
Predicting Finite-Bias Tunneling Current Properties from Zero-Bias Features: The Frontier Orbital Bias Dependence at an Exemplar Case of DNA Nucleotides in a Nanogap

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Computational Details
3. Results
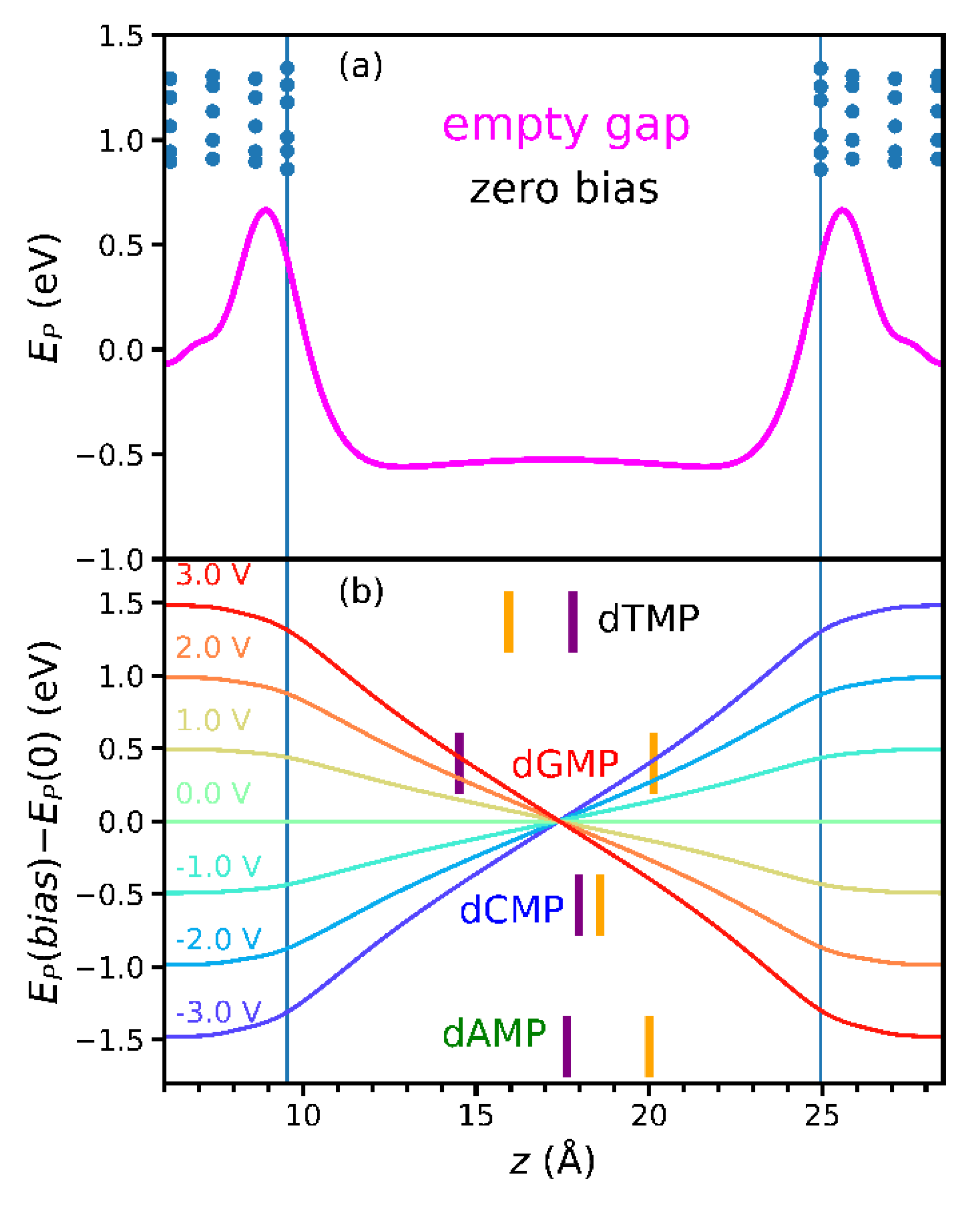
3.1. Empty Gap Properties and the Weak Pinning
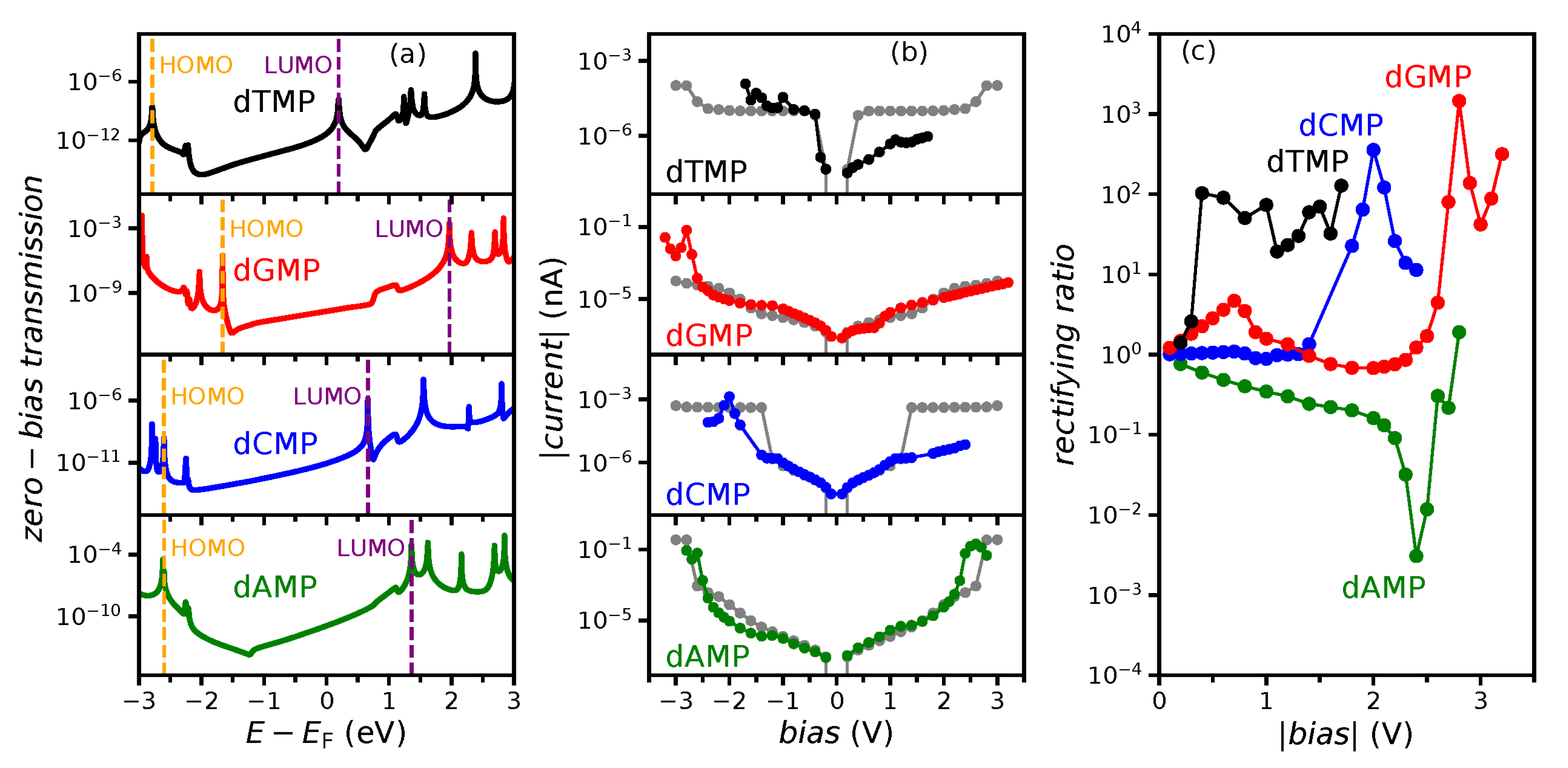
3.2. Finite-Bias Properties of the Nucleotide Molecular Junction
3.3. Estimation of FiniteBias from the Empty Gap and Zero-Bias Properties
3.4. The Tunneling Current and Rectification
3.5. Zero-Bias Approximation
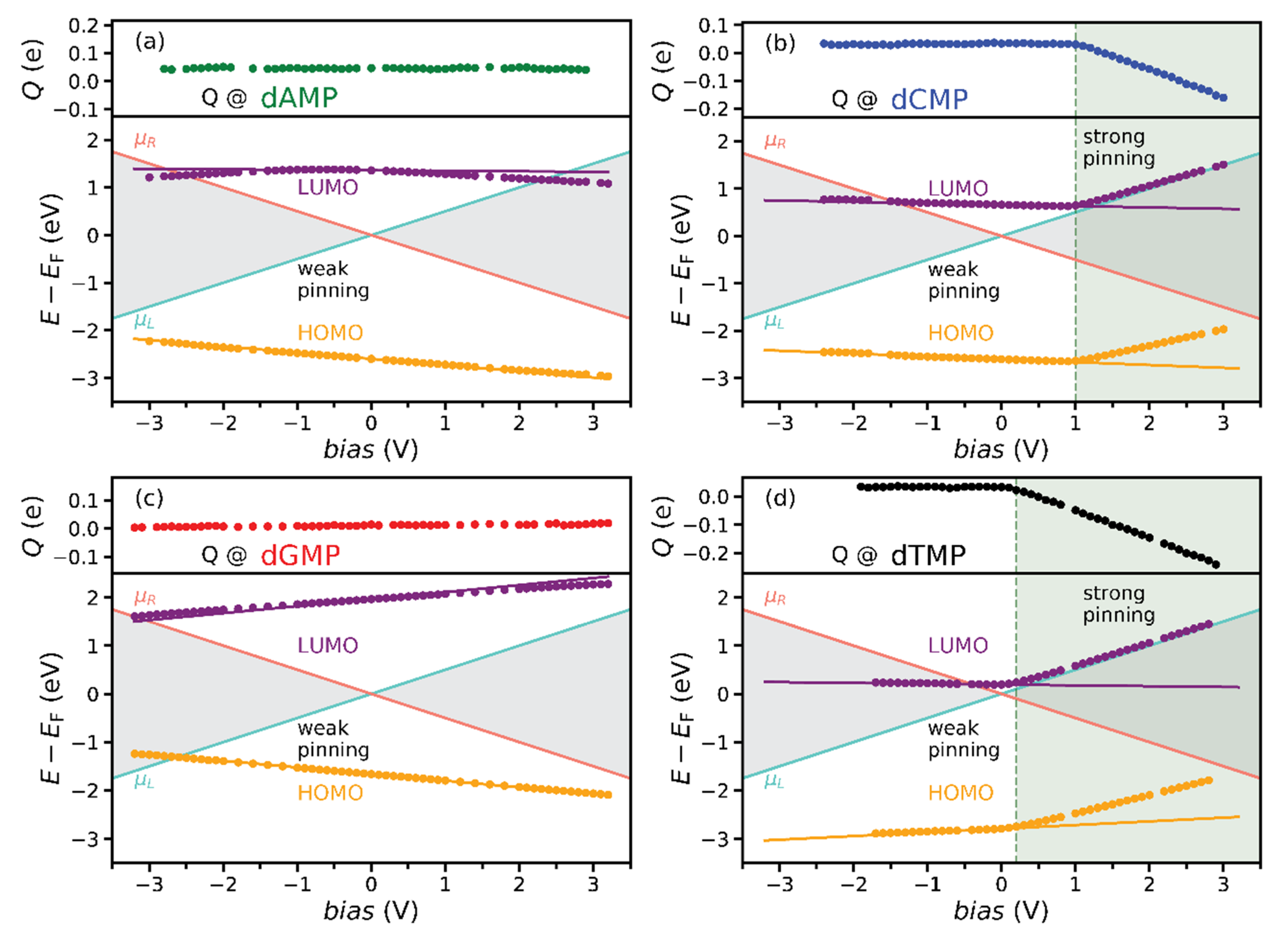
3.6. Rules for Pinning Regimes Diagram Construction
- From zero-bias transport calculation, obtain HOMO and LUMO: their energies EHOMO/LUMO, wave functions ψHOMO/LUMO, and average z coordinate <zHOMO/LUMO> of atoms having the most significant contribution to the orbital (whose atomic orbital coefficients c from Equation (1) are higher than 0.1, for example);
- Calculate the bias dependence of the empty-gap electrostatic potential EP(bias) − EP(0) at <zHOMO/LUMO> (in fact, this step does not even require finite-bias transport calculation);
- Calculate HOMO/LUMO weak-pinning energy EWPHOMO/LUMO(bias) from Equation (3) and voltage at which it becomes equal to μL and μR. At these voltages, the orbital energy enters either the bias window (a possible drastic increase in the tunneling current) or the SP regime (no dramatic tunneling current changes;
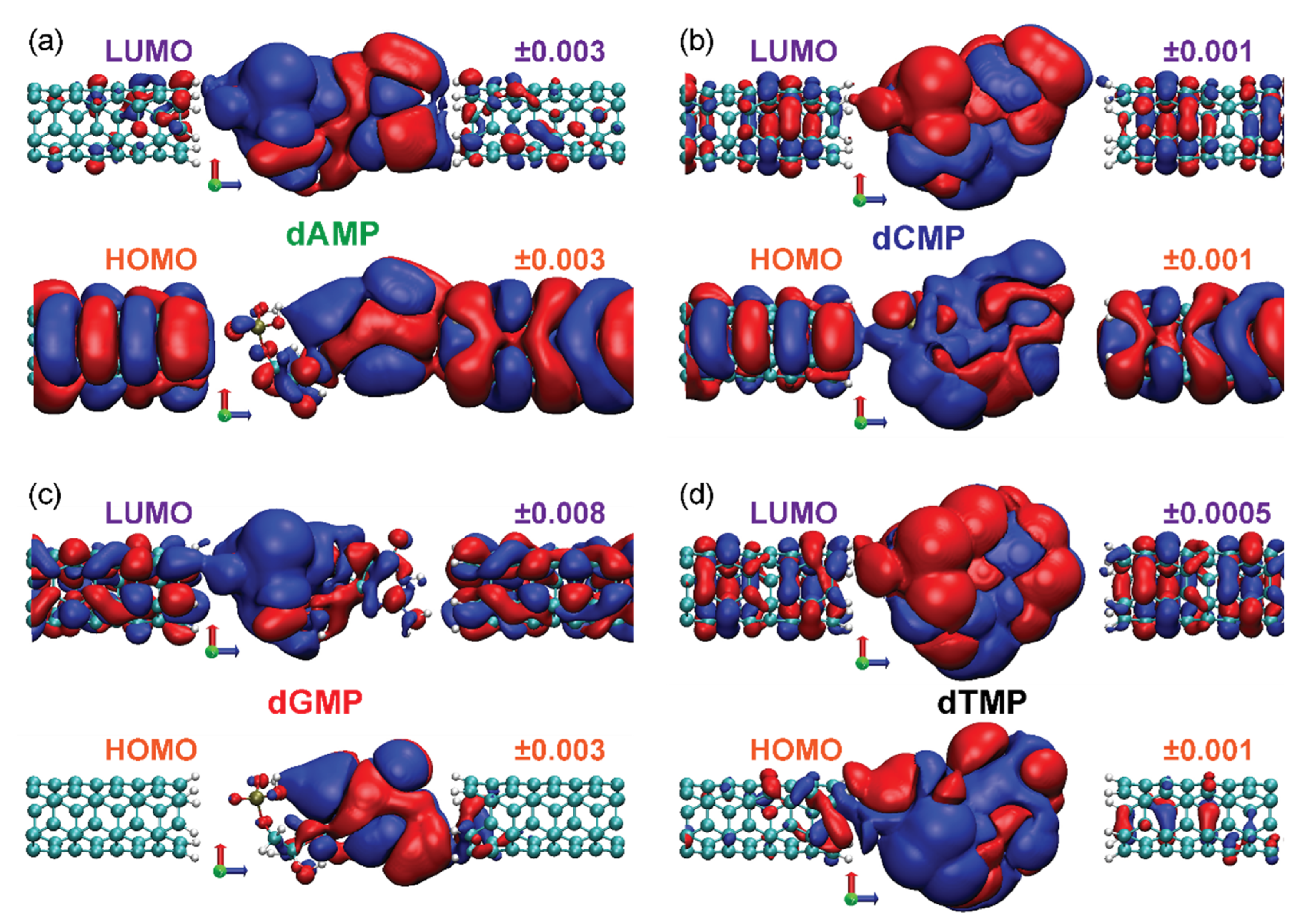
- Check the spatial overlap of the orbital with both electrodes (ψHOMO/LUMO). The strong pinning will occur if there exists a dominant overlap (hybridization) of the orbital with the electrode whose electrochemical potential EWPHOMO/LUMO intersects.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Branton, D.; Deamer, D.W.; Marziali, A.; Bayley, H.; Benner, S.A.; Butler, T.; Ventra, M.D.; Garaj, S.; Hibbs, A.; Huang, X.; et al. The potential and challenges of nanopore sequencing. Nat. Biotechnol. 2008, 26, 1146. [Google Scholar] [CrossRef]
- Di Ventra, M.; Taniguchi, M. Decoding DNA, RNA and peptides with quantum tunneling. Nat. Nanotechnol. 2016, 11, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Heerema, S.J.; Dekker, C. Graphene nanodevices for DNA sequencing. Nature Nanotechnol. 2016, 11, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Restrepo-Pérez, L.; Joo, C.; Dekker, C. Paving the way to single-molecule protein sequencing. Nat. Nanotechnol. 2018, 13, 786–796. [Google Scholar] [CrossRef] [Green Version]
- Yokota, K.; Tsutsui, M.; Taniguchi, M. Electrode-embedded nanopores for label-free single-molecule sequencing by electric currents. RSC Adv. 2014, 4, 15886. [Google Scholar] [CrossRef]
- Fanget, A.; Traversi, F.; Khlybov, S.; Granjon, P.; Magrez, A.; Forró, L.; Radenovic, A. Nanopore integrated nanogaps for DNA detection. Nano Lett. 2014, 14, 244–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Mihovilovic, M.; Chan, J.; Stein, D. Fabrication of nanopores with embedded annular electrodes and transverse carbon nanotube electrodes. J. Phys.-Condens. Matter 2010, 22, 454114. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Rungger, I.; Pemmaraju, C.D.; Schwingenschlögl, U.; Sanvito, S. First-principles study of high-conductance DNA sequencing with carbon nanotube electrodes. Phys. Rev. B 2012, 85, 115436. [Google Scholar] [CrossRef] [Green Version]
- Prasongkit, J.; Grigoriev, A.; Pathak, B.; Ahuja, R.; Scheicher, R.H. Transverse conductance of DNA nucleotides in a graphene nanogap from first principles. Nano Lett. 2011, 11, 1941–1945. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, J.N.; Boynton, P.; Di Ventra, M.; Jauho, A.-P.; Flyvbjerg, H. Classification of DNA nucleotides with transverse tunneling currents. Nanotechnology 2016, 28, 015502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graf, M.; Lihter, M.; Altus, D.; Marion, S.; Radenovic, A. Transverse detection of DNA using a MoS2 nanopore. Nano Lett. 2019, 19, 9075–9083. [Google Scholar] [CrossRef]
- Tsutsui, M.; Taniguchi, M.; Yokota, K.; Kawai, T. Identifying single nucleotides by tunnelling current. Nat. Nanotechnol. 2010, 5, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Zikic, R.; Krstić, P.S.; Zhang, X.-G.; Fuentes-Cabrera, M.; Wells, J.; Zhao, X. Characterization of the tunneling conductance across DNA bases. Phys. Rev. E 2006, 74, 011919. [Google Scholar] [CrossRef] [PubMed]
- Amorim, R.G.; Rocha, A.R.; Scheicher, R.H. Boosting DNA recognition sensitivity of graphene nanogaps through nitrogen edge functionalization. J. Phys. Chem. C 2016, 120, 19384–19388. [Google Scholar] [CrossRef]
- Bellunato, A.; Vrbica, S.D.; Sabater, C.; de Vos, E.W.; Fermin, R.; Kanneworff, K.N.; Galli, F.; van Ruitenbeek, J.M.; Schneider, G.F. Dynamic tunneling junctions at the atomic intersection of two twisted graphene edges. Nano Lett. 2018, 18, 2505–2510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhuri, P.; Frota, H.O.; Mota, C.; Ghosh, A. Conductance through glycine in a graphene nanogap. J. Nanopart. Res. 2018, 20, 158. [Google Scholar] [CrossRef] [Green Version]
- Meunier, V.; Krstić, P.S. Enhancement of the transverse conductance in DNA nucleotides. J. Chem. Phys. 2008, 128, 041103. [Google Scholar] [CrossRef] [PubMed]
- Djurišić, I.; Dražić, M.S.; Tomović, A.Ž.; Spasenović, M.; Šljivančanin, Ž.; Jovanović, V.P.; Zikic, R. DNA sequencing with single-stranded DNA rectification in a nanogap gated by N-terminated carbon nanotube electrodes. ACS Appl. Nano Mater. 2020, 3, 3034–3043. [Google Scholar] [CrossRef]
- Shukla, V.; Jena, N.K.; Grigoriev, A.; Ahuja, R. Prospects of graphene−hBN heterostructure nanogap for DNA sequencing. ACS Appl. Mater. Interfaces 2017, 9, 39945–39952. [Google Scholar] [CrossRef]
- Prasongkit, J.; Grigoriev, A.; Pathak, B.; Ahuja, R.; Scheicher, R.H. Theoretical study of electronic transport through DNA nucleotides in a double-functionalized graphene nanogap. J. Phys. Chem. C 2013, 117, 15421–15428. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.; Sen, S.; Im, J.O.; Biswas, S.; Krstic, P.; Ashcroft, B.; Borges, C.; Zhao, Y.; Lindsay, S.; Zhang, P. Universal readers based on hydrogen bonding or π−π stacking for identification of DNA nucleotides in electron tunnel junctions. ACS Nano 2016, 10, 11304–11316. [Google Scholar] [CrossRef] [PubMed]
- Datta, S. Electronic Transport in Mesoscopic Systems; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Ding, W.; Koepf, M.; Koenigsmann, C.; Batra, A.; Venkataraman, L.; Negre, C.F.A.; Brudvig, G.W.; Crabtree, R.H.; Schmuttenmaer, C.A.; Batista, V.S. Computational design of intrinsic molecular rectifiers based on asymmetric functionalization of N-phenylbenzamide. J. Chem. Theory Comput. 2015, 11, 5888–5896. [Google Scholar] [CrossRef] [PubMed]
- Van Dyck, C.; Ratner, M.A. Molecular junctions: Control of the energy gap achieved by a pinning effect. J. Phys. Chem. C 2017, 121, 3013–3024. [Google Scholar] [CrossRef]
- Xin, N.; Guan, J.; Zhou, C.; Chen, X.; Gu, C.; Li, Y.; Ratner, M.A.; Nitzan, A.; Stoddart, J.F.; Guo, X. Concepts in the design and engineering of single-molecule electronic devices. Nat. Rev. Phys. 2019, 1, 211–230. [Google Scholar] [CrossRef]
- Capozzi, B.; Xia, J.; Adak, O.; Dell, E.J.; Liu, Z.-F.; Taylor, J.C.; Neaton, J.B.; Campos, L.M.; Venkataraman, L. Single-molecule diodes with high rectification ratios through environmental control. Nat. Nanotechnol. 2015, 10, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Adak, O.; Korytaár, R.; Joe, A.Y.; Evers, F.; Venkataraman, L. Impact of electrode density of states on transport through pyridine-linked single molecule junctions. Nano Lett. 2015, 15, 3716–3722. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Roemer, M.; Yuan, L.; Du, W.; Thompson, D.; del Barco, E.; Nijhuis, C.A. Molecular diodes with rectification ratios exceeding 105 driven by electrostatic interactions. Nat. Nanotechnol. 2017, 12, 797–803. [Google Scholar] [CrossRef]
- Djurišić, I.; Dražić, M.S.; Tomović, A.Ž.; Jovanović, V.P.; Zikic, R. Electrostatically driven energy shift of molecular orbitals of benzene and nicotine in carbon nanotube gaps. J. Nanopart. Res. 2021, 23, 15. [Google Scholar] [CrossRef]
- Van Dyck, C.; Ratner, M.A. Molecular rectifiers: A new design based on asymmetric anchoring moieties. Nano Lett. 2015, 15, 1577–1584. [Google Scholar] [CrossRef]
- Thong, A.Z.; Shaffer, M.S.P.; Horsfield, A.P. HOMO–LUMO coupling: The fourth rule for highly effective molecular rectifiers. Nanoscale 2017, 9, 8119–8125. [Google Scholar] [CrossRef] [Green Version]
- Soler, J.M.; Artacho, E.; Gale, J.D.; García, A.; Junquera, J.; Ordejón, P.; Sánchez-Portal, D. The SIESTA method for ab initio order-N materials simulation. J. Phys.-Condens. Matter 2002, 14, 2745–2779. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.; Brandbyge, M.; Stokbro, K. Theory of rectification in Tour wires: The role of electrode coupling. Phys. Rev. Lett. 2002, 89, 138301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machón, M.; Reich, S.; Thomsen, C.; Sánchez-Portal, D.; Ordejón, P. Ab initio calculations of the optical properties of 4-Å-diameter single-walled nanotubes. Phys. Rev. B 2002, 66, 155410. [Google Scholar] [CrossRef] [Green Version]
- Perdew, J.P.; Burke, K.; Ernzerhof, M. Generalized gradient approximation made simple. Phys. Rev. Lett. 1996, 77, 3865–3868. [Google Scholar] [CrossRef] [Green Version]
- Troullier, N.; Martins, J.L. Efficient pseudopotentials for plane-wave calculations. Phys. Rev. B 1991, 43, 1993–2006. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD—Visual Molecular Dynamics. J. Molec. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Papior, N. Sisl, v0.11.0. 2021. Available online: https://zenodo.org/record/4544334/export/json (accessed on 17 February 2021).
- Van Dyck, C.; Geskin, V.; Kronemeijer, A.J.; de Leeuw, D.M.; Cornil, J. Impact of derivatization on electron transmission through dithienylethene-based photoswitches in molecular junctions. Phys. Chem. Chem. Phys. 2013, 15, 4392. [Google Scholar] [CrossRef] [Green Version]
- Djurišić, I.; Dražić, M.S.; Tomović, A.Ž.; Spasenović, M.; Šljivančanin, Ž.; Jovanović, V.P.; Zikic, R. Field effect and local gating in nitrogen-terminated nanopores (NtNP) and nanogaps (NtNG) in graphene. ChemPhysChem 2021, 22, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Rungger, I.; Sanvito, S.; Inoue, N.; Tominaga, J.; Asai, Y. Resistive switching mechanism of GeTe–Sb2Te3 interfacial phase change memory and topological properties of embedded two-dimensional states. Nanoscale 2017, 9, 9386. [Google Scholar] [CrossRef]
- Inoue, N.; Nakamura, H. Structural transition pathway and bipolar switching of the GeTe-Sb2Te3 superlattice as interfacial phase change memory. Faraday Discuss. 2019, 213, 303–319. [Google Scholar] [CrossRef]
- Seeman, N.; Sleiman, H. DNA nanotechnology. Nat. Rev. Mater. 2018, 3, 17068. [Google Scholar] [CrossRef]
- Wei, S.; Haojie, Y.; Gensheng, W.; Chang, C.; Meng, Y. Manipulation of DNA transport through solid-state nanopores by atomic force microscopy. Mater. Res. Express 2020, 7, 095404. [Google Scholar]
- Pawlak, R.; Vilhena, J.G.; Hinaut, A.; Meier, T.; Glatzel, T.; Baratoff, A.; Gnecco, E.; Pérez, R.; Meyer, E. Conformations and cryo-force spectroscopy of spray-deposited single-strand DNA on gold. Nat. Commun. 2019, 10, 685. [Google Scholar] [CrossRef] [Green Version]
- Romero-Muñiz, C.; Ortega, M.; Vilhena, J.G.; Díez-Pérez, I.; Pérez, R.; Cuevas, J.C.; Zotti, L.A. Can electron transport through a blue-copper azurin be coherent? An ab initio study. J. Phys. Chem. C 2021, 125, 1693–1702. [Google Scholar] [CrossRef]
- Zhuravel, R.; Huang, H.; Polycarpou, G.; Polydorides, S.; Motamarri, P.; Katrivas, L.; Rotem, D.; Sperling, J.; Zotti, L.A.; Kotlyar, A.B.; et al. Backbone charge transport in double-stranded DNA. Nat. Nanotechnol. 2020, 15, 836–840. [Google Scholar] [CrossRef]
- Metzger, R.M. Unimolecular electronics. Chem. Rev. 2015, 115, 5056–5115. [Google Scholar] [CrossRef]
- Berdiyorov, G.R.; Hamoudi, H. Effect of anchoring groups on the electronic transport properties of biphenyl and phenylpyridine molecules. J. Mater. Res. Technol. 2021, 12, 193–201. [Google Scholar] [CrossRef]
- Na, Z.; Wai-Yip, L.; Zhengxu, C.; Lianwei, L.; Luping, Y. Molecular Rectification Tuned by Through-Space Gating Effect. Nano Lett. 2017, 17, 308–312. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djurišić, I.; Jovanović, V.P.; Dražić, M.S.; Tomović, A.Ž.; Zikic, R. Predicting Finite-Bias Tunneling Current Properties from Zero-Bias Features: The Frontier Orbital Bias Dependence at an Exemplar Case of DNA Nucleotides in a Nanogap. Nanomaterials 2021, 11, 3021. https://doi.org/10.3390/nano11113021
Djurišić I, Jovanović VP, Dražić MS, Tomović AŽ, Zikic R. Predicting Finite-Bias Tunneling Current Properties from Zero-Bias Features: The Frontier Orbital Bias Dependence at an Exemplar Case of DNA Nucleotides in a Nanogap. Nanomaterials. 2021; 11(11):3021. https://doi.org/10.3390/nano11113021
Chicago/Turabian StyleDjurišić, Ivana, Vladimir P. Jovanović, Miloš S. Dražić, Aleksandar Ž. Tomović, and Radomir Zikic. 2021. "Predicting Finite-Bias Tunneling Current Properties from Zero-Bias Features: The Frontier Orbital Bias Dependence at an Exemplar Case of DNA Nucleotides in a Nanogap" Nanomaterials 11, no. 11: 3021. https://doi.org/10.3390/nano11113021
APA StyleDjurišić, I., Jovanović, V. P., Dražić, M. S., Tomović, A. Ž., & Zikic, R. (2021). Predicting Finite-Bias Tunneling Current Properties from Zero-Bias Features: The Frontier Orbital Bias Dependence at an Exemplar Case of DNA Nucleotides in a Nanogap. Nanomaterials, 11(11), 3021. https://doi.org/10.3390/nano11113021