
Magnetic Properties and Initiation of Biogenic Reefs in Xisha Islands, South China Sea, at the Oligo–Miocene Boundary



Abstract
:1. Introduction
2. Materials and Methods
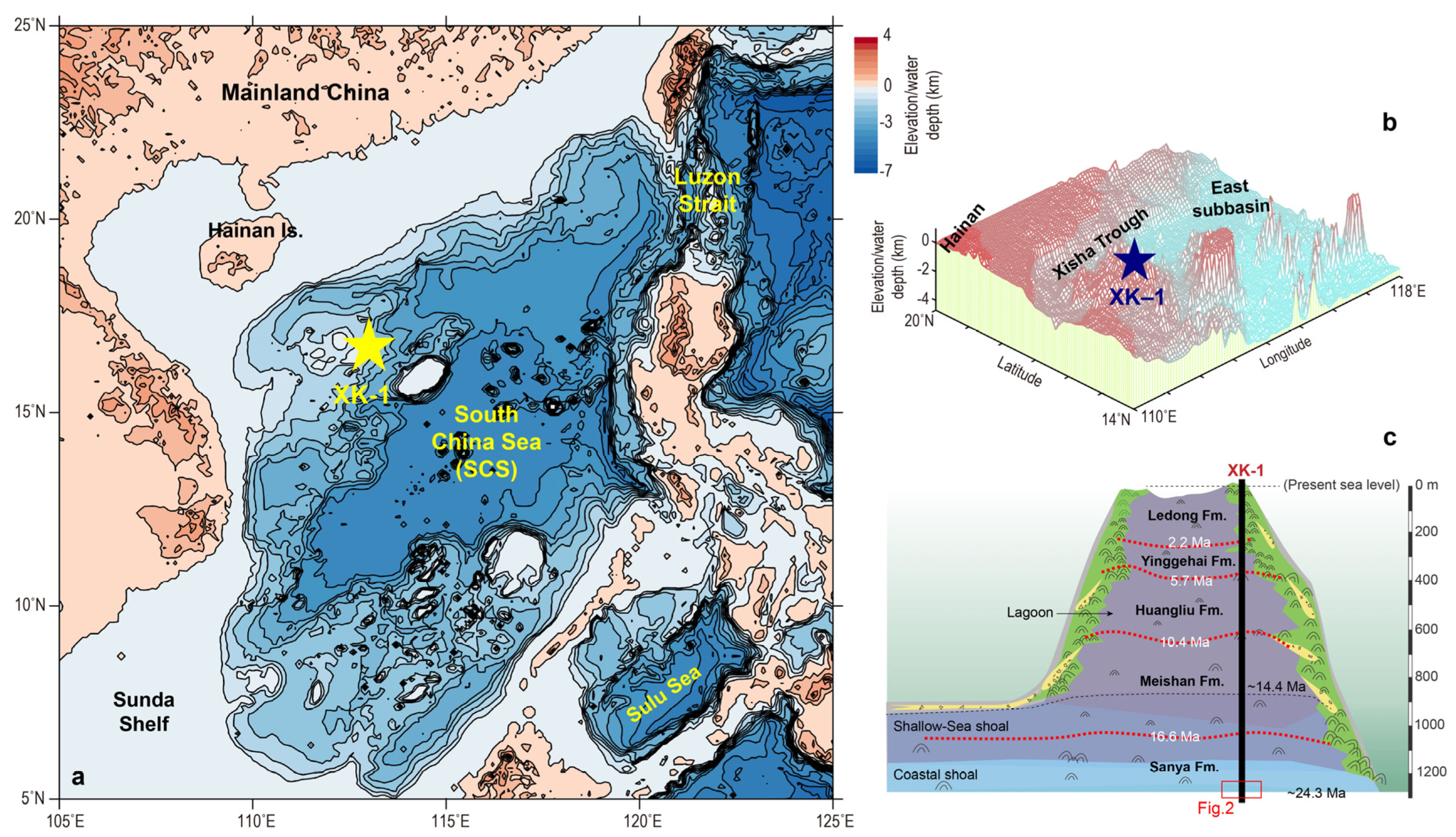
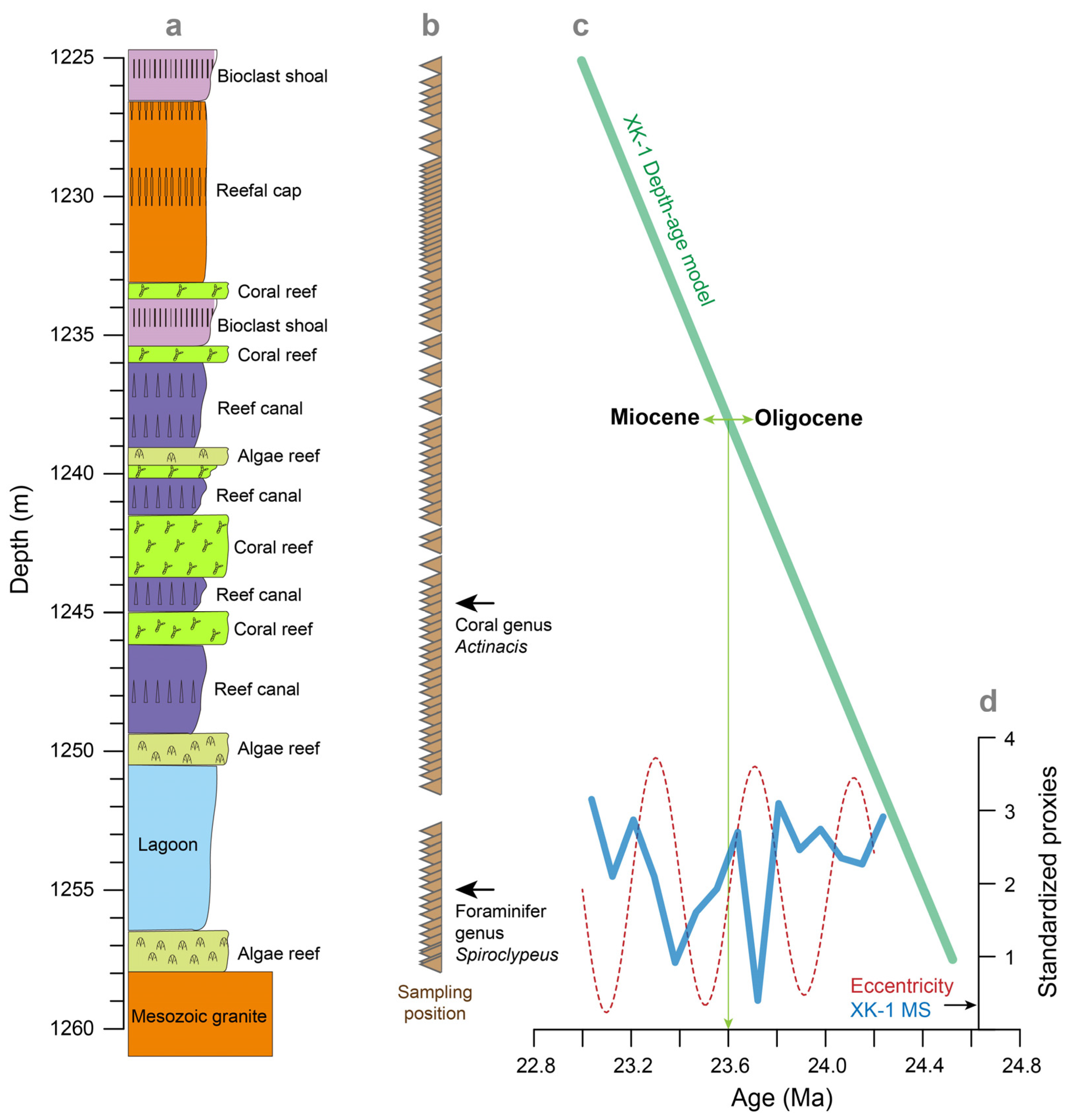
2.1. Core XK-1
2.2. Measurements
3. Results
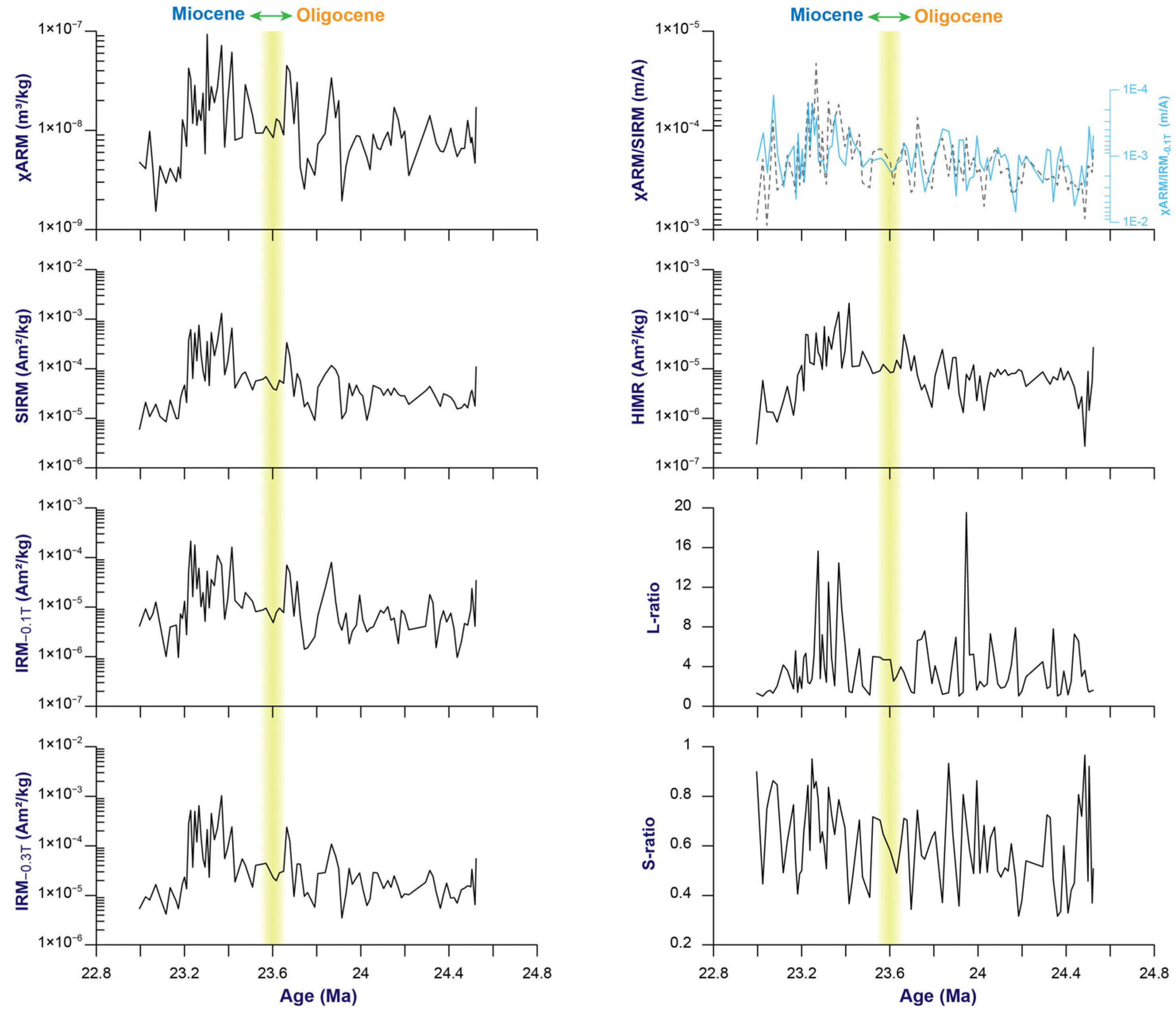
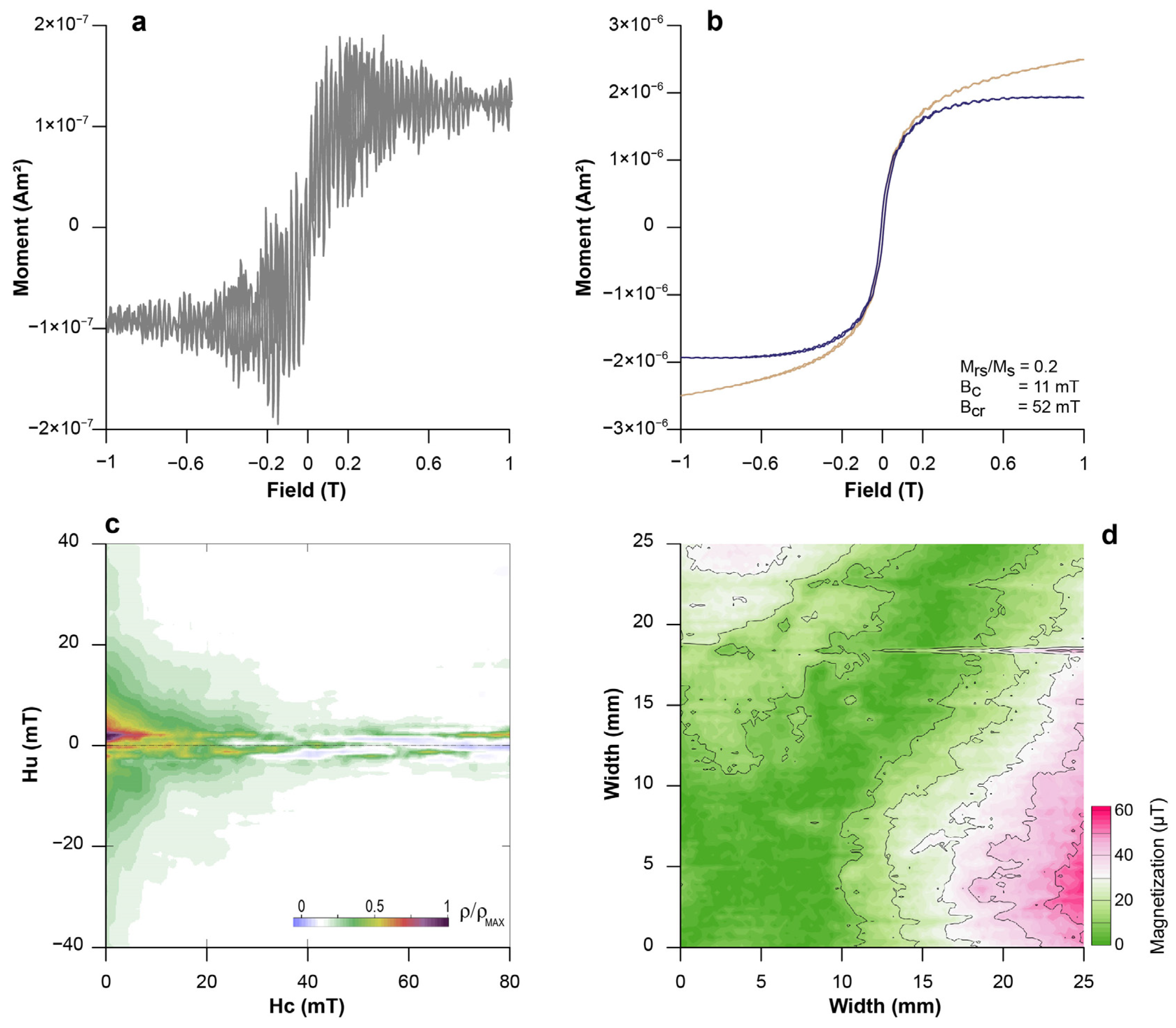
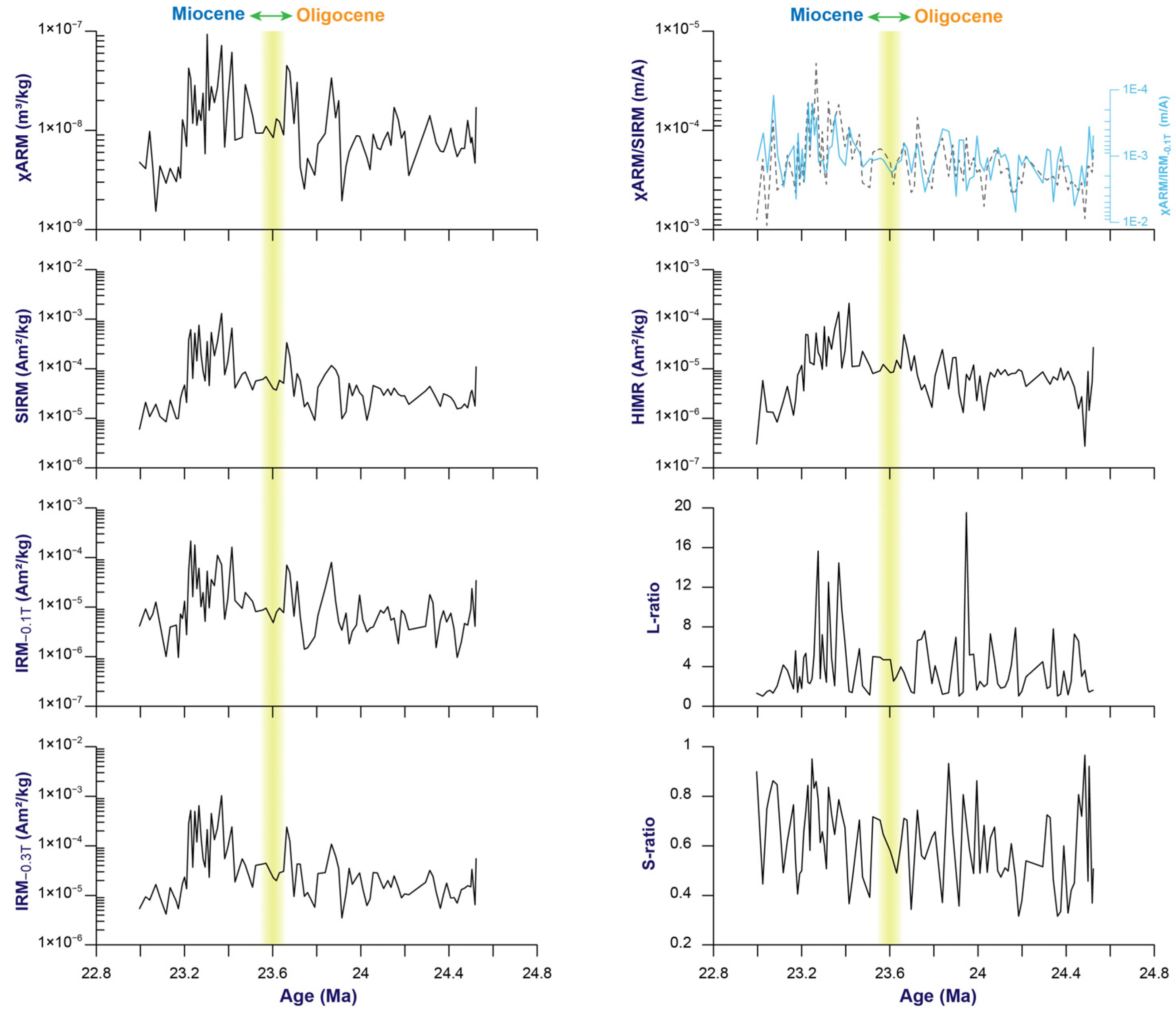
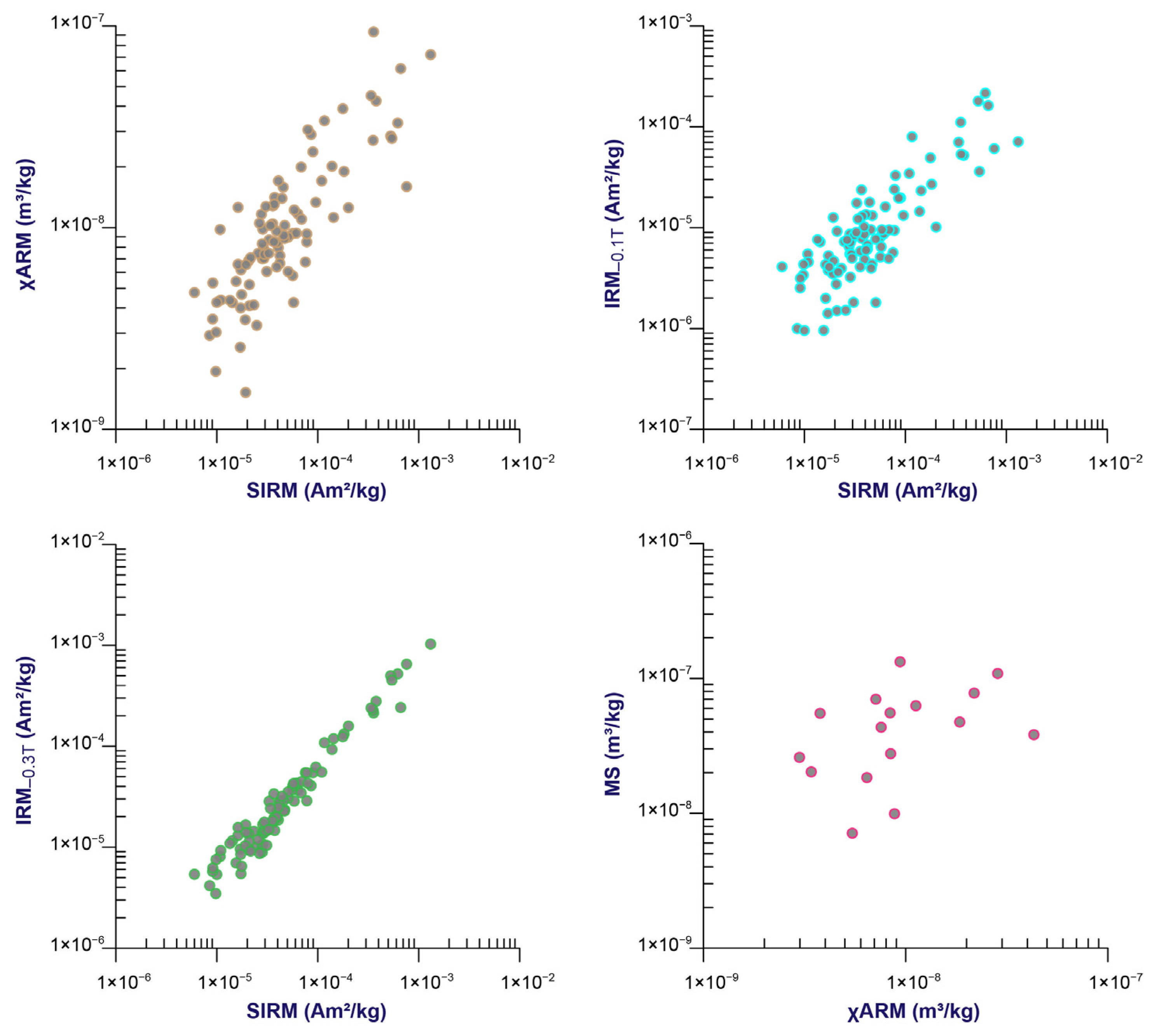
3.1. Changes in Magnetic Parameters
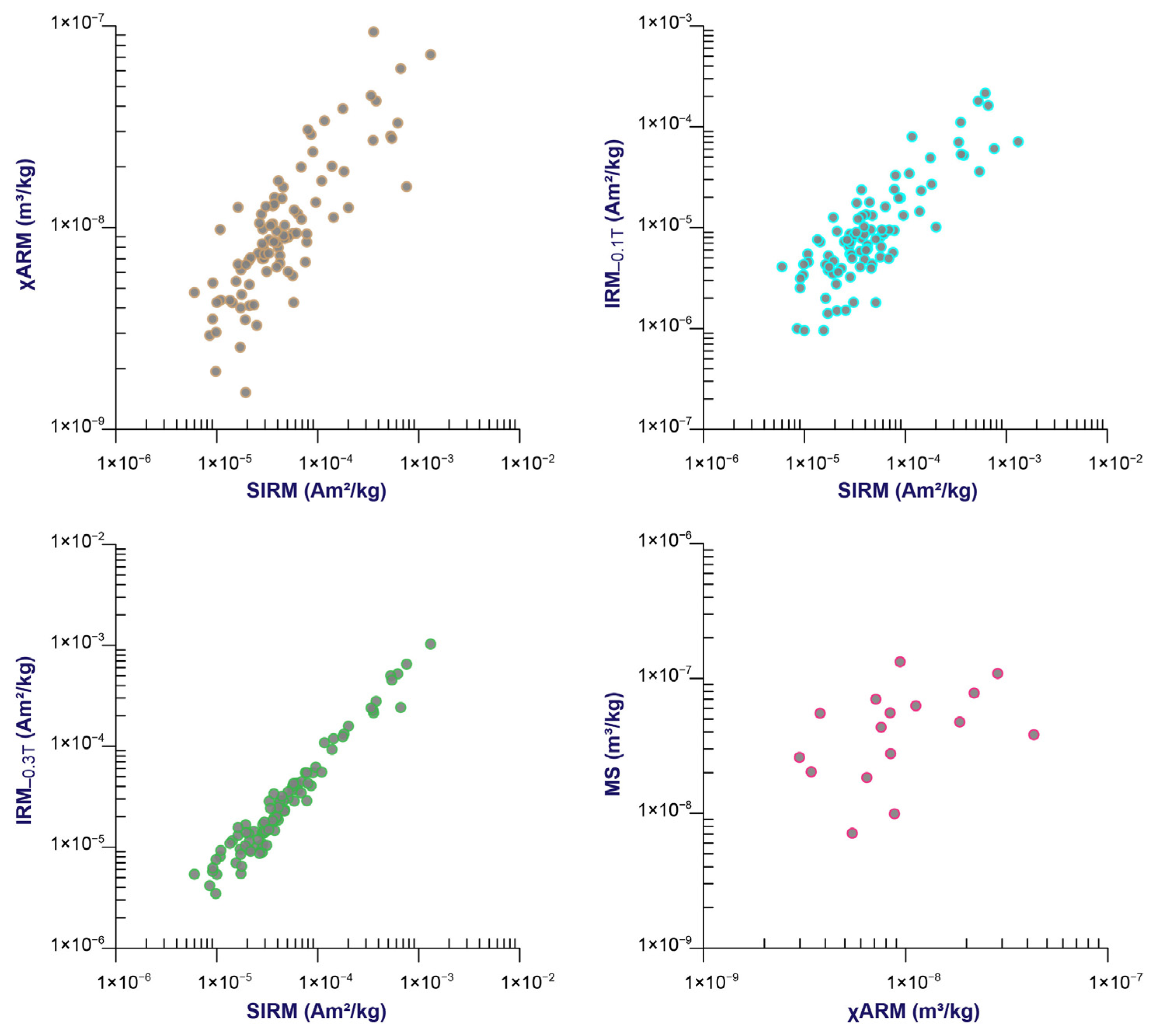
3.2. Magnetic Minerals and the Proxy
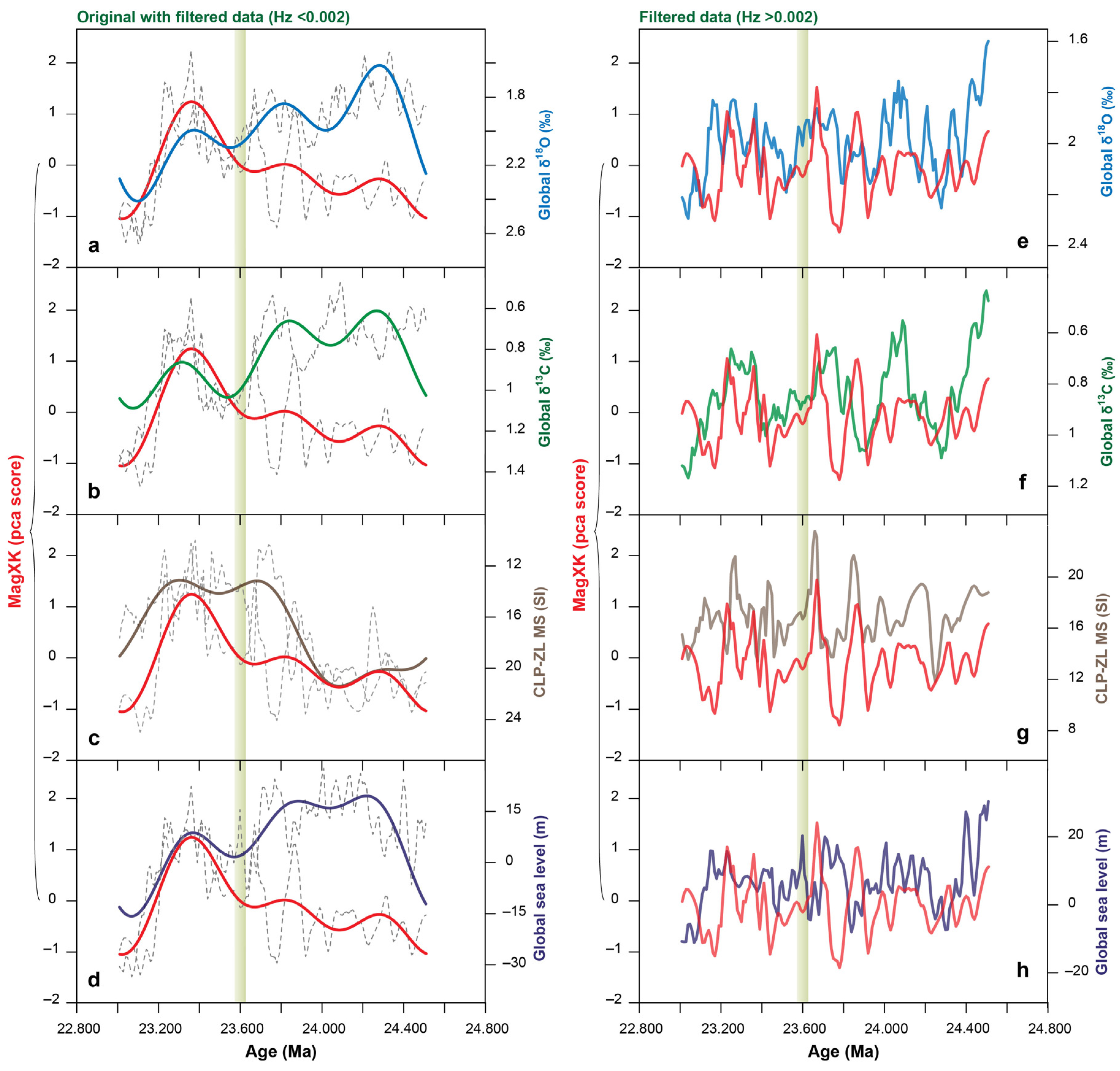
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Milliman, J.D. Production and accumulation of calcium carbonate in the ocean: Budget of a nonsteady state. Glob. Biogeochem. Cycles 1993, 7, 927–957. [Google Scholar] [CrossRef]
- Milliman, J.D.; Droxler, A.W. Neritic and pelagic carbonate sedimentation in the marine environment: Ignorance is not bliss. Geol. Rundsch. 1996, 85, 496–504. [Google Scholar] [CrossRef]
- Eyre, B.D.; Andersson, A.J.; Cyronak, T. Benthic coral reef calcium carbonate dissolution in an acidifying ocean. Nat. Clim. Chang. 2014, 4, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [Green Version]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Pandolfi, J. Incorporating Uncertainty in Predicting the Future Response of Coral Reefs to Climate Change. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 281–303. [Google Scholar] [CrossRef]
- Cumming, G.S.; Morrison, T.H.; Hughes, T.P. New Directions for Understanding the Spatial Resilience of Social–Ecological Systems. Ecosystems 2017, 20, 649–664. [Google Scholar] [CrossRef]
- Levin, S.; Xepapadeas, T.; Crépin, A.-S.; Norberg, J.; de Zeeuw, A.; Folke, C.; Hughes, T.; Arrow, K.; Barrett, S.; Daily, G.; et al. Social-ecological systems as complex adaptive systems: Modeling and policy implications. Environ. Dev. Econ. 2012, 18, 111–132. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.; Gardner, T.A.; Bennett, E.M.; Balvanera, P.; Biggs, R.; Carpenter, S.; Daw, T.; Folke, C.; Hill, R.; Hughes, T.P.; et al. Advancing sustainability through mainstreaming a social–ecological systems perspective. Curr. Opin. Environ. Sustain. 2015, 14, 144–149. [Google Scholar] [CrossRef]
- Martin, R.; Schlüter, M. Combining system dynamics and agent-based modeling to analyze social-ecological interactions—an example from modeling restoration of a shallow lake. Front. Environ. Sci. 2015, 3, 66. [Google Scholar] [CrossRef] [Green Version]
- Laborde, S.; Fernández, A.; Phang, S.C.; Hamilton, I.M.; Henry, N.; Jung, H.C.; Mahamat, A.; Ahmadou, M.; Labara, B.K.; Kari, S.; et al. Social-ecological feedbacks lead to unsustainable lock-in in an inland fishery. Glob. Environ. Chang. 2016, 41, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, J.; Pereira, H.M. Regime shifts in a socio-ecological model of farmland abandonment. Landsc. Ecol. 2011, 26, 737–749. [Google Scholar] [CrossRef]
- Ban, S.S.; Graham, N.A.J.; Connolly, S.R. Evidence for multiple stressor interactions and effects on coral reefs. Glob. Chang. Biol. 2014, 20, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.C.; Kleypas, J.; van de Leemput, I.A.; Lough, J.M.; Morrison, T.H.; et al. Coral reefs in the Anthropocene. Nature 2017, 546, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Quattrini, A.M.; Rodríguez, E.; Faircloth, B.C.; Cowman, P.F.; Brugler, M.R.; Farfan, G.A.; Hellberg, M.E.; Kitahara, M.V.; Morrison, C.L.; Paz-García, D.A.; et al. Palaeoclimate ocean conditions shaped the evolution of corals and their skeletons through deep time. Nat. Ecol. Evol. 2020, 4, 1531–1538. [Google Scholar] [CrossRef] [PubMed]
- Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; Billups, K. Trends, Rhythms, and Aberrations in Global Climate 65 Ma to Present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yang, Z.; Wang, D.; Lü, F.; Lüdmann, T.; Fulthorpe, C.; Wang, B. Architecture, development and geological control of the Xisha carbonate platforms, northwestern South China Sea. Mar. Geol. 2014, 350, 71–83. [Google Scholar] [CrossRef]
- Yi, L.; Jian, Z.; Liu, X.; Zhu, Y.; Zhang, D.; Wang, Z.; Deng, C. Astronomical tuning and magnetostratigraphy of Neogene biogenic reefs in Xisha Islands, South China Sea. Sci. Bull. 2018, 63, 564–573. [Google Scholar] [CrossRef] [Green Version]
- Fulthorpe, C.S.; Schlanger, S.O. Paleo-oceanographic and tectonic setting of Early Miocene reefs and associated carbonates offshore Southeast Asia. AAPG Bull. 1989, 73, 729–756. [Google Scholar]
- Sales, A.O.; Jacobsen, E.C.; Morado Jr, A.A.; Benavidez, J.J.; Navarro, F.A.; Lim, A.E. The petroleum potential of deep-water northwest Palawan Block GSEC 66. J. Asian Earth Sci. 1997, 15, 217–240. [Google Scholar] [CrossRef]
- Williams, H.H. Play concepts-northwest Palawan, Philippines. J. Asian Earth Sci. 1997, 15, 251–273. [Google Scholar] [CrossRef]
- Hutchison, C.S. Marginal basin evolution: The southern South China Sea. Mar. Pet. Geol. 2004, 21, 1129–1148. [Google Scholar] [CrossRef]
- Hutchison, C.S.; Vijayan, V.R. What are the Spratly Islands? J. Asian Earth Sci. 2010, 39, 371–385. [Google Scholar] [CrossRef]
- Ma, Y.; Wu, S.; Lv, F.; Dong, D.; Sun, Q.; Lu, Y.; Gu, M. Seismic characteristics and development of the Xisha carbonate platforms, northern margin of the South China Sea. J. Asian Earth Sci. 2011, 40, 770–783. [Google Scholar] [CrossRef]
- Yi, L.; Wang, Z.; Zhang, D.; Liu, X.; You, L.; Luo, W.; Zhu, Y.; Qin, H.; Deng, C. Magnetostratigraphy of biogenic reefs in the Sanya Formation of Borehole XK-1 from Xisha Islands and its environmental significance. Coast. Eng. 2016, 35, 1–11. (In Chinese) [Google Scholar]
- Zhang, M.S.; He, Q.X.; Ye, Z.J. The Geologic Research of Deposition of Bioherm Carbonate in the Xisha Islands; Science Press: Beijing, China, 1989. [Google Scholar]
- Zhao, H.T.; Sha, Q.A.; Zhu, Y.Z. Quaternary Coral Reef: Geology of Yongshu Reef, Nansha Islands; China Ocean Press: Beijing, China, 1992; p. 264. [Google Scholar]
- Zhu, Y.Z.; Sha, Q.A.; Guo, L.F.; Yu, K.F.; Zhao, H.T. Cenozoic Coral Reef Geology of Yongshu Reef, Nansha Islands; Science Press: Beijing, China, 1997; p. 137. [Google Scholar]
- Xu, S.; Yu, K.; Fan, T.; Jiang, W.; Wang, R.; Zhang, Y.; Yue, Y.; Wang, S. Coral reef carbonate δ13C records from the northern South China Sea: A useful proxy for seawater δ13C and the carbon cycle over the past 1.8 Ma. Glob. Planet. Chang. 2019, 182, 103003. [Google Scholar] [CrossRef]
- Yi, L.; Deng, C.; Yan, W.; Wu, H.; Zhang, C.; Xu, W.; Su, X.; He, H.; Guo, Z. Neogene–quaternary magnetostratigraphy of the biogenic reef sequence of core NK–1 in Nansha Qundao, South China Sea. Sci. Bull. 2021, 66, 200–203. [Google Scholar] [CrossRef]
- Qiao, P.; Zhu, W.; Shao, L.; Zhang, D.; Cheng, X.; Song, Y. Carbonate stable isotope stratigraphy of Well Xike-1, Xisha Islands. J. China Univ. Geosci. 2015, 40, 725–732. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, D.; Liu, X.; You, L.; Luo, W.; Yi, L.; Zhu, Y.; Qin, H.; Xie, Q.; Che, Z.; et al. Preliminary results of rock magnetism and magnetostratigraphy for Late Miocene to Pliocene biogenic reefs in the Xisha Islands, South China Sea. Chin. J. Geophys. 2016, 59, 4178–4187. [Google Scholar] [CrossRef]
- Lund, S.; Platzman, E.; Thouveny, N.; Camoin, G.; Corsetti, F.; Berelson, W. Biological control of paleomagnetic remanence acquisition in carbonate framework rocks of the Tahiti coral reef. Earth Planet. Sci. Lett. 2010, 298, 14–22. [Google Scholar] [CrossRef]
- Wei, X.; Deng, J.F.; Xie, W.Y.; Zhu, Y.J.; Zhao, G.C.; Li, Y.X.; Chen, Y.H. Constraints on biogenetic reef formation during evolution of the South China Sea and exploration potential analysis. Earth Geosci. Front. 2005, 12, 245–252. [Google Scholar]
- Wang, P.; Li, Q. The South China Sea: Paleoceanography and Sedimentology; Springer: Dordrecht, The Netherlands, 2009; Volume 13, p. 506. [Google Scholar]
- Xu, G.Q.; Lv, B.Q.; Wang, H.G. Drown event research: Insight from Cenozoic carbonate platform in northern South China Sea. J. Tongji Univ. 2002, 30, 35–40. [Google Scholar]
- Fyhn, M.B.W.; Boldreel, L.O.; Nielsen, L.H.; Giang, T.C.; Nga, L.H.; Hong, N.T.M.; Nguyen, N.D.; Abatzis, I. Carbonate platform growth and demise offshore Central Vietnam: Effects of Early Miocene transgression and subsequent onshore uplift. J. Asian Earth Sci. 2013, 76, 152–168. [Google Scholar] [CrossRef]
- Zhu, W.; Wang, Z.; Mi, L.; Du, X.; Xie, X.; Lu, Y.; Zhang, D.; Sun, Z.; Liu, X.; You, L. Sequence stratigraphic framework and reef growth unit of Well Xike-1 from Xisha Islands, South China Sea. J. China Univ. Geosicnes 2015, 40, 677–687. [Google Scholar]
- Luo, W.; Zhang, D.; Liu, X.; Wang, Z.; Hu, W.; Wang, Y. A comprehensive stratigraphic study of Well XK-1 in the Xisha Area. J. Stratigr. 2018, 42, 485–498. [Google Scholar]
- Wang, Z.; Zhang, D.; Liu, X.; You, L.; Luo, W.; Yi, L.; Tan, L.; Zhu, Y.; Qin, H.; Cheng, H.; et al. Magnetostratigraphy and 230Th dating of Pleistocene biogenic reefs in XK-1 borehole from Xisha Islands, South China Sea. Chin. J. Geophys. 2017, 60, 1027–1038. (In Chinese) [Google Scholar] [CrossRef]
- Wu, F.; Xie, X.; Zhu, Y.; Chen, B.; Shang, Z. Sequence stratigraphy of the Late Oligocene carbonate system on the Xisha Islands in the South China Sea. Int. J. Earth Sci. 2021, 110, 1611–1629. [Google Scholar] [CrossRef]
- Tauxe, L. Essentials of Paleomagnetism; University of California Press: Berkeley, CA, USA, 2010. [Google Scholar]
- Heslop, D. Numerical strategies for magnetic mineral unmixing. Earth-Sci. Rev. 2015, 150, 256–284. [Google Scholar] [CrossRef]
- Heslop, D.; Roberts, A.P. Unmixing Magnetic Hysteresis Loops. J. Geophys. Res. Atmos. 2012, 117, 3758. [Google Scholar] [CrossRef]
- Kletetschka, G.; Schnabl, P.; Šifnerová, K.; Tasáryová, Z.; Manda, Š.; Pruner, P. Magnetic scanning and interpretation of paleomagnetic data from Prague Synform’s volcanics. Studia Geophys. Geod. 2013, 57, 103–117. [Google Scholar] [CrossRef]
- Duan, Z.; Gao, X.; Liu, Q. Anhysteretic remanent magnetization (ARM) and its application to geoscience. Prog. Geophys. 2012, 27, 1929–1938. [Google Scholar] [CrossRef]
- Maher, B.A. Magnetic properties of some synthetic sub-micron magnetites. Geophys. J. 1988, 94, 83–96. [Google Scholar] [CrossRef]
- Evans, M.E.; Heller, F. Environmental Magnetism: Principles and Applications of Enviromagnetics; Academic Press: New York, NY, USA, 2003; p. 322. [Google Scholar]
- Yi, L.; Wang, H.; Liu, G.; Chen, Y.; Yao, H.; Deng, X. Magnetic minerals in Mid-Pleistocene sediments on the Caiwei Guyot, Northwest Pacific and their response to the Mid-Brunhes climate event. Acta Oceanol. Sin. 2021, 40, 253. [Google Scholar]
- Yang, D.; Wang, M.; Lu, H.; Ding, Z.; Liu, J.; Yan, C. Magnetic properties and correlation with heavy metals in mangrove sediments, the case study on the coast of Fujian, China. Mar. Pollut. Bull. 2019, 146, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Appel, E.; Fang, X.; Song, C.; Cirpka, O. Magnetostratigraphy of deep drilling core SG-1 in the western Qaidam Basin (NE Tibetan Plateau) and its tectonic implications. Quat. Res. 2012, 78, 139–148. [Google Scholar] [CrossRef]
- Roberts, A.P.; Heslop, D.; Zhao, X.; Pike, C.R. Understanding fine magnetic particle systems through use of first-order reversal curve diagrams. Rev. Geophys. 2014, 52, 557–602. [Google Scholar] [CrossRef]
- Roberts, A.P.; Pike, C.R.; Verosub, K.L. First-order reversal curve diagrams: A new tool for characterizing the magnetic properties of natural samples. J. Geophys. Res. Solid Earth 2000, 105, 28461–28475. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Liu, J.; Zhang, D.; Li, X.; Shi, Z. Characteristics and significance of manganese minerals in dolostones from Well Xike-1, Xisha Area, South China Sea. Mineral. Petrol. 2020, 40, 81–91. [Google Scholar]
- Laskar, J.; Fienga, A.; Gastineau, M.; Manche, H. La2010: A new orbital solution for the long-term motion of the Earth. Astron. Astrophys. 2011, 532, A89. [Google Scholar] [CrossRef]
- Grinsted, A.; Moore, J.C.; Jevrejeva, S. Application of the cross wavelet transform and wavelet coherence to geophysical time series. Nonlinear Process. Geophys. 2004, 11, 561–566. [Google Scholar] [CrossRef]
- Bachtel, S.L.; Kissling, R.D.; Martono, D.; Rahardjanto, S.P.; Dunn, P.A.; MacDonald, B.A. Seismic Stratigraphic Evolution of the Miocene–Pliocene Segitiga Platform, East Natuna Sea, Indonesia: The Origin, Growth Ad Demise of an Isolated Carbonate Platform. In Seismic Imaging of Carbonate Reservoir and Systems; Eberli, G.P., Massaferro, J.L., Sarg, J.F., Eds.; AAPG Memoir: Tulsa, OK, USA, 2003; Volume 81, pp. 309–328. [Google Scholar]
- Belopolsky, A.V.; Droxler, A.W. Imaging Tertiary carbonate systems, the Maldives, Indian Ocean: Insights into carbonate sequence interpretation. Lead. Edge 2003, 22, 646–652. [Google Scholar] [CrossRef]
- Epting, M. The Miocene carbonate buildups of central Luconia, offshore Sarawak. In Atlas of Seismic Stratigraphy; Bally, A.W., Ed.; AAPG: Tulsa, OK, USA, 1989; pp. 168–173. [Google Scholar]
- Wilson, M.E.J. Cenozoic carbonates in Southeast Asia: Implications for equatorial carbonate development. Sediment. Geol. 2002, 147, 295–428. [Google Scholar] [CrossRef]
- Qiang, X.; An, Z.; Song, Y.; Chang, H.; Sun, Y.; Liu, W.; Ao, H.; Dong, J.; Fu, C.; Wu, F.; et al. New eolian red clay sequence on the western Chinese Loess Plateau linked to onset of Asian desertification about 25 Ma ago. Sci. China Earth Sci. 2011, 54, 136–144. [Google Scholar] [CrossRef]
- Miller, K.G.; Browning, J.V.; Schmelz, W.J.; Kopp, R.E.; Mountain, G.S.; Wright, J.D. Cenozoic sea-level and cryospheric evolution from deep-sea geochemical and continental margin records. Sci. Adv. 2020, 6, eaaz1346. [Google Scholar] [CrossRef] [PubMed]
- Lisiecki, L.E. A benthic δ13C-based proxy for atmospheric pCO2 over the last 1.5 Myr. Geophys. Res. Lett. 2010, 37, L21708. [Google Scholar] [CrossRef] [Green Version]
- Crook, E.D.; Cohen, A.L.; Rebolledo-Vieyra, M.; Hernandez, L.; Paytan, A. Reduced calcification and lack of acclimatization by coral colonies growing in areas of persistent natural acidification. Proc. Natl. Acad. Sci. USA 2013, 110, 11044–11049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fantazzini, P.; Mengoli, S.; Pasquini, L.; Bortolotti, V.; Brizi, L.; Mariani, M.; Di Giosia, M.; Fermani, S.; Capaccioni, B.; Caroselli, E.; et al. Gains and losses of coral skeletal porosity changes with ocean acidification acclimation. Nat. Commun. 2015, 6, 7785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollica, N.R.; Guo, W.; Cohen, A.L.; Huang, K.F.; Foster, G.L.; Donald, H.K.; Solow, A.R. Ocean acidification affects coral growth by reducing skeletal density. Proc. Natl. Acad. Sci. USA 2018, 115, 1754–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tambutté, E.; Venn, A.A.; Holcomb, M.; Segonds, N.; Techer, N.; Zoccola, D.; Allemand, D.; Tambutté, S. Morphological plasticity of the coral skeleton under CO2-driven seawater acidification. Nat. Commun. 2015, 6, 7368. [Google Scholar] [CrossRef] [Green Version]
- Foster, T.; Falter, J.; McCulloch, M.; Clode, P. Ocean acidification causes structural deformities in juvenile coral skeletons. Sci. Adv. 2016, 2, e1501130. [Google Scholar] [CrossRef] [Green Version]
- Frölicher, T.L.; Fischer, E.M.; Gruber, N. Marine heatwaves under global warming. Nature 2018, 560, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Smale, D.A.; Wernberg, T.; Oliver, E.C.J.; Thomsen, M.; Harvey, B.P.; Straub, S.C.; Burrows, M.T.; Alexander, L.V.; Benthuysen, J.A.; Donat, M.G.; et al. Marine heatwaves threaten global biodiversity and the provision of ecosystem services. Nat. Clim. Chang. 2019, 9, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C.; et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Enochs, I.C.; Manzello, D.P.; Donham, E.M.; Kolodziej, G.; Okano, R.; Johnston, L.; Young, C.; Iguel, J.; Edwards, C.B.; Fox, M.D.; et al. Shift from coral to macroalgae dominance on a volcanically acidified reef. Nat. Clim. Chang. 2015, 5, 1083–1088. [Google Scholar] [CrossRef]
- Albright, R.; Caldeira, L.; Hosfelt, J.; Kwiatkowski, L.; Maclaren, J.K.; Mason, B.M.; Nebuchina, Y.; Ninokawa, A.; Pongratz, J.; Ricke, K.L.; et al. Reversal of ocean acidification enhances net coral reef calcification. Nature 2016, 531, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.; O’Leary, R.; Lough, J. Growth of Western Australian Corals in the Anthropocene. Science 2012, 335, 593–596. [Google Scholar] [CrossRef] [Green Version]
- Betzler, C.; Eberli, G.P.; Kroon, D.; Wright, J.D.; Swart, P.K.; Nath, B.N.; Alvarez-Zarikian, C.A.; Alonso-García, M.; Bialik, O.M.; Blättler, C.L.; et al. The abrupt onset of the modern South Asian Monsoon winds. Sci. Rep. 2016, 6, 29838. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Ruddiman, W.F.; Hao, Q.; Wu, H.; Qiao, Y.; Zhu, R.; Peng, S.; Wei, J.J.; Yuan, B.; Liu, T.S. Onset of Asian desertification by 22 Myr ago inferred from loess deposits in China. Nature 2002, 416, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Clemens, S.C.; An, Z.; Yu, Z. Astronomical timescale and palaeoclimatic implication of stacked 3.6-Myr monsoon records from the Chinese Loess Plateau. Quat. Sci. Rev. 2006, 25, 33–48. [Google Scholar] [CrossRef]
- Huang, H.; Dong, Z.; Lian, J. Establishment of nature reserve of coral reef ecosystem on the Xisha Islands. Trop. Geogr. 2008, 28, 540–544. [Google Scholar]
- Edinger, E.N.; Risk, M.J. Oligocene-Miocene extinction and geographic restriction of Caribbean corals: Roles of turbidity, temperature, and nutrients. Palaios 1994, 9, 576–598. [Google Scholar] [CrossRef]
- Webster, J.M.; Braga, J.C.; Humblet, M.; Potts, D.C.; Iryu, Y.; Yokoyama, Y.; Fujita, K.; Bourillot, R.; Esat, T.M.; Fallon, S.; et al. Response of the Great Barrier Reef to sea-level and environmental changes over the past 30,000 years. Nat. Geosci. 2018, 11, 426–432. [Google Scholar] [CrossRef] [Green Version]
- Kiessling, W.; Simpson, C.; Beck, B.; Mewis, H.; Pandolfi, J.M. Equatorial decline of reef corals during the last Pleistocene interglacial. Proc. Natl. Acad. Sci. USA 2012, 109, 21378–21383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Edwards, R.L.; Sinha, A.; Spötl, C.; Yi, L.; Chen, S.; Kelly, M.; Kathayat, G.; Wang, X.; Li, X.; et al. The Asian monsoon over the past 640,000 years and ice age terminations. Nature 2016, 534, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D.; Ogg, G.M. Geologic Time Scale 2020; Elsevier: Amsterdam, The Netherlands, 2020; p. 1357. [Google Scholar] [CrossRef]
- Naish, T.; Powell, R.; Levy, R.; Wilson, G.; Scherer, R.; Talarico, F.; Krissek, L.; Niessen, F.; Pompilio, M.; Wilson, T. Obliquity-paced Pliocene West Antarctic ice sheet oscillations. Nature 2009, 458, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Gao, Y.; Ma, C.; Jones, M.M.; Zeeden, C.; Ibarra, D.E.; Wu, H.; Wang, C. Organic carbon burial is paced by a ~173-ka obliquity cycle in the middle to high latitudes. Sci. Adv. 2021, 7, eabf9489. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
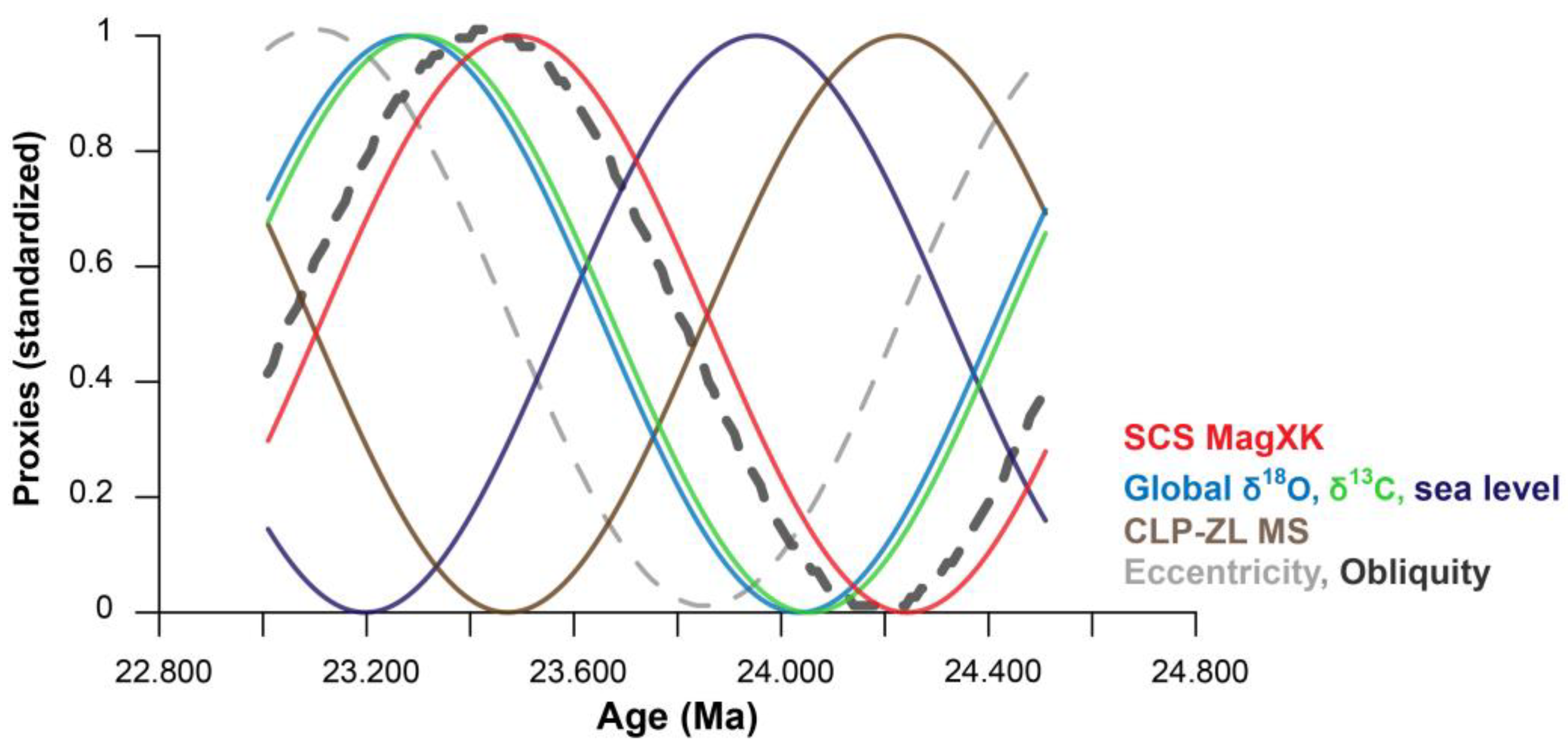{kind=link}
| Component 1 | Extraction Sums of Squared Loadings | ||
|---|---|---|---|
| Total | % of Variance | Cumulative % | |
| MagXK | 3.51 | 87.77 | 83.77 |
| Not included | 0.25 | 6.27 | 94.03 |
| 0.22 | 5.39 | 99.42 | |
| 0.02 | 0.58 | 100 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Liu, X.; Chen, W.; Yi, L. Magnetic Properties and Initiation of Biogenic Reefs in Xisha Islands, South China Sea, at the Oligo–Miocene Boundary. J. Mar. Sci. Eng. 2021, 9, 1031. https://doi.org/10.3390/jmse9091031
Li Y, Liu X, Chen W, Yi L. Magnetic Properties and Initiation of Biogenic Reefs in Xisha Islands, South China Sea, at the Oligo–Miocene Boundary. Journal of Marine Science and Engineering. 2021; 9(9):1031. https://doi.org/10.3390/jmse9091031
Chicago/Turabian StyleLi, Yibing, Xinyu Liu, Weiwei Chen, and Liang Yi. 2021. "Magnetic Properties and Initiation of Biogenic Reefs in Xisha Islands, South China Sea, at the Oligo–Miocene Boundary" Journal of Marine Science and Engineering 9, no. 9: 1031. https://doi.org/10.3390/jmse9091031
APA StyleLi, Y., Liu, X., Chen, W., & Yi, L. (2021). Magnetic Properties and Initiation of Biogenic Reefs in Xisha Islands, South China Sea, at the Oligo–Miocene Boundary. Journal of Marine Science and Engineering, 9(9), 1031. https://doi.org/10.3390/jmse9091031