Cellular Responses of Human Lymphatic Endothelial Cells to Carbon Nanomaterials


 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Carbon Nanomaterials and Their Dispersion
2.2. Cell Culture
2.3. Cell Viability
2.4. Inflammatory Response
2.5. Cellular Uptake
2.5.1. Observation of Cells by Fluorescence Microscopy
2.5.2. Observation of Cells by TEM
2.5.3. Quantification of Intracellular CNMs
2.6. Statistical Analysis
3. Results
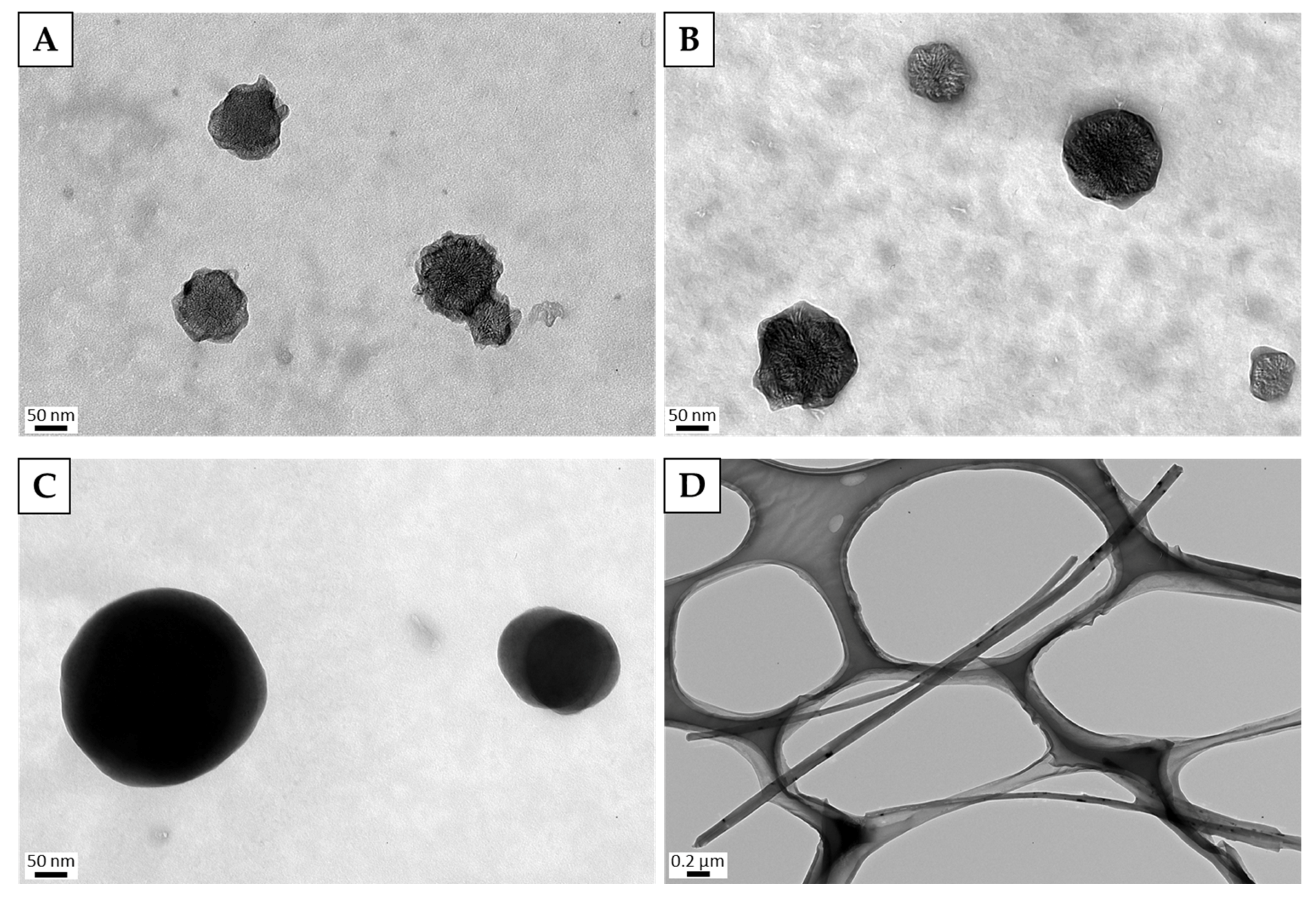
3.1. Characterization of Dispersed CNMs
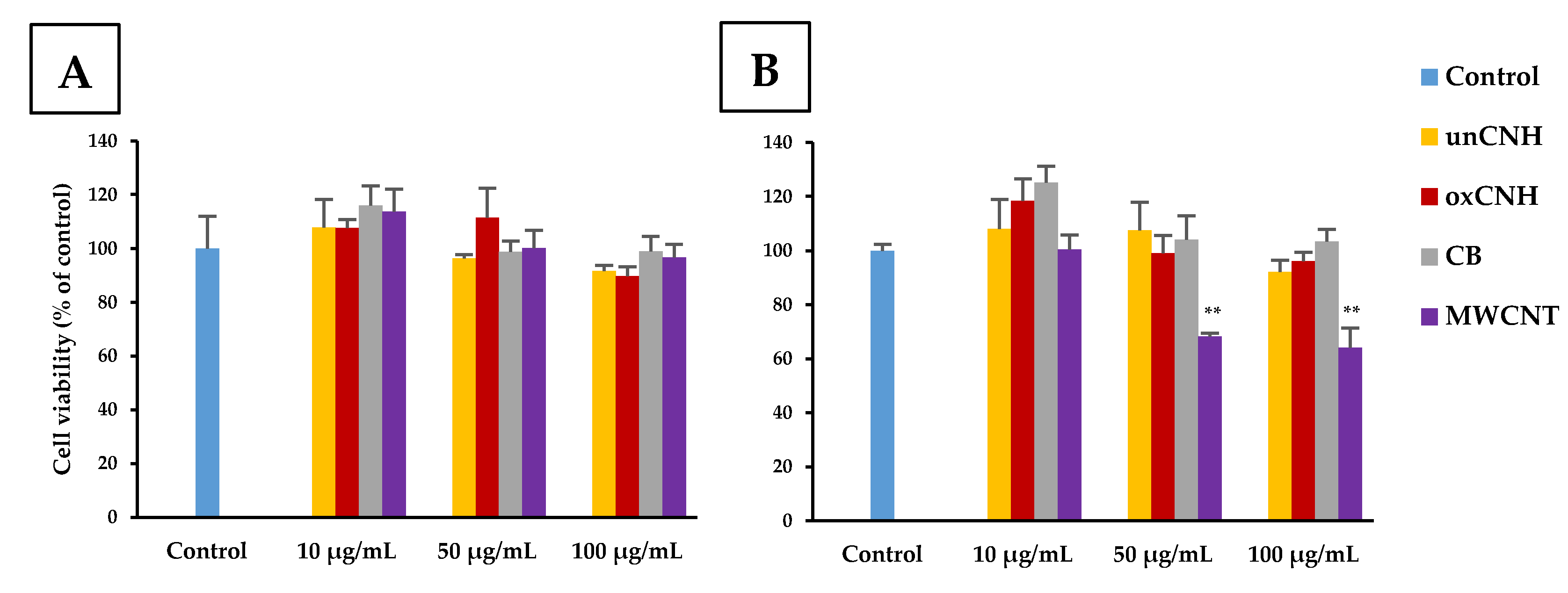
3.2. Cell Viability
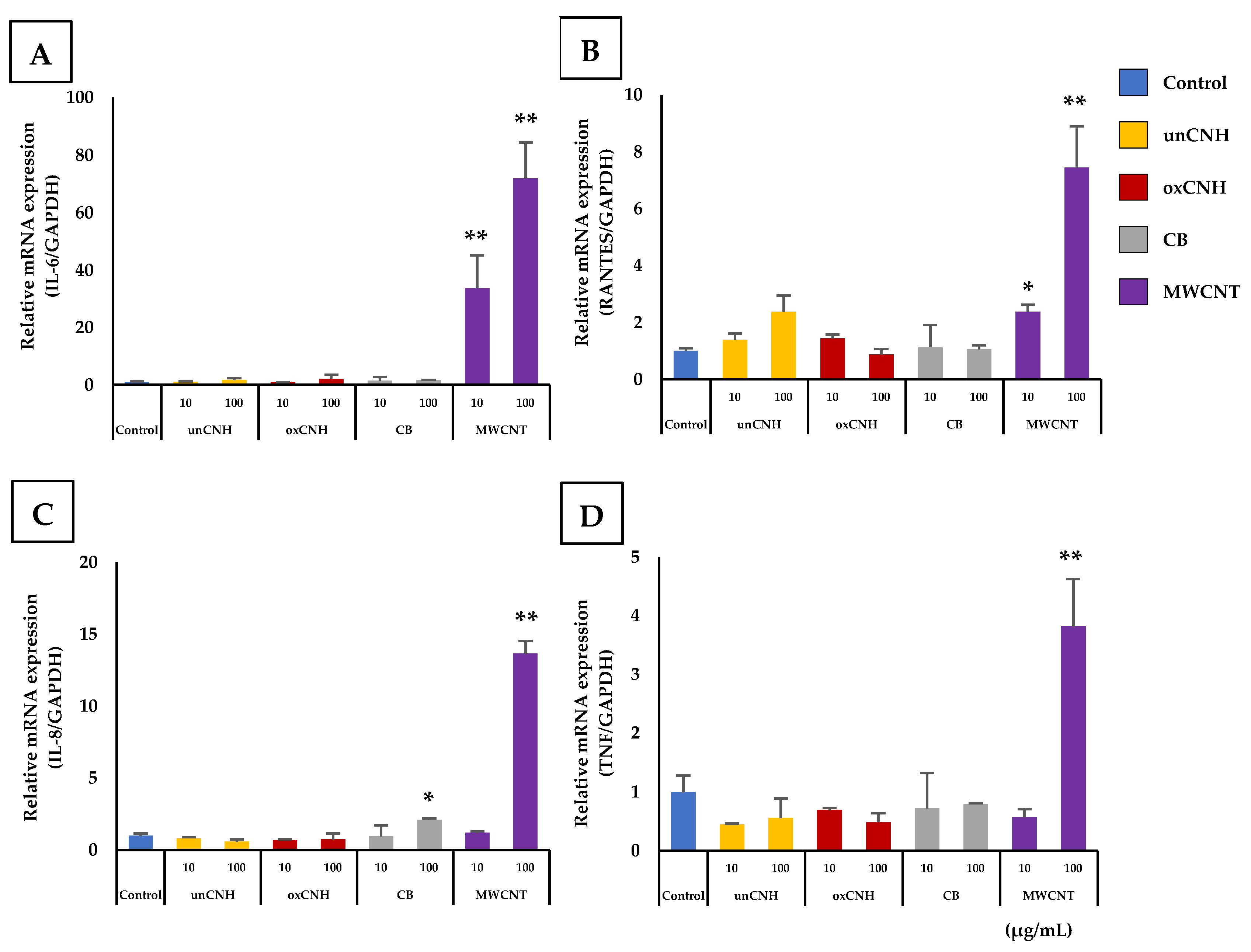
3.3. Inflammatory Responses
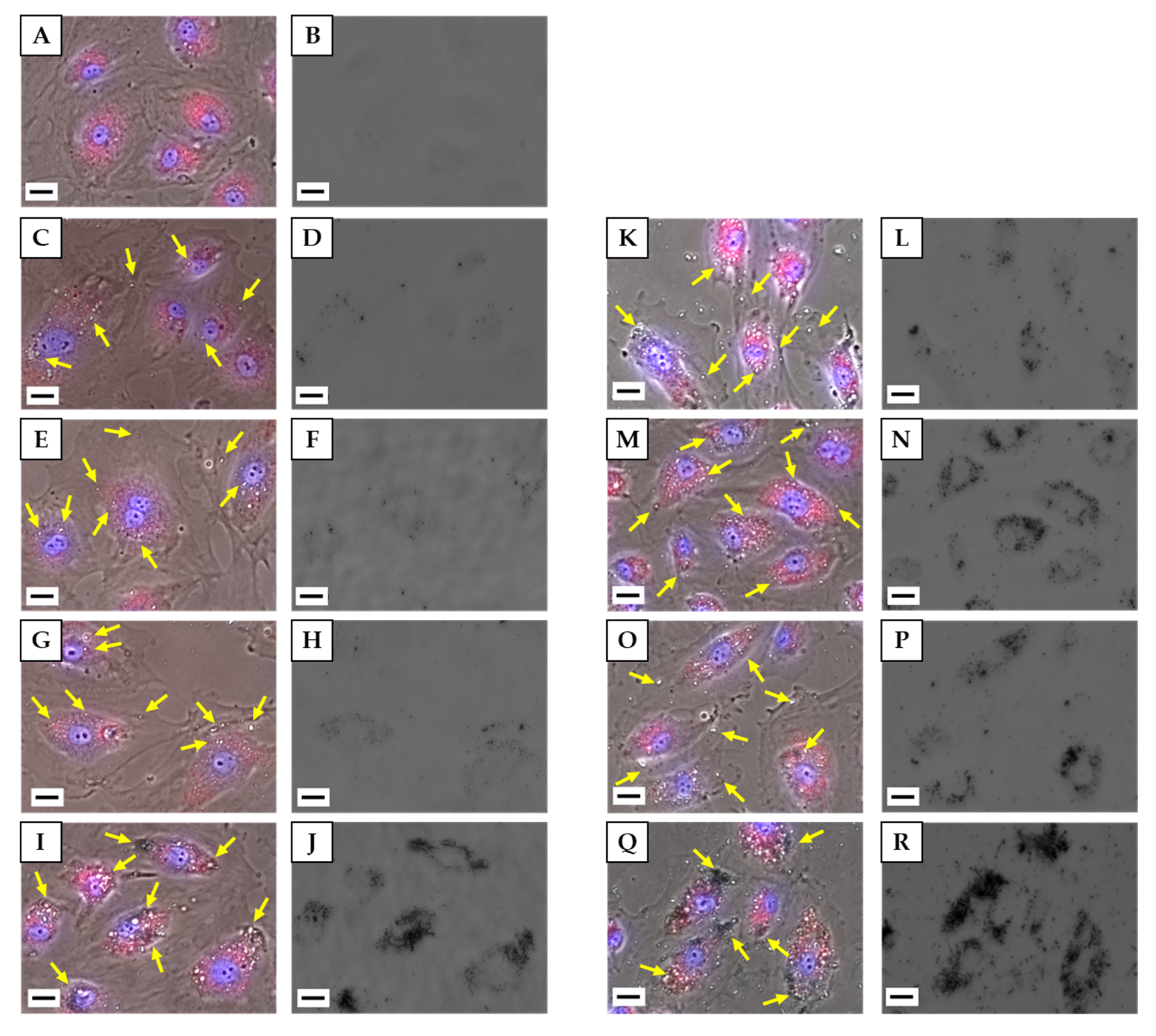
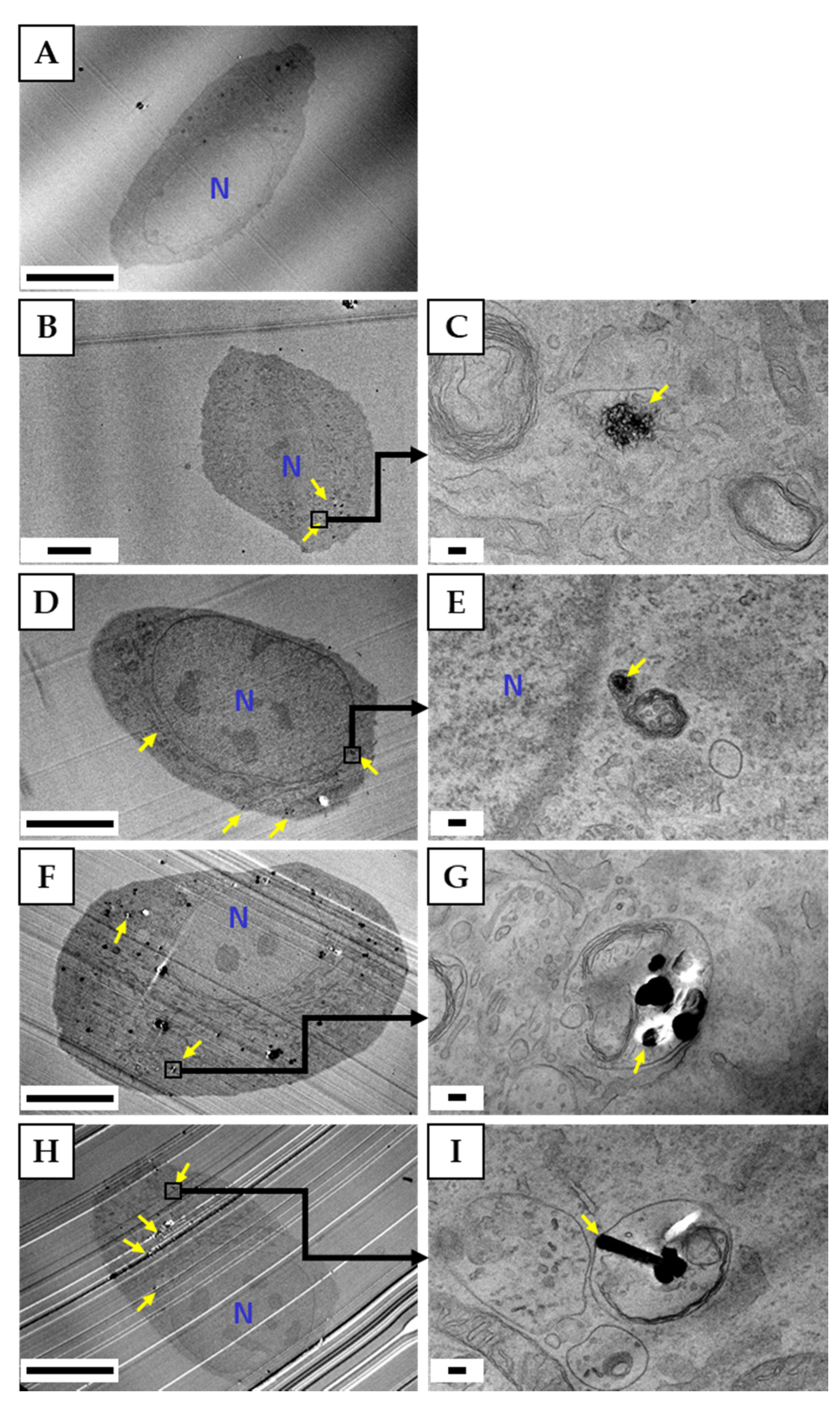
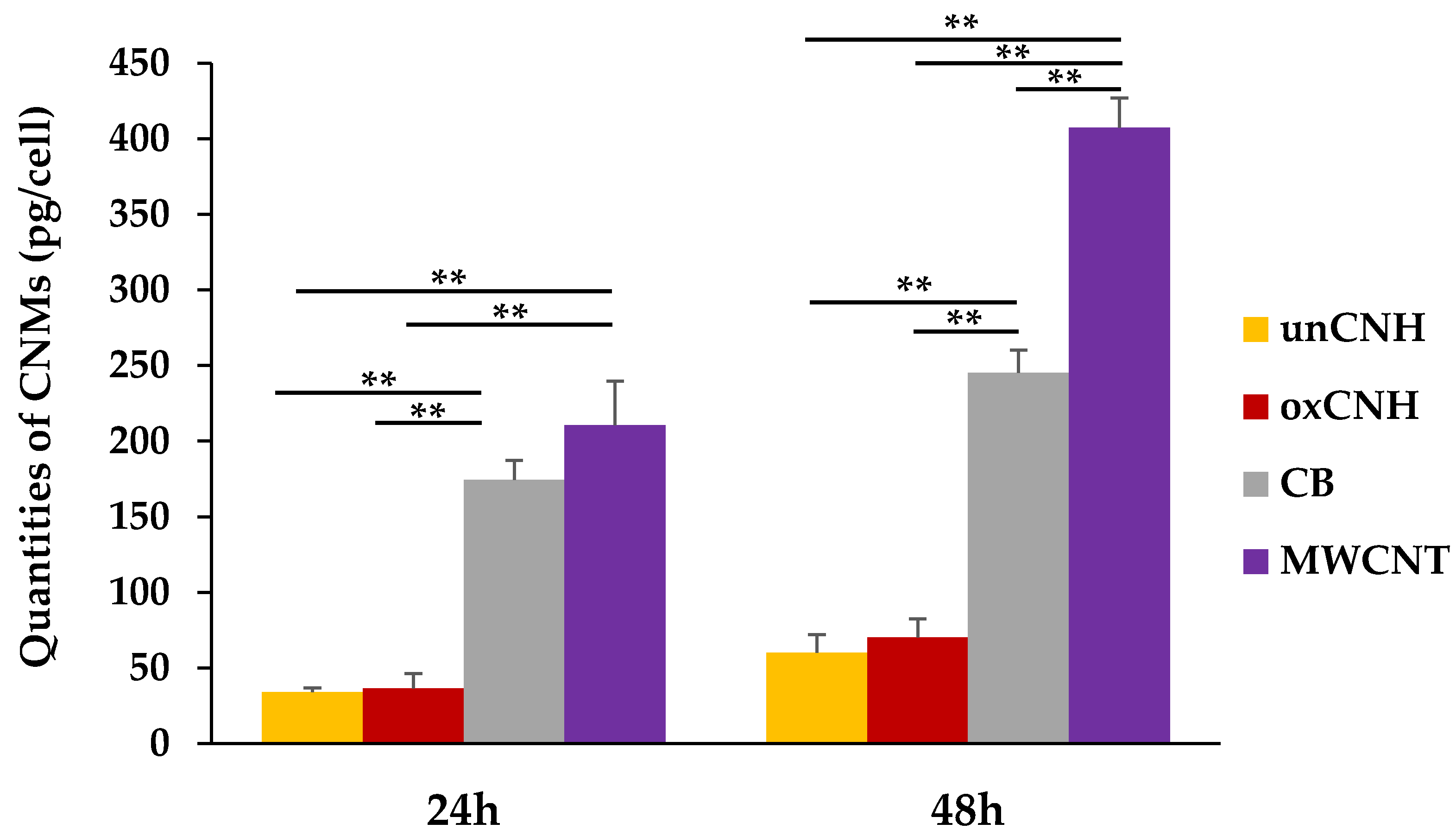
3.4. Cellular Uptake
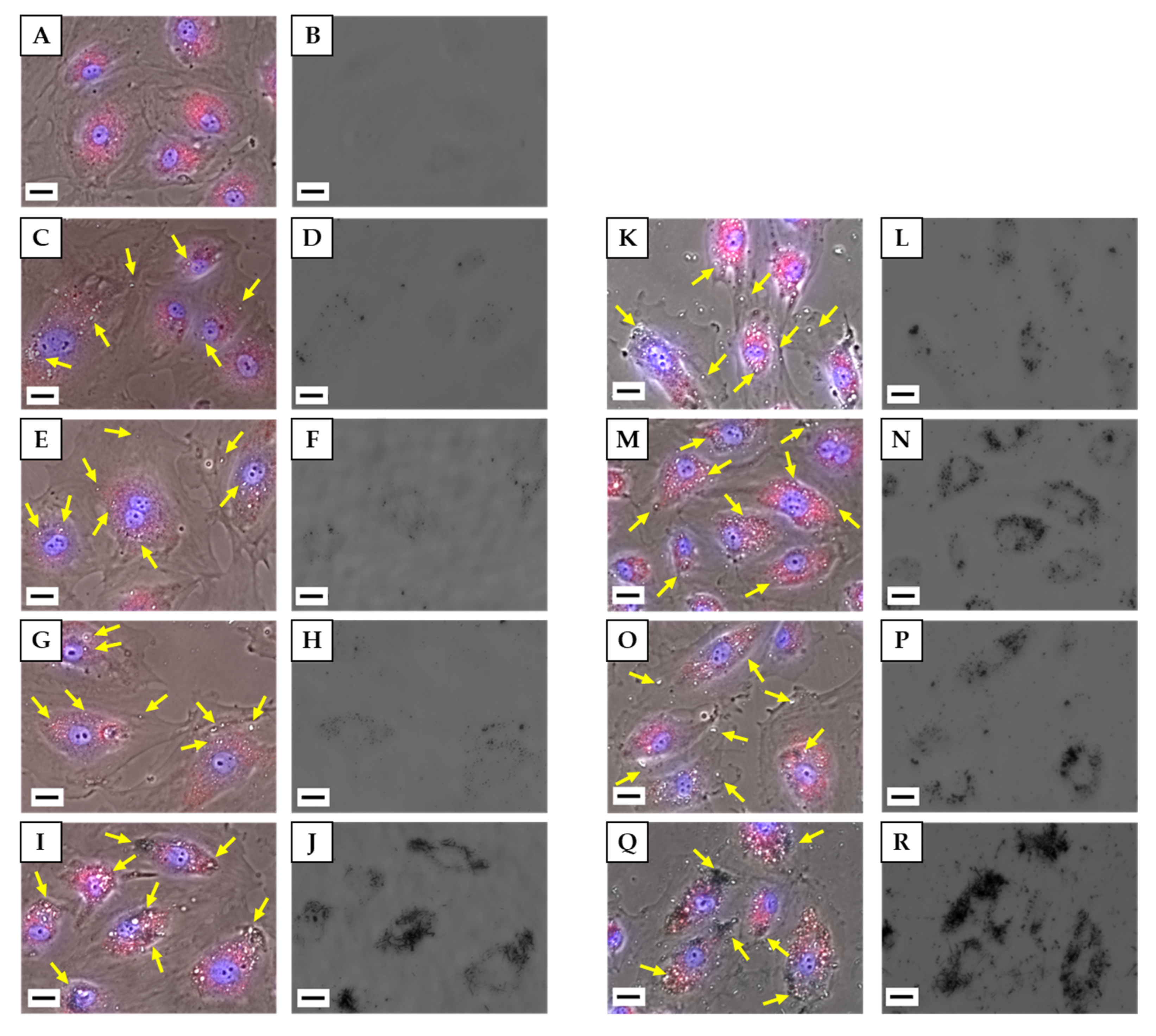
3.4.1. Observation of Cells by Fluorescence Microscopy
3.4.2. Observation of Cells by TEM
3.4.3. Quantification of Intracellular CNMs
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klębowski, B.; Depciuch, J.; Parlińska-Wojtan, M.; Baran, J. Applications of Noble Metal-Based Nanoparticles in Medicine. Int. J. Mol. Sci. 2018, 19, 4031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rostami, I.; Rezvani Alanagh, H.; Hu, Z.; Shahmoradian, S.H. Breakthroughs in medicine and bioimaging with up-conversion nanoparticles. Int. J. Nanomed. 2019, 14, 7759–7780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, M.P.; Sharei, A.; Ding, X.; Sahay, G.; Langer, R.; Jensen, K.F. In vitro and ex vivo strategies for intracellular delivery. Nature 2016, 538, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, N.; Haniu, H.; Usui, Y.; Aoki, K.; Hara, K.; Takanashi, S.; Shimizu, M.; Narita, N.; Okamoto, M.; Kobayashi, S.; et al. Safe Clinical Use of Carbon Nanotubes as Innovative Biomaterials. Chem. Rev. 2014, 114, 6040–6079. [Google Scholar] [CrossRef]
- Chung, S.; Revia, R.A.; Zhang, M. Graphene Quantum Dots and Their Applications in Bioimaging, Biosensing, and Therapy. Adv. Mater. 2019, e1904362. [Google Scholar] [CrossRef]
- Bacakova, L.; Pajorova, J.; Tomkova, M.; Matejka, R.; Broz, A.; Stepanovska, J.; Prazak, S.; Skogberg, A.; Siljander, S.; Kallio, P. Applications of Nanocellulose/Nanocarbon Composites: Focus on Biotechnology and Medicine. Nanomaterials 2020, 10, 196. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Witkowski, C.M.; Craig, M.M.; Greenwade, M.M.; Joseph, K.L. Cytotoxicity Effects of Different Surfactant Molecules Conjugated to Carbon Nanotubes on Human Astrocytoma Cells. Nanoscale Res. Lett. 2009, 4, 1517–1523. [Google Scholar] [CrossRef] [Green Version]
- Madannejad, R.; Shoaie, N.; Jahanpeyma, F.; Darvishi, M.H.; Azimzadeh, M.; Javadi, H. Toxicity of carbon-based nanomaterials: Reviewing recent reports in medical and biological systems. Chem. Biol. Interact 2019, 307, 206–222. [Google Scholar] [CrossRef]
- Fadeel, B.; Bussy, C.; Merino, S.; Vázquez, E.; Flahaut, E.; Mouchet, F.; Evariste, L.; Gauthier, L.; Koivisto, A.J.; Vogel, U.; et al. Safety Assessment of Graphene-Based Materials: Focus on Human Health and the Environment. ACS Nano 2018, 12, 10582–10620. [Google Scholar] [CrossRef]
- Iijima, S.; Yudasaka, M.; Yamada, R.; Bandow, S.; Suenaga, K.; Koka, I.F.; Takahashi, K. Nano-Aggregates of Single-Walled Graphitic Carbon Nano-Horns. Chem. Phys. Lett. 1999, 309, 165–170. [Google Scholar] [CrossRef]
- Utsumi, S.; Miyawaki, J.; Tanaka, H.; Hattori, Y.; Itoi, T.; Ichikuni, N.; Kanoh, H.; Yudasaka, M.; Iijima, S.; Kaneko, K. Opening mechanism of internal nanoporosity of single-wall carbon nanohorn. J. Phys. Chem. B 2005, 109, 14319–14324. [Google Scholar] [CrossRef] [PubMed]
- Ajima, K.; Yudasaka, M.; Suenaga, K.; Kasuya, D.; Azami, T.; Iijima, S. Materials Storage Mechanism in Porous Nanocarbons. Adv. Mater. 2004, 16, 397–401. [Google Scholar] [CrossRef]
- Fan, J.; Yudasaka, M.; Miyawaki, J.; Ajima, K.; Murata, K.; Iijima, S. Control of Hole Opening in Single-Wall Carbon Nanotubes and Single-Wall Carbon Nanohorns Using Oxygen. J. Phys. Chem. B 2006, 110, 1587–1591. [Google Scholar] [CrossRef]
- Ajima, K.; Murakami, T.; Mizoguchi, Y.; Tsuchida, K.; Ichihashi, T.; Iijima, S.; Yudasaka, M. Enhancement of in vivo anticancer effects of cisplatin by incorporation inside single-wall carbon nanohorns. ACS Nano 2008, 2, 2057–2064. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Murakami, T.; Ajima, K.; Tsuchida, K.; Sandanayaka, A.S.; Ito, O.; Iijima, S.; Yudasaka, M. Fabrication of ZnPc/protein nanohorns for double photodynamic and hyperthermic cancer phototherapy. Proc. Natl. Acad. Sci. USA 2008, 105, 14773–14778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Shi, Z.; Li, S.; Zhang, Y.; He, B.; Peng, D.; Tian, J.; Zhao, M.; Wang, X.; Zhang, Q. The interactions of single-wall carbon nanohorns with polar epithelium. Int. J. Nanomed. 2017, 12, 4177–4194. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Yokoyama, A.; Nodasaka, Y.; Kohgo, T.; Motomiya, K.; Matsumoto, H.; Nakazawa, E.; Numata, T.; Zhang, M.; Yudasaka, M.; et al. Long-term biopersistence of tangled oxidized carbon nanotubes inside and outside macrophages in rat subcutaneous tissue. Sci. Rep. 2013, 3, 2516. [Google Scholar] [CrossRef]
- Miyawaki, J.; Yudasaka, M.; Azami, T.; Kubo, Y.; Iijima, S. Toxicity of single-walled carbon nanohorns. ACS Nano 2008, 2, 213–226. [Google Scholar] [CrossRef]
- Shi, Y.; Peng, D.; Wang, D.; Zhao, Z.; Chen, B.; He, B.; Zhu, Y.; Wang, K.; Tian, J.; Zhang, Q. Biodistribution Survey of Oxidized Single-Wall Carbon Nanohorns Following Different Administration Routes by Using Label-Free Multispectral Optoacoustic Tomography. Int. J. Nanomed. 2019, 14, 9809–9821. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Yamaguchi, T.; Iijima, S.; Yudasaka, M. Size-dependent biodistribution of carbon nanohorns in vivo. Nanomedicine 2013, 9, 657–664. [Google Scholar] [CrossRef]
- Miyawaki, J.; Matsumura, S.; Yuge, R.; Murakami, T.; Sato, S.; Tomida, A.; Tsuruo, T.; Ichihashi, T.; Fujinami, T.; Irie, H.; et al. Biodistribution and ultrastructural localization of single-walled carbon nanohorns determined in vivo with embedded Gd2O3 labels. ACS Nano 2009, 3, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, N.R.; Møller, P.; Clausen, P.A.; Saber, A.T.; Micheletti, C.; Jensen, K.A.; Wallin, H.; Vogel, U. Biodistribution of Carbon Nanotubes in Animal Models. Basic Clin. Pharmacol. Toxicol. 2017, 121 (Suppl. 3), 30–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinohara, N.; Nakazato, T.; Ohkawa, K.; Tamura, M.; Kobayashi, N.; Morimoto, Y.; Oyabu, T.; Myojo, T.; Shimada, M.; Yamamoto, K.; et al. Long-term retention of pristine multi-walled carbon nanotubes in rat lungs after intratracheal instillation. J. Appl. Toxicol. 2016, 36, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Monteiro-Riviere, N.A.; Riviere, J.E. Pharmacokinetics of metallic nanoparticles. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2015, 7, 189–217. [Google Scholar] [CrossRef]
- Hara, K.; Aoki, K.; Usui, Y.; Shimizu, M.; Narita, N.; Ogihara, N.; Nakamura, K.; Ishigaki, N.; Sano, K.; Haniu, H.; et al. Evaluation of CNT toxicity in comparison to tattoo ink nanoparticles for use as a biomaterial. Mater. Today 2011, 14, 434–440. [Google Scholar] [CrossRef]
- Werengowska-Ciećwierz, K.; Wiśniewski, M.; Terzyk, A.P.; Roszek, K.; Czarnecka, J.; Bolibok, P.; Rychlicki, G. Conscious Changes of Carbon Nanotubes Cytotoxicity by Manipulation with Selected Nanofactors. Appl. Biochem. Biotechnol. 2015, 176, 730–741. [Google Scholar] [CrossRef] [Green Version]
- Czarnecka, J.; Wiśniewski, M.; Forbot, N.; Bolibok, P.; Terzyk, A.P.; Roszek, K. Cytotoxic or Not? Disclosing the Toxic Nature of Carbonaceous Nanomaterials through Nano-Bio Interactions. Materials 2020, 13, 2060. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhou, X.; Iijima, S.; Yudasaka, M. Small-sized carbon nanohorns enabling cellular uptake control. Small 2012, 8, 2524–2531. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, C.; Haniu, H.; Ajima, K.; Tanaka, M.; Sobajima, A.; Ishida, H.; Tsukahara, T.; Matsuda, Y.; Aoki, K.; Kato, H.; et al. The Dispersion State of Tangled Multi-Walled Carbon Nanotubes Affects Their Cytotoxicity. Nanomaterials 2016, 6, 219. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, C.; Ueda, K.; Haniu, H.; Ishida, H.; Okano, S.; Takizawa, T.; Sobajima, A.; Kamanaka, T.; Yoshida, K.; Okamoto, M.; et al. Different aggregation and shape characteristics of carbon materials affect biological responses in RAW264 cells. Int. J. Nanomed. 2018, 13, 6079–6088. [Google Scholar] [CrossRef] [Green Version]
- Caputo, F.; De Nicola, M.; Ghibelli, L. Pharmacological potential of bioactive engineered nanomaterials. Biochem. Pharmacol. 2014, 92, 112–130. [Google Scholar] [CrossRef]
- Serup, J.; Carlsen, K.H.; Sepehri, M. Tattoo complaints and complications: Diagnosis and clinical spectrum. In Tattooed Skin and Health; Karger Publishers: Basel, Switzerland, 2015; Volume 48, pp. 48–60. [Google Scholar]
- Kluger, N.; Koljonen, V. Tattoos, inks, and cancer. Lancet Oncol. 2012, 13, e161–e168. [Google Scholar] [CrossRef]
- Haniu, H.; Saito, N.; Matsuda, Y.; Kim, Y.A.; Park, K.C.; Tsukahara, T.; Usui, Y.; Aoki, K.; Shimizu, M.; Ogihara, N.; et al. Elucidation mechanism of different biological responses to multi-walled carbon nanotubes using four cell lines. Int. J. Nanomed. 2011, 6, 3487–3497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Zhang, X.; Sun, L.; Wei, Y.; Wei, X. Cellular Toxicity and Immunological Effects of Carbon-based Nanomaterials. Part Fibre Toxicol. 2019, 16, 18. [Google Scholar] [CrossRef] [PubMed]
- Flahaut, E.; Durrieu, M.C.; Remy-Zolghadri, M.; Bareille, R.; Baquey, C. Investigation of the cytotoxicity of CCVD carbon nanotubes towards human umbilical vein endothelial cells. Carbon 2006, 44, 1093–1099. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Chang, S.; Long, J.; Li, J.; Li, X.; Cao, Y. The toxicity of multi-walled carbon nanotubes (MWCNTs) to human endothelial cells: The influence of diameters of MWCNTs. Food Chem. Toxicol. 2019, 126, 169–177. [Google Scholar] [CrossRef]
- Yamawaki, H.; Iwai, N. Cytotoxicity of water-soluble fullerene in vascular endothelial cells. Am. J. Physiol. Cell Physiol. 2006, 290, C1495–C1502. [Google Scholar] [CrossRef] [Green Version]
- Yamawaki, H.; Iwai, N. Mechanisms underlying nano-sized air-pollution-mediated progression of atherosclerosis: Carbon black causes cytotoxic injury/inflammation and inhibits cell growth in vascular endothelial cells. Circ. J. 2006, 70, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Tammela, T.; Alitalo, K. Lymphangiogenesis: Molecular mechanisms and future promise. Cell 2010, 140, 460–476. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Cheng, Y. Gene expression profile of lymphatic endothelial cells. Cell Biol. Int. 2011, 35, 1177–1187. [Google Scholar] [CrossRef]
- Niemelä, H.; Elima, K.; Henttinen, T.; Irjala, H.; Salmi, M.; Jalkanen, S. Molecular identification of PAL-E, a widely used endothelial-cell marker. Blood 2005, 106, 3405–3409. [Google Scholar] [CrossRef] [Green Version]
- Stan, R.V.; Tkachenko, E.; Niesman, I.R. PV1 is a key structural component for the formation of the stomatal and fenestral diaphragms. Mol. Biol. Cell 2004, 15, 3615–3630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosma, E.K.; van Noorden, C.J.F.; Schlingemann, R.O.; Klaassen, I. The role of plasmalemma vesicle-associated protein in pathological breakdown of blood-brain and blood-retinal barriers: Potential novel therapeutic target for cerebral edema and diabetic macular edema. Fluids Barriers CNS 2018, 15, 24. [Google Scholar] [CrossRef] [Green Version]
- Pelkmans, L.; Kartenbeck, J.; Helenius, A. Caveolar endocytosis of simian virus 40 reveals a new two-step vesicular-transport pathway to the ER. Nat. Cell Biol. 2001, 3, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Nichols, B.J. A distinct class of endosome mediates clathrin-independent endocytosis to the Golgi complex. Nat. Cell Biol. 2002, 4, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Wolff, B.; Burns, A.R.; Middleton, J.; Rot, A. Endothelial cell “memory” of inflammatory stimulation: Human venular endothelial cells store interleukin 8 in Weibel-Palade bodies. J. Exp. Med. 1998, 188, 1757–1762. [Google Scholar] [CrossRef] [Green Version]
- Van Mourik, J.A.; Romani de Wit, T.; Voorberg, J. Biogenesis and exocytosis of Weibel-Palade bodies. Histochem. Cell Biol. 2002, 117, 113–122. [Google Scholar] [CrossRef]
- Kawecki, C.; Lenting, P.J.; Denis, C.V. von Willebrand factor and inflammation. J. Thromb. Haemost. 2017, 15, 1285–1294. [Google Scholar] [CrossRef]
- Maruyama, K.; Haniu, H.; Saito, N.; Matsuda, Y.; Tsukahara, T.; Kobayashi, S.; Tanaka, M.; Aoki, K.; Takanashi, S.; Okamoto, M.; et al. Endocytosis of Multiwalled Carbon Nanotubes in Bronchial Epithelial and Mesothelial Cells. Biomed Res. Int. 2015, 2015, 793186. [Google Scholar] [CrossRef]
- Liu, N.; Tang, M. Toxic effects and involved molecular pathways of nanoparticles on cells and subcellular organelles. J. Appl. Toxicol. 2020, 40, 16–36. [Google Scholar] [CrossRef]
- Alnasser, F.; Castagnola, V.; Boselli, L.; Esquivel-Gaon, M.; Efeoglu, E.; McIntyre, J.; Byrne, H.J.; Dawson, K.A. Graphene Nanoflake Uptake Mediated by Scavenger Receptors. Nano Lett. 2019, 19, 1260–1268. [Google Scholar] [CrossRef] [PubMed]
- Hirano, S.; Fujitani, Y.; Furuyama, A.; Kanno, S. Macrophage receptor with collagenous structure (MARCO) is a dynamic adhesive molecule that enhances uptake of carbon nanotubes by CHO-K1 cells. Toxicol. Appl. Pharmacol. 2012, 259, 96–103. [Google Scholar] [CrossRef] [PubMed]
- ENSG00000019169-MARCO and ENSG00000073060-SCARB1. Available online: https://www.proteinatlas.org/ (accessed on 10 July 2020).
- Shannahan, J.H.; Brown, J.M.; Chen, R.; Ke, P.C.; Lai, X.; Mitra, S.; Witzmann, F.A. Comparison of nanotube-protein corona composition in cell culture media. Small 2013, 9, 2171–2181. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, M.; Saeedi-Eslami, S.N.; Shokrgozar, M.A.; Azadmanesh, K.; Hassanlou, M.; Kalhor, H.R.; Burtea, C.; Rothen-Rutishauser, B.; Laurent, S.; Sheibani, S.; et al. Cell “vision”: Complementary factor of protein corona in nanotoxicology. Nanoscale 2012, 4, 5461–5468. [Google Scholar] [CrossRef] [PubMed]
- Holder, A.L.; Carter, B.J.; Goth–Goldstein, R.; Lucas, D.; Koshland, C.P. Increased cytotoxicity of oxidized flame soot. Atmos Pollut. Res. 2012, 3, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Eldridge, B.N.; Xing, F.; Fahrenholtz, C.D.; Singh, R.N. Evaluation of multiwalled carbon nanotube cytotoxicity in cultures of human brain microvascular endothelial cells grown on plastic or basement membrane. Toxicol. In Vitro 2017, 41, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Haniu, H.; Saito, N.; Matsuda, Y.; Kim, Y.A.; Park, K.C.; Tsukahara, T.; Usui, Y.; Aoki, K.; Shimizu, M.; Ogihara, N.; et al. Effect of dispersants of multi-walled carbon nanotubes on cellular uptake and biological responses. Int. J. Nanomed. 2011, 6, 3295–3307. [Google Scholar] [CrossRef] [Green Version]
- Takeda, A.; Hollmén, M.; Dermadi, D.; Pan, J.; Brulois, K.F.; Kaukonen, R.; Lönnberg, T.; Boström, P.; Koskivuo, I.; Irjala, H.; et al. Single-cell survey of human lymphatics unveils marked endothelial cell heterogeneity and mechanisms of homing for neutrophils. Immunity 2019, 51, 561–572. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CNMs | Average Particle Size (nm) |
|---|---|
| unCNH | 155.5 ± 61.6 |
| oxCNH | 150.7 ± 55.6 |
| CB | 222.2 ± 111.8 |
| MWCNT | 677.5 ± 697.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sano, M.; Izumiya, M.; Haniu, H.; Ueda, K.; Konishi, K.; Ishida, H.; Kuroda, C.; Uemura, T.; Aoki, K.; Matsuda, Y.; et al. Cellular Responses of Human Lymphatic Endothelial Cells to Carbon Nanomaterials. Nanomaterials 2020, 10, 1374. https://doi.org/10.3390/nano10071374
Sano M, Izumiya M, Haniu H, Ueda K, Konishi K, Ishida H, Kuroda C, Uemura T, Aoki K, Matsuda Y, et al. Cellular Responses of Human Lymphatic Endothelial Cells to Carbon Nanomaterials. Nanomaterials. 2020; 10(7):1374. https://doi.org/10.3390/nano10071374
Chicago/Turabian StyleSano, Mahoko, Makoto Izumiya, Hisao Haniu, Katsuya Ueda, Kosuke Konishi, Haruka Ishida, Chika Kuroda, Takeshi Uemura, Kaoru Aoki, Yoshikazu Matsuda, and et al. 2020. "Cellular Responses of Human Lymphatic Endothelial Cells to Carbon Nanomaterials" Nanomaterials 10, no. 7: 1374. https://doi.org/10.3390/nano10071374
APA StyleSano, M., Izumiya, M., Haniu, H., Ueda, K., Konishi, K., Ishida, H., Kuroda, C., Uemura, T., Aoki, K., Matsuda, Y., & Saito, N. (2020). Cellular Responses of Human Lymphatic Endothelial Cells to Carbon Nanomaterials. Nanomaterials, 10(7), 1374. https://doi.org/10.3390/nano10071374