelines or a decrease in the thermal transfer capacity of the heat exchangers [
4]. It has been estimated that 20% of all corrosion damage of the heat exchangers is caused or influenced by microorganisms [
4]. The efficiency and availability of a power plant depends to a great extent on the performance and cleanness of materials used. More than 5% of the efficiency of a power plant can be lost due to biofouling on surfaces [
5].
Chemical cleaning of the affected area is the main method used for removing unwanted biofilms in the cooling water systems. Currently the most common biocide to prevent biofouling in once-through cooling systems is hypochlorite [
5,
6]. The biocidal effect of hypochlorite is due to its oxidative effect that causes it to react with bromide, which is naturally present in seawater and also possibly with other compounds, both organic and inorganic, leading to the formation of hypobromite and halogenated organic compounds [
5,
7]. The high oxidative power of the hypobromite, formed through the above-mentioned reaction, cause fast inactivation of enzymes and other cellular components of microorganisms. The usage of hypochlorite, however, raises environmental concerns due to production of by-products that are toxic to aquatic organisms [
7,
8].
In addition to biocides, biofouling may be prevented by application of coatings developed to prevent the settlement of fouling organisms. The functionality of coatings is based on either biocide release, which inhibits fouling settlement by releasing biocidal products into the marine environment, or nontoxic fouling release coatings, whose antifouling efficiency relies on low adhesion strength and easy release of fouling organisms from such a coating [
9]. Coatings also allow the use of less corrosion resistant metals such as carbon steel, since the base material is not in direct contact with seawater. However, biofilm forming microorganisms are known to deteriorate various polymeric materials, including some coating materials [
10,
11]. Even though the microfouling species would not directly cause deterioration of the coatings, the original antifouling properties such as hydrophobicity of surface can be masked [
10].
For effective antifouling treatment, all factors including the environmental impact of biocides and the cost of biocide treatments and antifouling coatings have to be considered [
7]. Given the costs of biofouling treatment, power plants require biofouling and biocorrosion control solutions that are economically efficient, easy to apply to the system, as well as environmentally sustainable. This can be achieved by adjusting the timing and magnitude of chlorination in combination with special antifouling surfaces [
6,
12,
13,
14]. There is a need, however, for deeper understanding of the structure of the microbial community responsible for biofouling of the surfaces under real industrial conditions.
The aim of this research was: (1) to compare the suitability of two commercially available coatings to be used in cooling water systems using hypochlorite as a biocide; (2) to characterize the primary biofilm forming a microbial community in system with or without a hypochlorite treatment and on different coatings. Here we focused on microbial fouling community (bacteria, archaea, and fungi) that attached to the surface of coated (fouling release) carbon steel in a power plant utilizing brackish seawater from the Baltic Sea. Hypochlorite treatment was used to prevent biological fouling in the cooling water system during summer months, when the seawater temperature rises and biofouling is more intensive. The results of a three-month survey of the effect of chlorination on coating performance and species composition of microfouling on coated surfaces are presented and discussed.
2. Materials and Methods
2.1. Materials
The test material was coated with low alloy carbon steel (corresponding with the composition of carbon steel AISI 1005) from cold-rolled thin sheet (1 mm). The surface of the carbon steel was as received and the coupon size was 25 mm × 75 mm. Two different coatings, a biocide free silicon coating, BioCleanEco (CMP, Tokyo, Japan) and a synthetic purified epoxy tar coating TeknoTar100 (Teknos, Helsinki, Finland), were applied to the carbon steel specimens as instructed by the manufacturers. The coupons were cleaned with the FreeBact-20 (AquaFix, Satsjöbaden, Sweden) and sterile ultra pure water and air-dried before exposure. Each biofouling cell contained eight coupons of each coating. Three parallel coupons were used for material analyses, three coupons for molecular biological analyses, and two for microscopy subsequently. The coupons were photographed prior to the experiment.
2.2. Experimental Procedure
To study the fouling in the cooling water cycle, a specially developed biofouling cell was installed into the cooling water cycle (
Figure 1) at a power plant where the hypochlorite treatment is routinely used to control biofouling. Two units were installed in the cycle before the chlorination point (“controls”) and two units after the hypochlorite dosage point (“hypochlorite”). The water flow through the biofouling cell was 0.1 m·s
−1.
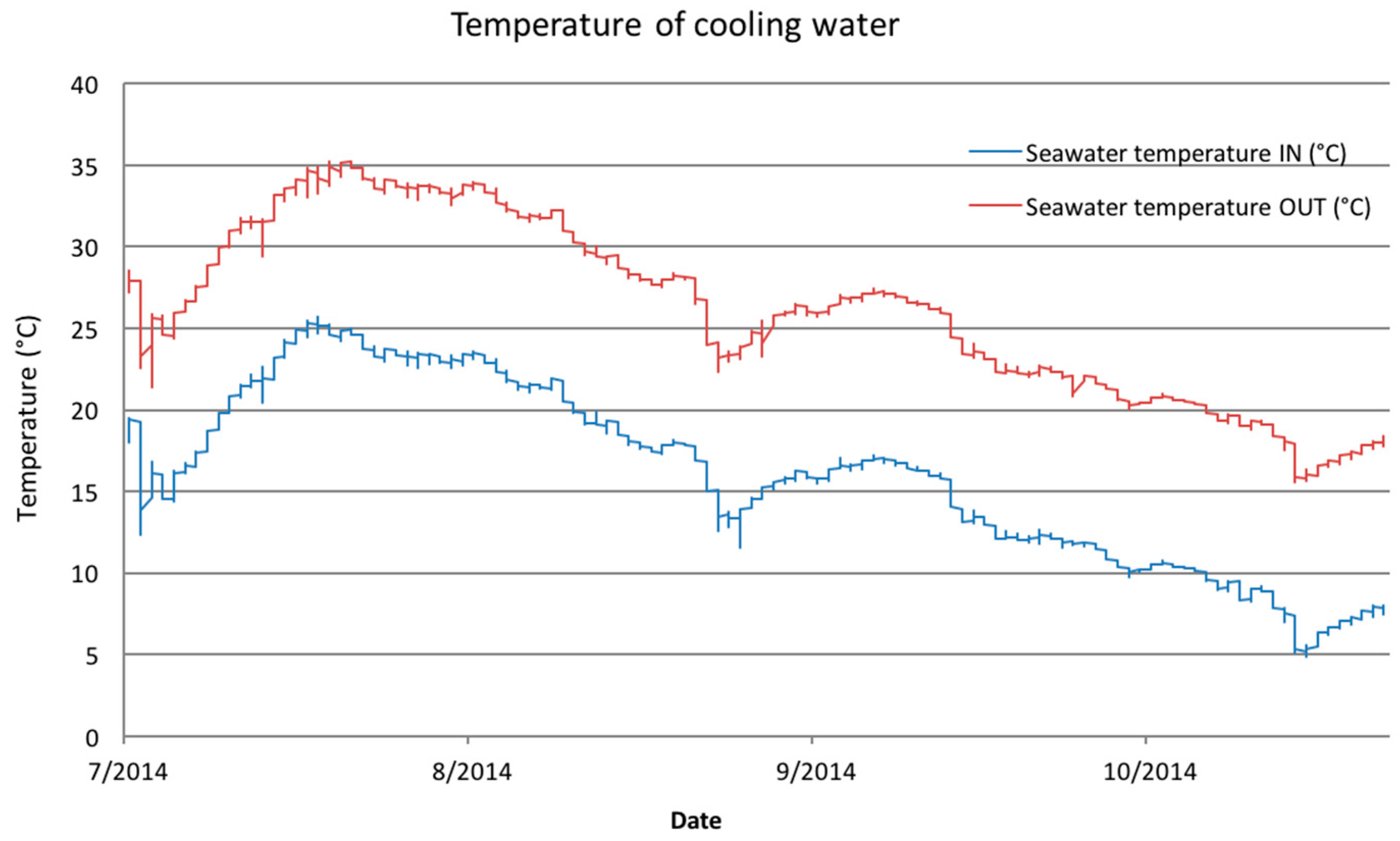
Na-hypochlorite (15%) was dosed in the cooling system to obtain a final target concentration of 3 mg·L−1. During the first 30 days chlorination was applied for two hours with 10 h breaks between the treatments, and after that for one hour with 11 h breaks in between. The experiment period started at July 2014, a week after chlorination had already started. The first set of biofouling cells, one control cell and one cell after the hypochlorite dosage point were removed from the cooling water cycle after 32 days of exposure (one month). The second sampling was performed after 90 days of exposure (3 months). The biofouling cells were removed from the water cycle and sealed so that water remained inside the cells. The cells were dismantled aseptically in the laboratory.
The temperature of seawater used for cooling (seawater IN) and the temperature of cooling water after the cycle (seawater OUT) were monitored continuously during the experiment period. The chemical parameters of seawater used for cooling were measured once a month during the experiment period.
2.3. Surface Characterization
The coupons for the material studies were immersed in 96% ethanol, air-dried, and stored in a glass desiccator until studied. All coupons for material analyses were photographed and examined with a stereomicroscope.
The condition of the exposed coated surfaces was examined using a Zeiss ULTRAplus field emission scanning electron microscope (Carl Zeiss Microscopy GmbH, Jena, Germany).
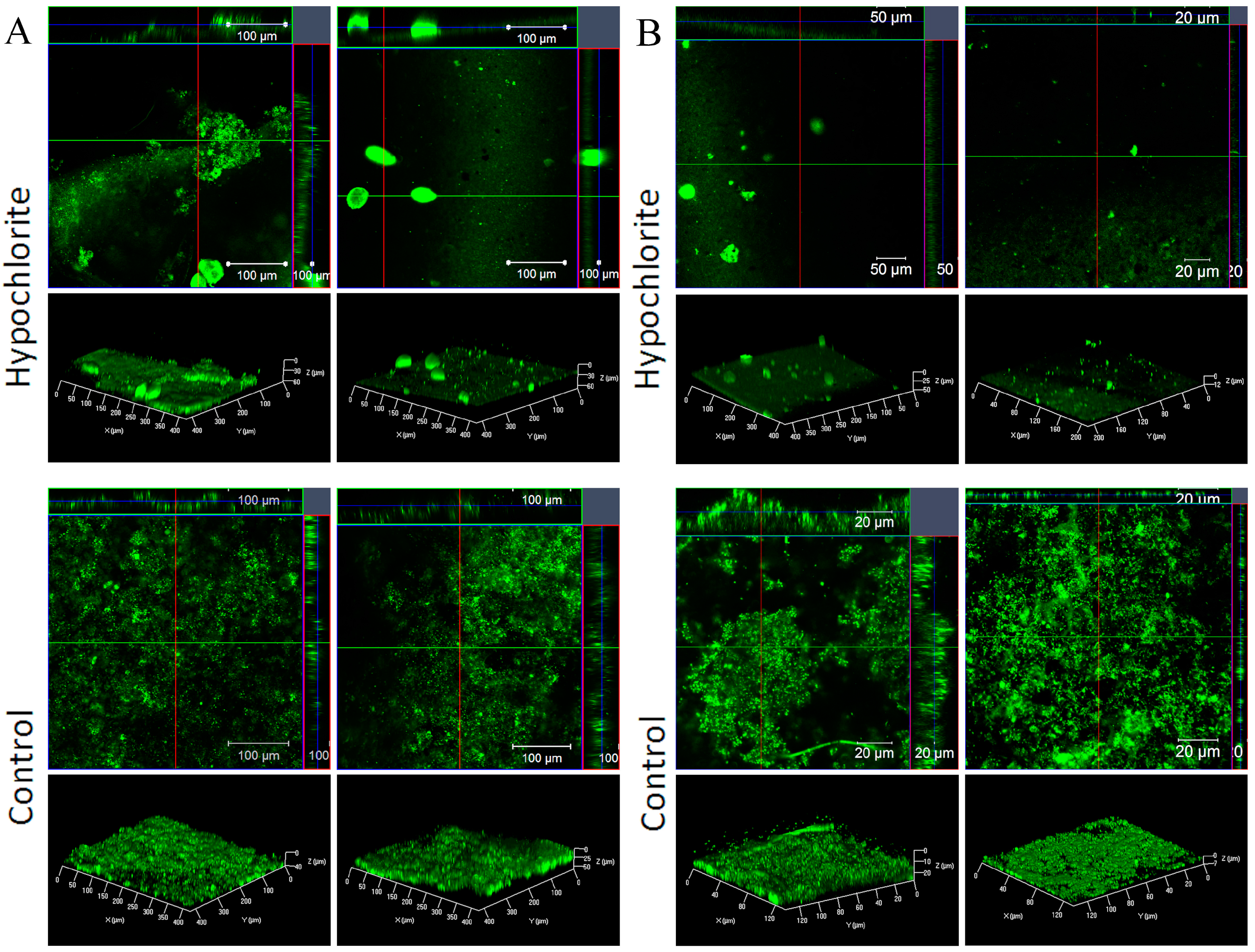
For the confocal microscopy, the samples were gently rinsed in water and stained with the DNA specific fluorescence dye SYTO9 (Thermo Fisher Scientific, Eugene, OR, USA) for 25 min in dark at room temperature. Visualization of the biofilms was done by confocal laser scanning microscopy (CLSM) using a Zeiss LSM 710 microscope (Carl Zeiss Microscopy GmbH) using the EC Epiplan-Neofluar 20 × 0.5 NA or the 63 × 0.9 NA (oil) objectives. The 488 nm Ar laser was used and a green emission was collected at 498–566 nm. The images were acquired and analyzed using the ZenPro (black edition) software (Carl Zeiss Microscopy GmbH). Z-stacks (vertical sections) were recorded with the optimal parameters suggested by the software. The number of the Z-stacks depended on the biofilm thickness. Auto-fluorescence of the coatings was checked using unused samples. The TerknoTar100 coating showed auto-fluorescence in the green area of the spectrum.
2.4. DNA Extraction
The coupons for microbiological analyses were subjected to the biofilm extraction immediately after they were removed from the biofouling cell. The biomass from the coupon surfaces was removed by brushing for 60 s using Omron Sonic Style B458 brush (Omron, Wegalaan, The Netherlands). The brushes were placed aseptically into tubes containing 20 mL sterile Ringer-solution (Merck, Darmstadt, Germany). The tubes were subsequently vortexed for 60 s. The suspension containing the biomass was filtered on the Sterivex filter unit (Sterivex-GP 0.22 µm; Millipore, Billerica, MA, USA). From both sampling times (32 days and 90 days) 500 mL of chlorinated and non-chlorinated seawater was filtered onto Sterivex or Corning (Corning 0.22 µm CA 430626/PES 431161; Corning, NY, USA) filter units as two replicates. Sterivex filtering units and Corning filters were cut with a sterile scalpel and stored frozen (−80 °C) until DNA extraction. For DNA extraction, Sterivex filter units were aseptically broken with a hammer, and both Sterivex and Corning filters were cut into pieces and placed into the lysing tube of the DNA extraction kit. The DNA extraction was performed using a Fast DNA Spin kit for soil (MP Biomedicals, Santa Ana, CA, USA) according to manufacturer’s protocol with the modification that the cells were homogenised in a FastPrep-24 instrument (MP Biomedicals, Santa Ana, CA, USA) at 6 m·s−1 for 3 min.
2.5. Quantitative PCR
The number of bacteria forming biofilm on the surfaces was estimated by quantitative polymerase chain reaction (qPCR) of bacterial 16S ribosomal RNA (rRNA). An approximately 200 bp fragment of the 16S rRNA gene was amplified with primers 358F and 534R [
15] and detected with SYBR green based detection of double-stranded DNA, using LightCycler
® 480 qPCR machine (Roche Diagnostics, Basel, Switzerland). The DNA concentration of all samples was measured with Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific) and adjusted to ≤10 ng·µL
−1 with molecular-biology-grade water (Sigma, St. Louis, MO, USA) prior to qPCR in order to avoid PCR inhibition. The 10-µL reaction mixture contained 1 µL of DNA template, 1 × KAPA SYBR
® FAST Universal qPCR Master Mix (KAPA Biosystems, MA, USA), and 150 nM of both primers. The PCR program consisted of initial 15 min incubation at 95 °C, followed by 45 cycles of denaturation at 95 °C for 10 s, annealing at 57 °C for 35 s and extension at 72 °C for 30 s, followed by a final extension at 72 °C for 3 min and a melting curve analysis. As an external standard, a dilution series (100–10
8 colony forming units (cfu)·µL
−1) of
Escherichia coli VTT E-90418 genomic DNA was used. The results were calculated using Absolute quantification/second derivative maximum in LightCycler
® 480 Software 1.5.0 (Roche Diagnostics). The statistical significance of differences of the means was calculated applying two-way ANOVA (IBM SPSS Statistics version 21, Armonk, NY, USA).
2.6. Amplification Library Preparation
The amplification libraries for high-throughput sequencing with Ion Torrent PGM were prepared by PCR from the DNA samples. Bacterial 16S rRNA genes were amplified with the primers [
16], targeting the variable region V3-V4 of the 16S rDNA gene, archaeal 16S rRNA genes with the primers [
17], targeting the V4 region of the gene and fungal internal transcribed spacer (ITS) gene markers with the primer pair ITS1 and ITS2 targeting the fungal ITS1 region [
18]. PCR amplification was performed in parallel 25 μL reactions for every sample containing 1 × MyTaq™ Red Mix (Bioline, London, UK), 20 pmol of each primer, up to 25 μL molecular-biology-grade water (Sigma) and 2 μL of template. The PCR program consisted of an initial denaturation step at 95 °C for 3 min, 35 cycles for bacteria and fungi, and 40 cycles for archaea of 15 s at 95 °C, 15 s at 50 °C, and 15 s at 72 °C. A final elongation step of 30 s was performed at 72 °C. The PCR products were verified with agarose gel electrophoresis. Amplicons were sequenced using Ion Torrent PGM sequencing platform (Thermo Fisher Scinetific) at Bioser (University of Oulu, Finland) and amplicons were purified before sequencing.
2.7. Sequence Processing and Analysis
The sequence reads obtained from Ion Torrent sequencing were first subjected to quality control using the QIIME software [
19]. During this step, adapters, barcodes, and primers were removed from the sequence reads, and the quality of base-calls was assessed in order to remove erroneous reads from the data set. Subsequently, chimeric sequence reads, which are a type of sequencing artefact arising from sequences from separate sources fusing into one, were removed from the data set with the USEARCH algorithm version 5.2.236 [
20] by de novo detection and through similarity searches against the Greengenes reference database [
21] with bacterial and archaeal sequences and UNITE database [
22] with fungal sequences.
Groups of similar sequences—i.e., Operational Taxonomic Units (OTUs)—were selected from the chimera-filtered sequence data set following open-reference OTU-picking protocol of QIIME version 1.8.0 against the 97% identity Greengenes or UNITE reference database sets. OTU clustering was performed with UCLUST version 1.2.22q [
20] and the seed sequences were selected as the representative OTU sequences. All reads that failed to hit the Greengenes or UNITE reference database with a minimum of 60% identity threshold were discarded as sequencing error. Next, singleton OTUs—i.e., OTUs that were represented by a single sequence—were filtered from the data set. Finally, taxonomy from the domain-level up to species-level was assigned to OTUs via representative OTU sequences with the Ribosomal Database Project classifier algorithm at minimum of 80% confidence [
23] with bacterial and archaeal sequences. With fungal ITS sequences, taxonomic assignments were made by the BLASTN algorithm with a maximum
E-value of 0.001 [
20].
4. Discussion
Microfouling on the surfaces of studied anti-fouling coatings was intensive in the cooling water cycle during the warm summer months. As in most cases in natural water environments the fouling was formed by both inorganic fouling and biofouling. The formed biofilm was thick on the surfaces of untreated control samples. Our results indicated that the amount of bacteria on the surfaces from the control system was 10
8 cells·cm
−2. Similar numbers of bacterial microfouling have been reported on anti-fouling surfaces in oceanic waters, also in much warmer sites [
24,
25,
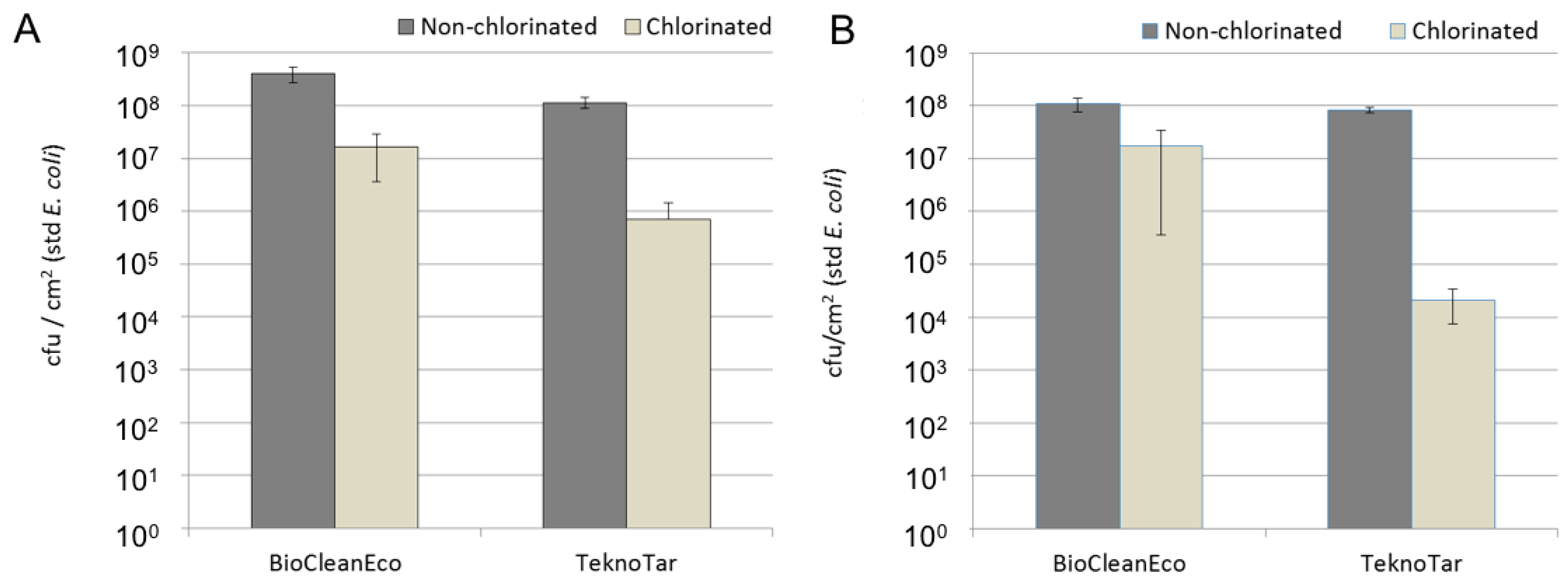
26]. Hypochlorite treatment reduced the bacterial numbers by 10–1000-fold, but the efficiency depended on the coating. The combination of TeknoTar coating and hypochlorite treatment was most efficient in reducing the attachment of bacteria, causing a 100–1000-fold decrease in number of attached bacteria compared to the untreated control, depending on the exposure time. With BioCleanEco the difference between the untreated control and hypochlorite-treated samples was 10-fold at both exposure times. The confocal results suggest that the chlorination of the cooling water probably did not prevent the bacterial adherence to the surface, but rather killed cells later, resulting in accumulation of the cell debris on the surface.
Biocidal coatings have been demonstrated to decrease the bacterial diversity of a microfouling community when compared to biocide-free coating [
27]. The antifouling coating used here did not produce as straightforward results combined with the hypochlorite treatment. The highest bacterial diversity was detected on TeknoTar coatings after a one-month exposure irrespective to whether they had received a hypochlorite treatment or not, even though the number of bacteria was significantly decreased due to the hypochlorite. On the other hand, the lowest bacterial diversity was detected on the three-month hypochlorite-treated BioCleanEco specimens.
In addition to decreasing the bacterial number, the hypochlorite treatment also changed the species composition forming the biofilm on the surfaces of two antifouling coatings. Whereas mainly Alphaproteobacteria formed the bacterial biofilm in the non-chlorinated system, in the hypochlorite-treated system the relative abundance of Beta- and Gammaproteobacteria increased. Cassé et al. [
27] compared the number of microfouling bacteria and the species composition on different coatings in a marine environment. Similar to our experiment, these researchers demonstrated that there was none or only little difference in the number of microfouling bacteria, but the species composition differed between the coatings [
27].
The dominant alphaproteobacterial family was
Rhodobacteraceae, which has previously been linked especially to early development of biofilm in marine environments, but also detected from this same environment earlier [
28,
29,
30,
31]. Many bacteria belonging to
Rhodobacteraceae are known to perform phototrophic iron oxidation [
32]. In addition, it has been speculated that the members of
Rhodobacteraceae form the initial biofilm since they are able to survive on nutrient poor conditions of early stages of biofilm formation. Moreover, these microorganisms are capable of forming biofilm on copper-based biocidal antifouling coating [
30,
33].
Archaeal fouling has not been studied as extensively as bacterial fouling and only few reports describe the archaeal community in marine fouling [
2,
31]. The family
Nitrosopumilus was the most commonly detected archaea. They perform chemoautotrophic ammonia oxidation and could thus be important partners in the biofilm-forming community, providing the source of nitrogen compounds to the other microorganisms [
34].
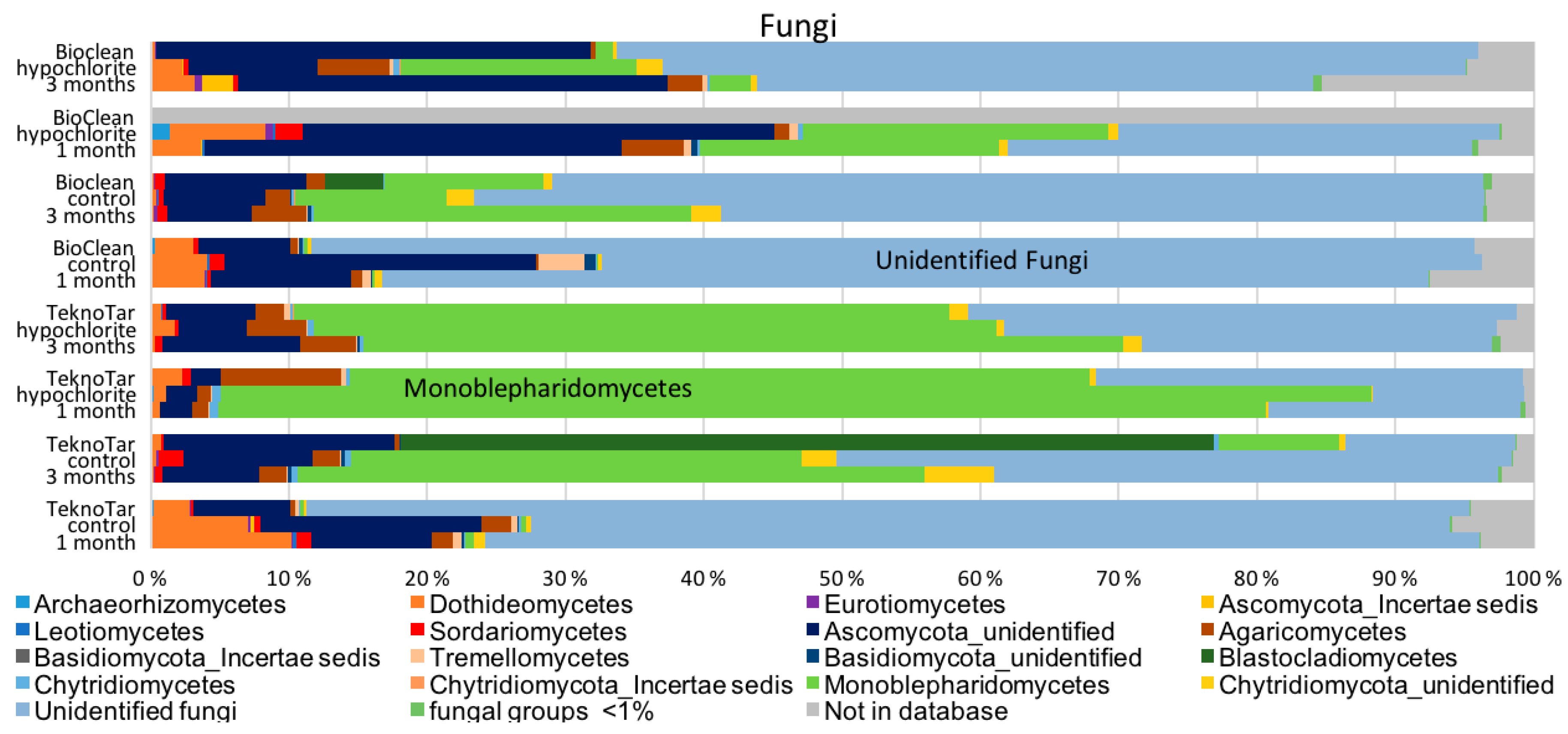
Monoblepharidomycetes and unidentified Ascomycota were the dominant fungi detected. Most aquatic fungi belong to Ascomycetes and Chytridiomycetes, as do the fungi detected here [
35]. Only small differences were detected in fungal communities between the control system and the hypochlorite-treated coupons. Similar fungal composition has been reported earlier from the same environment but different from current results; Monoblepharidomycetes were earlier detected only from the hypochlorite-treated environment [
31]. Fungi has been connected to deterioration of polymeric coatings [
36]. Fungi also exert physical forces, unlike bacteria or archaea, and have the ability to form hyphae, to penetrate hard substrates, and to spread on and through materials [
37]. Overall, fungi have an important role in the biofilm since they are able to degrade autochthonous and allochthonous organic matter [
35].
The biofouling and biodeterioration of man-made materials, including metal and their alloys, due to biofilm formation has a great environmental and economical implications [
38]. Antifouling coatings are widely used on marine structures to control biofouling [
39]. Both coatings included in this study were fouling release type of coatings [
9]. The effectiveness of most of the non-stick coatings is based on a combination of surface free energy, surface structure, and surface roughness [
40]. Here, the anti-fouling coatings alone did not prevent the settlement of microfouling organisms. Since microbial fouling affects the performance of commercial antifouling coatings the quantitative and qualitative analyses of marine microbial fouling communities is of great interest [
25,
39]. As summarized by Flemming [
4], detailed knowledge about biofilms is crucial for understanding and preventing biofouling, as well as to choose a successful anti-fouling strategy. Microorganisms may cause deterioration of polymeric substances, such as the coatings [
10]. However, during the relatively short survey time (three months) in this study, no degradation of coatings was evident. Even though the microfouling species would not directly cause deterioration of the coatings, the original surface properties, such as hydrophobicity, can be masked [
10].
The present study demonstrates that microfouling combined to inorganic fouling was extensive and formed by a diverse group of microorganisms in a cooling water cycle utilizing brackish Baltic Sea water. The composition of a microfouling community, not only the number of microorganisms, has been identified to be an important factor in either promoting or inhibiting the macrofouling settlement depending on the species composition [
3]. The hypochlorite treatment combined to anti-fouling coatings proved to be efficient at least during this three-month survey period.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









