
Collagen Film Activation with Nanoscale IKVAV-Capped Dendrimers for Selective Neural Cell Response


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Collagen Film Preparation
2.3. Dendrimer Fluorescence Tag Conjugation
2.4. Incorporation of Dendrimers into 2D Collagen Films
2.5. Dissociation of Dendrimers from 2D Collagen Films
2.6. Cell Attachment Assay
2.7. Cell Proliferation Assay
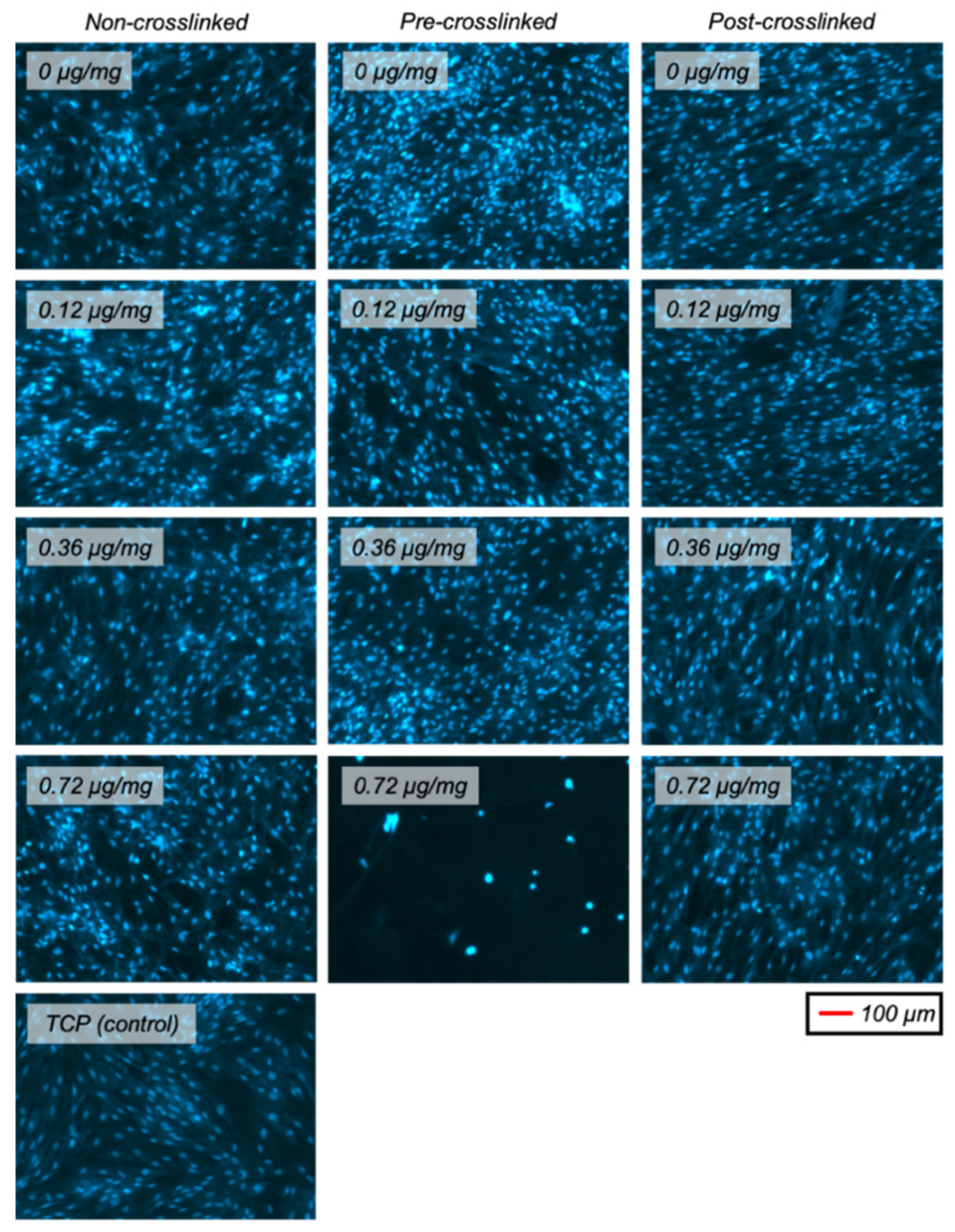
2.8. Fixed Cell Imaging with Fluorescence Microscopy
2.9. Statistical Analysis
3. Results
3.1. Dye-Tagging Dendrimers for Visualisation on Collagen Substrates
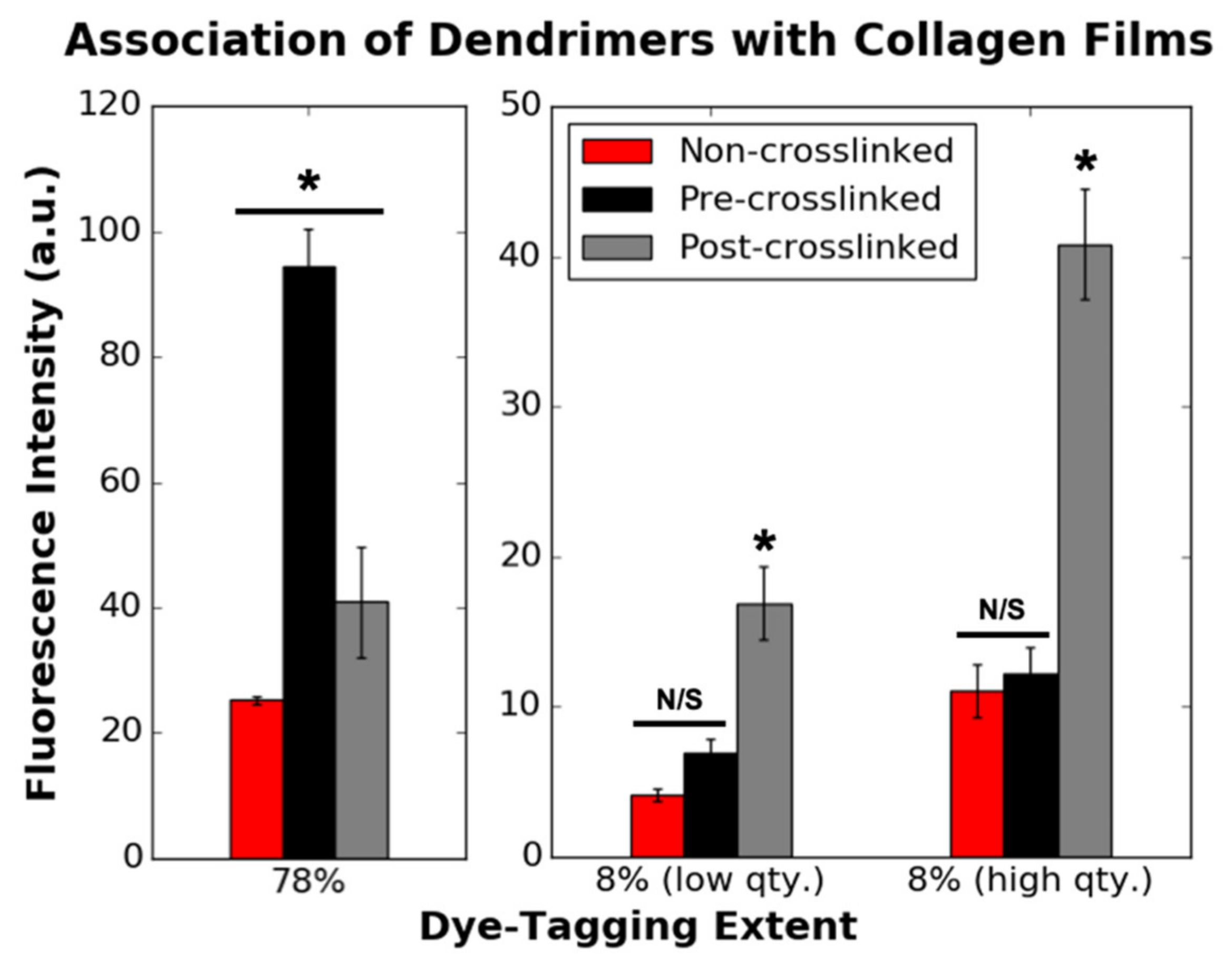
3.2. Association of Dye-Tagged Dendrimers with 2D Collagen Films
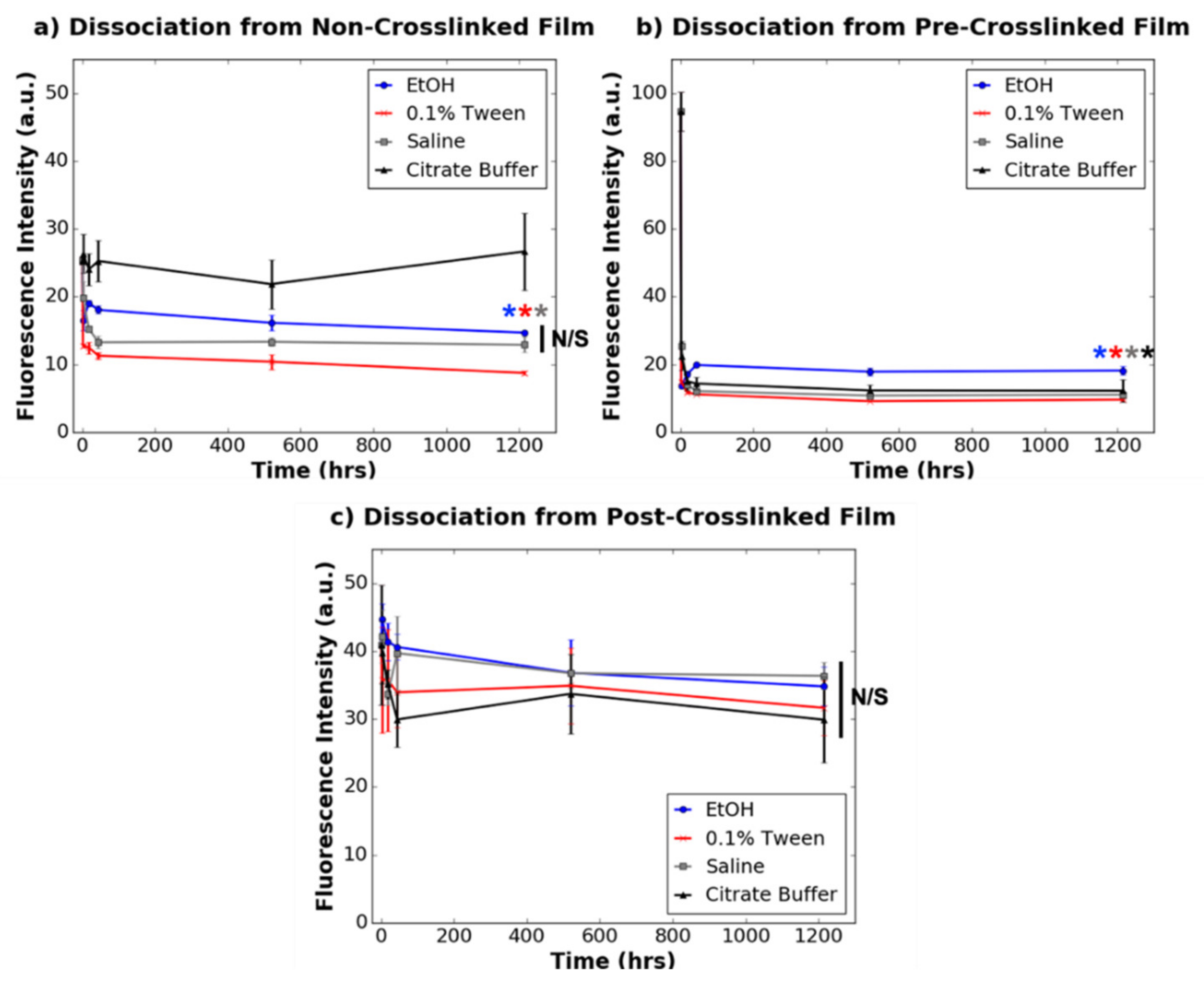
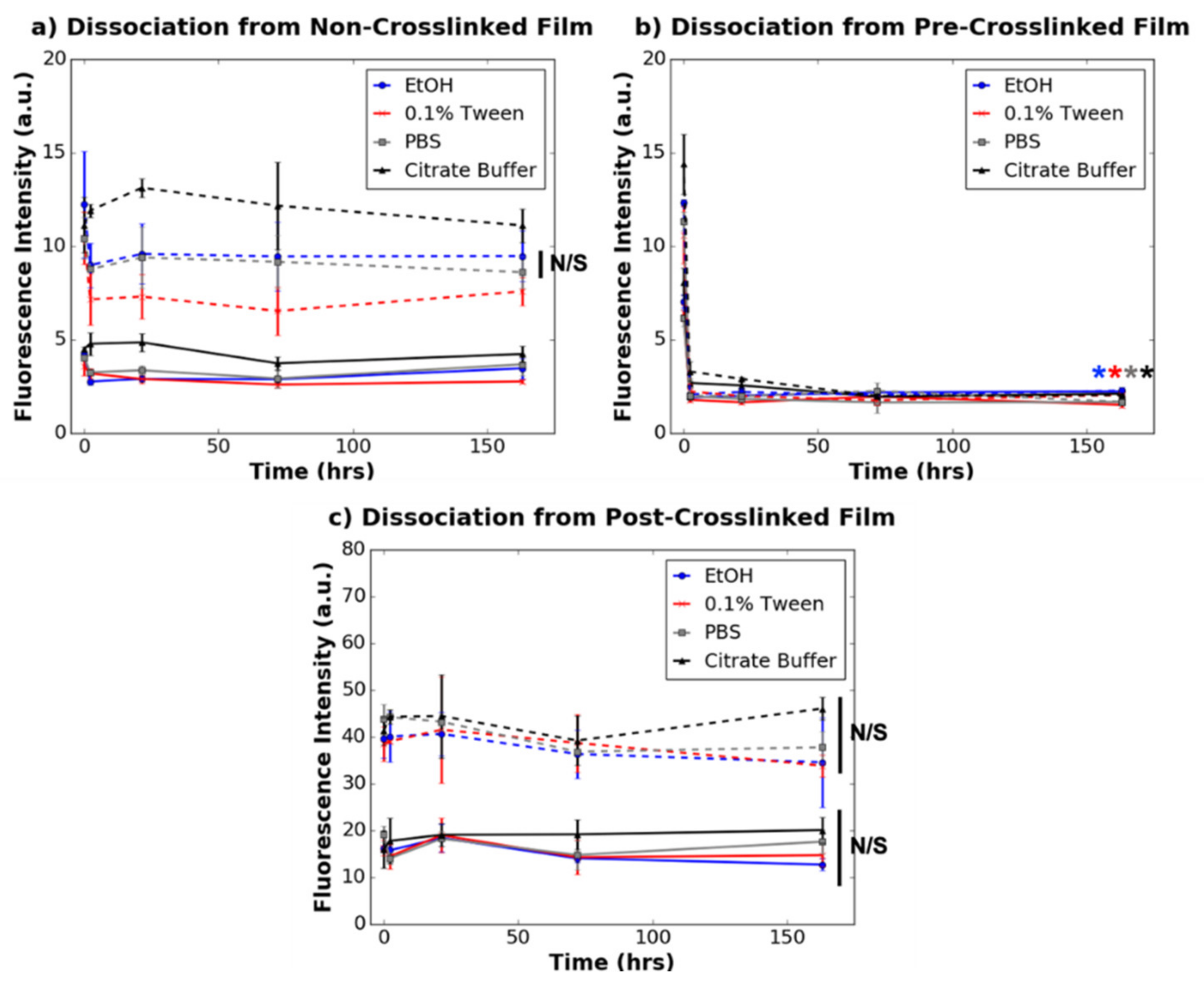
3.3. Dissociation of Dye-Tagged Dendrimers from 2D Collagen Films
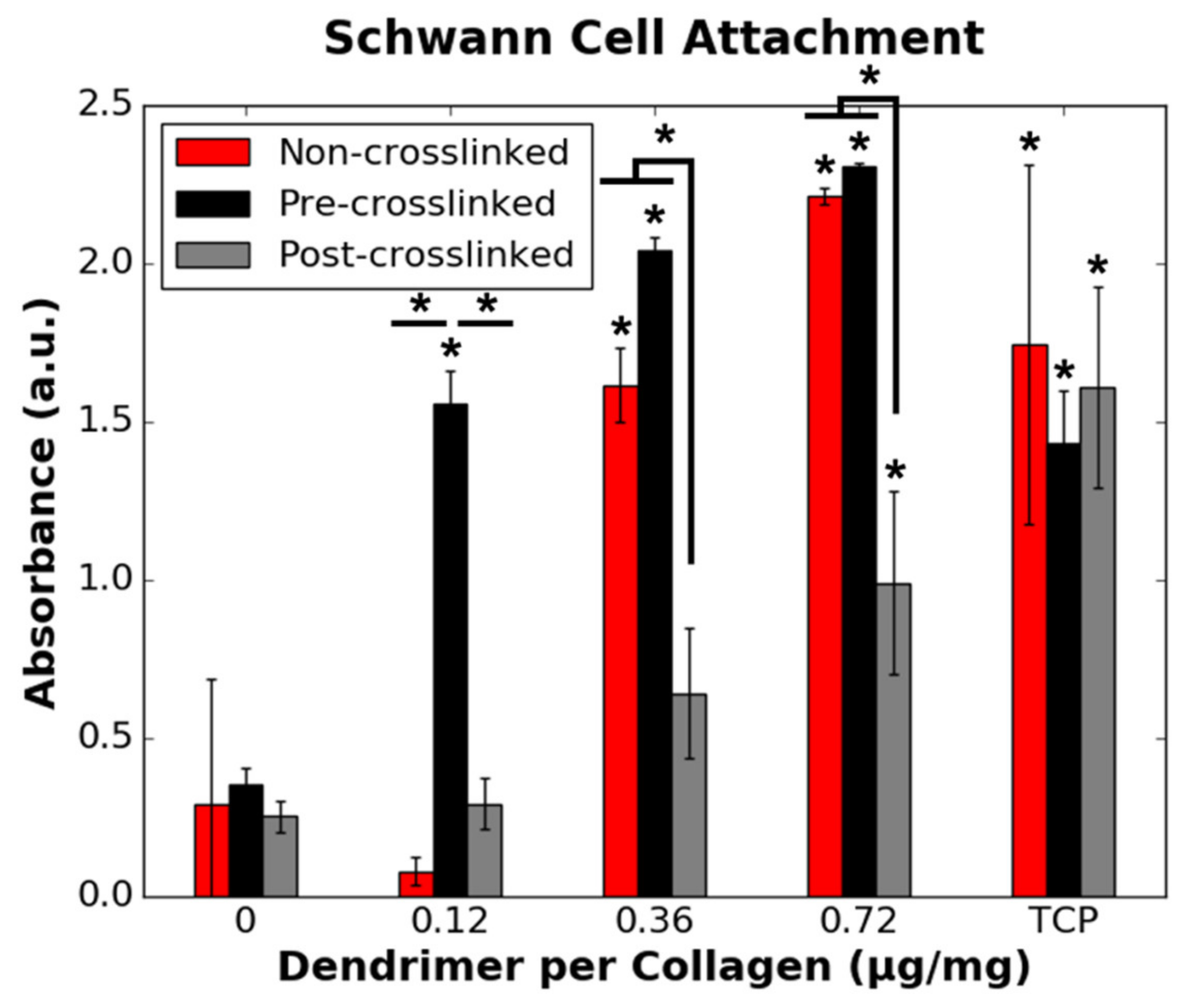
3.4. RSC and HDF Attachment to Dendrimer-Activated Collagen Films
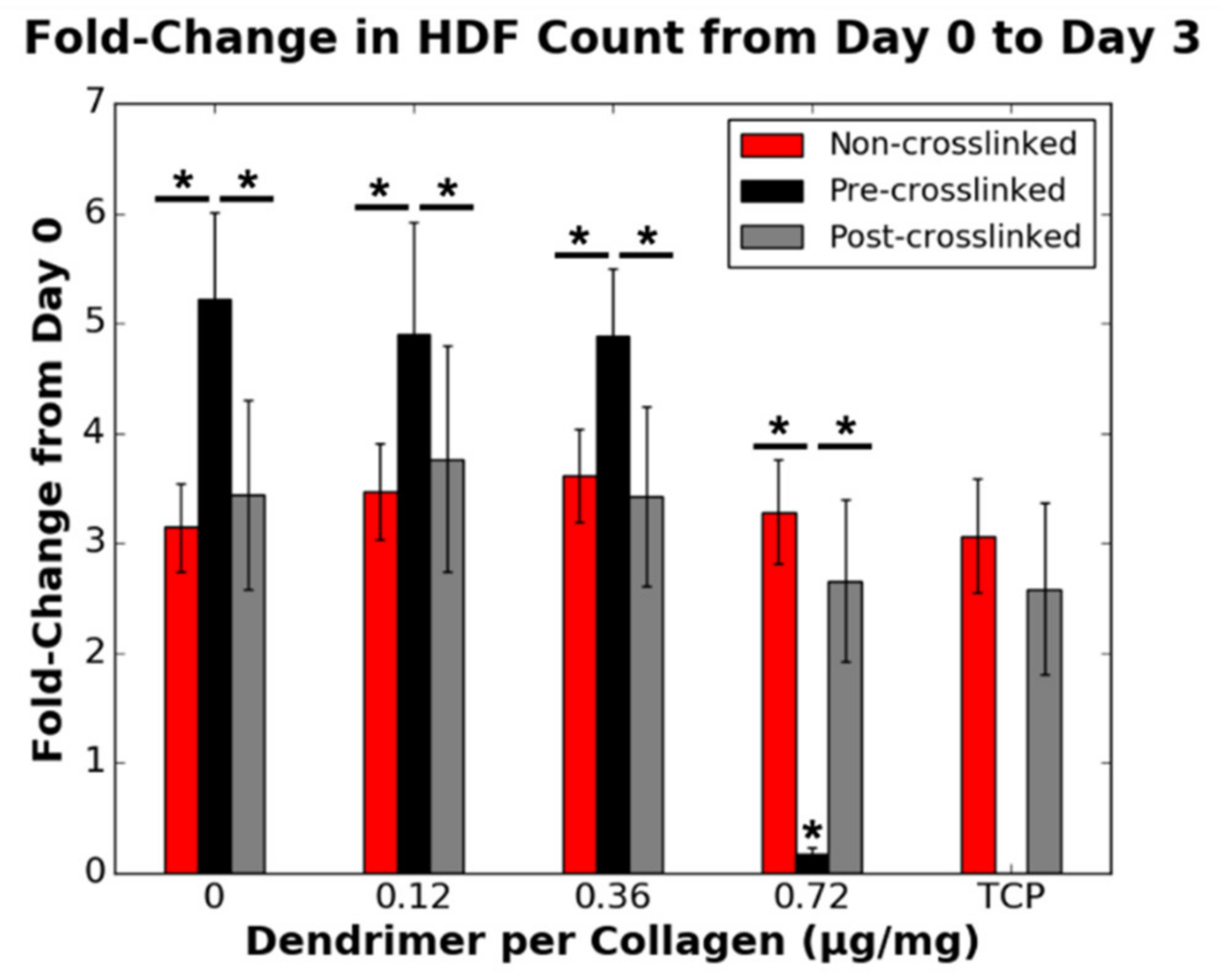
3.5. RSC and HDF Proliferation on Dendrimer-Activated Collagen Films
4. Discussion
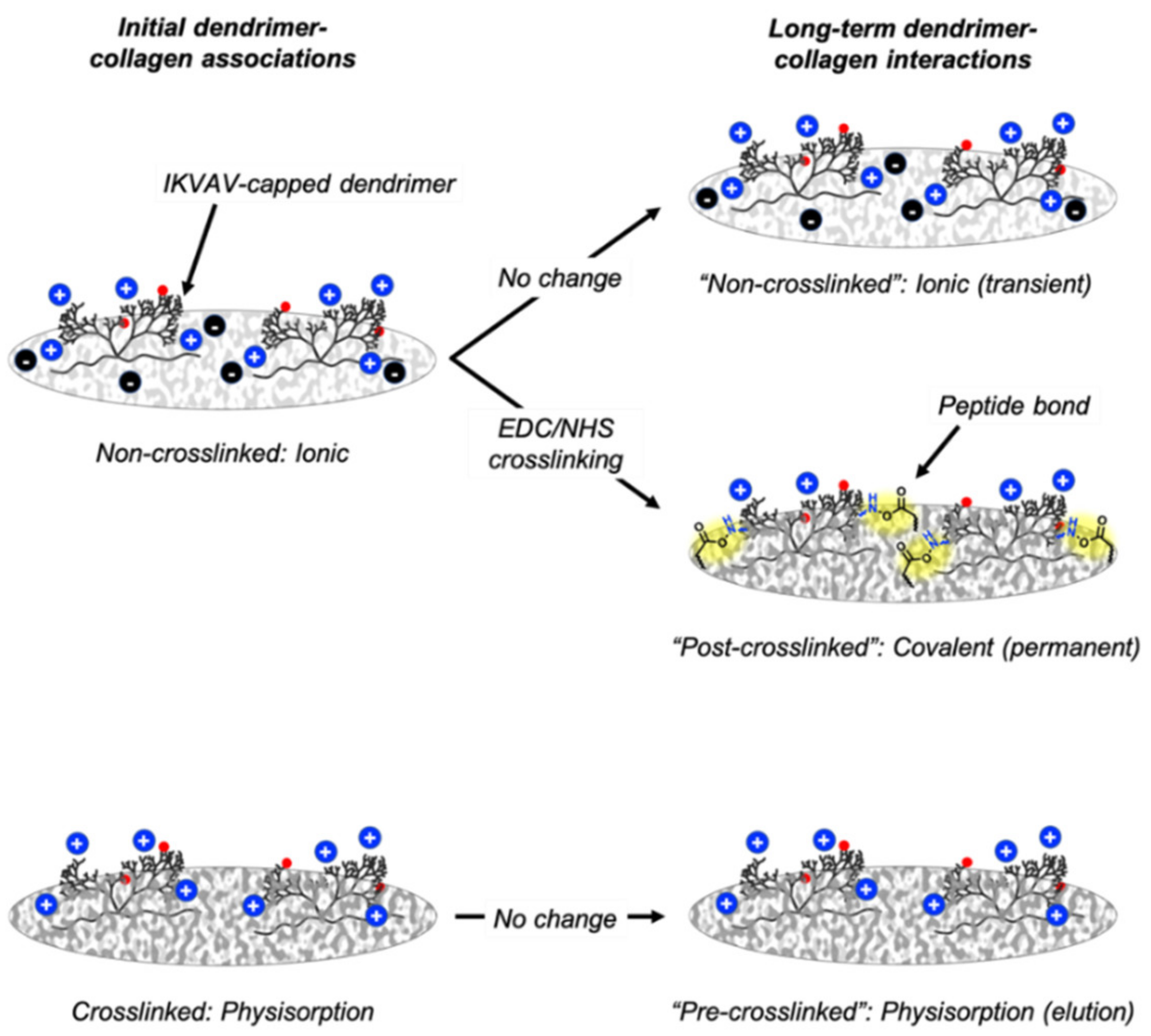
4.1. Dendrimer-Collagen Associations and Long-Term Interactions
4.2. Effect of Dendrimer-Collagen Interactions on Cell Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Daly, W.; Yao, L.; Zeugolis, D.; Windebank, A.; Pandit, A. A biomaterials approach to peripheral nerve regeneration: Bridging the peripheral nerve gap and enhancing functional recovery. J. R. Soc. Interface 2012, 9, 202–221. [Google Scholar] [CrossRef] [Green Version]
- Kubiak, C.A.; Kung, T.A.; Brown, D.L.; Cederna, P.S.; Kemp, S.W.P. State-of-the-Art techniques in treating peripheral nerve injury. Plast. Reconstr. Surg. 2018, 141, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wong, C.; Li, W.; Ruven, C.; He, L.; Wu, X.; Lang, B.T.; Silver, J.; Wu, W. Enhanced regeneration and functional recovery after spinal root avulsion by manipulation of the proteoglycan receptor PTPσ. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.L.; Rivlin, M.; Graham, J.G.; Beredjiklian, P.K. Peripheral nerve injury, scarring, and recovery. Connect. Tissue Res. 2019, 60, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Dreesmann, L.; Mittnacht, U.; Lietz, M.; Schlosshauer, B. Nerve fibroblast impact on Schwann cell behavior. Eur. J. Cell Biol. 2009, 88, 285–300. [Google Scholar] [CrossRef]
- Hinz, B. Tissue stiffness, latent TGF-β1 Activation, and mechanical signal transduction: Implications for the pathogenesis and treatment of fibrosis. Curr. Rheumatol. Rep. 2009, 11, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.; Traka, M.; von Boxberg, Y.; Bouquet, C.; Karagogeos, D.; Nothias, F. Neuronal and glial expression of the adhesion molecule TAG-1 is regulated after peripheral nerve lesion or central neurodegeneration of adult nervous system. Eur. J. Neurosci. 2004, 21, 1169–1180. [Google Scholar] [CrossRef]
- Carvalho, C.R.; Oliveira, J.M.; Reis, R.L. Modern Trends for Peripheral Nerve Repair and Regeneration: Beyond the Hollow Nerve Guidance Conduit. Front. Bioeng. Biotechnol. 2019, 7, 337. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Chen, H.; Qing, L.; Yang, X.; Jia, X. Biomimetic neural scaffolds: A crucial step towards optimal peripheral nerve regeneration. Biomater. Sci. 2018, 6, 1299–1311. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; McLaughlin, C.; Griffith, M.; Sheardown, H. Biofunctionalization of collagen for improved biological response: Scaffolds for corneal tissue engineering. Biomaterials 2007, 28, 78–88. [Google Scholar] [CrossRef]
- Itoh, S.; Takakuda, K.; Samejima, H.; Ohta, T.; Shinomiya, K.; Ichinose, S. Synthetic collagen fibers coated with a synthetic peptide containing the YIGSR sequence of laminin to promote peripheral nerve regeneration in vivo. J. Mater. Sci. Mater. Med. 1999, 10, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Hosseinkhani, H.; Hiraoka, Y.; Li, C.H.; Chen, Y.R.; Yu, D.S.; Hong, P.D.; Ou, K.L. Engineering three-dimensional collagen-IKVAV matrix to mimic neural microenvironment. ACS Chem. Neurosci. 2013, 4, 1229–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massia, S.P.; Holecko, M.M.; Ehteshami, G.R. In vitro assessment of bioactive coatings for neural implant applications. J. Biomed. Mater. Res. Part A 2004, 68, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Perugini, V.; Santin, M. A comparative in vitro study of the effect of biospecific integrin recognition processes and substrate nanostructure on stem cell 3D spheroid formation. J. Mater. Sci. Mater. Med. 2020, 31, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perugini, V.; Meikle, S.T.; Guildford, A.L.; Santin, M. Hyperbranched poly(ϵ-lysine) substrate presenting the laminin sequence YIGSR induces the formation of spheroids in adult bone marrow stem cells. PLoS ONE 2017, 12, e0187182. [Google Scholar] [CrossRef] [Green Version]
- Al-azzawi, S.; Masheta, D.; Guildford, A.; Phillips, G.; Santin, M. Designing and characterization of a novel delivery system for improved cellular uptake by brain using dendronised Apo-E-derived peptide. Front. Bioeng. Biotechnol. 2019, 7, 49. [Google Scholar] [CrossRef]
- Duan, X.; Sheardown, H. Crosslinking of collagen with dendrimers. J. Biomed. Mater. Res. Part A 2005, 75, 510–518. [Google Scholar] [CrossRef]
- Duan, X.; Sheardown, H. Incorporation of cell-adhesion peptides into collagen scaffolds promotes corneal epithelial stratification. J. Biomater. Sci. Polym. Ed. 2007, 18, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Sheardown, H. Dendrimer crosslinked collagen as a corneal tissue engineering scaffold: Mechanical properties and corneal epithelial cell interactions. Biomaterials 2006, 27, 4608–4617. [Google Scholar] [CrossRef] [PubMed]
- Nair, M.; Johal, R.K.; Hamaia, S.W.; Best, S.M.; Cameron, R.E. Tunable bioactivity and mechanics of collagen-based tissue engineering constructs: A comparison of EDC-NHS, genipin and TG2 crosslinkers. Biomaterials 2020, 254, 120109. [Google Scholar] [CrossRef]
- Bax, D.V.; Davidenko, N.; Gullberg, D.; Hamaia, S.W.; Farndale, R.W.; Best, S.M.; Cameron, R.E. Fundamental insight into the effect of carbodiimide crosslinking on cellular recognition of collagen-based scaffolds. Acta Biomater. 2017, 49, 218–234. [Google Scholar] [CrossRef] [PubMed]
- Bax, D.V.; Davidenko, N.; Hamaia, S.W.; Farndale, R.W.; Best, S.M.; Cameron, R.E. Impact of UV- and carbodiimide-based crosslinking on the integrin-binding properties of collagen-based materials. Acta Biomater. 2019, 100, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Olde Damink, L.H.H.; Dijkstra, P.J.; Van Luyn, M.J.A.; Van Wachem, P.B.; Nieuwenhuis, P.; Feijen, J. Cross-linking of dermal sheep collagen using a water-soluble carbodiimide. Biomaterials 1996, 17, 765–773. [Google Scholar] [CrossRef] [Green Version]
- Davidenko, N.; Hamaia, S.; Bax, D.V.; Malcor, J.D.; Schuster, C.F.; Gullberg, D.; Farndale, R.W.; Best, S.M.; Cameron, R.E. Selecting the correct cellular model for assessing of the biological response of collagen-based biomaterials. Acta Biomater. 2018, 65, 88–101. [Google Scholar] [CrossRef]
- Lee, J.M.; Edwards, H.H.L.; Pereira, C.A.; Samii, S.I. Crosslinking of tissue-derived biomaterials in 1-ethyl-3-(3-dimethylaminopropyl)-carbodiimide (EDC). J. Mater. Sci. Mater. Med. 1996, 7, 531–541. [Google Scholar] [CrossRef]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2010, 339, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Knight, C.G.; Morton, L.F.; Peachey, A.R.; Tuckwell, D.S.; Farndale, R.W.; Barnes, M.J. The collagen-binding a-domains of integrins α1/β1 and α2/β1 recognize the same specific amino acid sequence, GFOGER, in native (triple-helical) collagens. J. Biol. Chem. 2000, 275, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Davidenko, N.; Schuster, C.F.; Bax, D.V.; Farndale, R.W.; Hamaia, S.; Best, S.M.; Cameron, R.E. Evaluation of cell binding to collagen and gelatin: A study of the effect of 2D and 3D architecture and surface chemistry. J. Mater. Sci. Mater. Med. 2016, 27, 148. [Google Scholar] [CrossRef] [Green Version]
- Grover, C.N.; Gwynne, J.H.; Pugh, N.; Hamaia, S.; Farndale, R.W.; Best, S.M.; Cameron, R.E. Crosslinking and composition influence the surface properties, mechanical stiffness and cell reactivity of collagen-based films. Acta Biomater. 2012, 8, 3080–3090. [Google Scholar] [CrossRef] [Green Version]
- Bella, J.; Brodsky, B.; Berman, H.M. Hydration structure of a collagen peptide. Structure 1995, 3, 893–906. [Google Scholar] [CrossRef] [Green Version]
- Lide, D.R. Handbook of Chemistry and Physics, 72nd ed.; CRC Press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Roberts, D.; Keeling, R.; Tracka, M.; van der Walle, C.F.; Uddin, S.; Warwicker, J.; Curtis, R. Specific Ion and Buffer Effects on Protein-Protein Interactions of a Monoclonal Antibody. Mol. Pharm. 2015, 12, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Davidenko, N.; Schuster, C.F.; Bax, D.V.; Raynal, N.; Farndale, R.W.; Best, S.M.; Cameron, R.E. Control of crosslinking for tailoring collagen-based scaffolds stability and mechanics. Acta Biomater. 2015, 25, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Grover, C.N.; Farndale, R.W.; Best, S.M.; Cameron, R.E. The interplay between physical and chemical properties of protein films affects their bioactivity. J. Biomed. Mater. Res. Part A 2012, 100, 2401–2411. [Google Scholar] [CrossRef]
- Kasai, S.; Ohga, Y.; Mochizuki, M.; Nishi, N.; Kadoya, Y.; Nomizu, M. Multifunctional Peptide Fibrils for Biomedical Materials. Biopolymers 2004, 76, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Emsley, J.; Knight, C.G.; Farndale, R.W.; Barnes, M.J. Structure of the Integrin α2β1-binding Collagen Peptide. J. Mol. Biol. 2004, 335, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Shi-wen, X.; Blumbach, K.; Eastwood, M.; Denton, C.P.; Eckes, B.; Krieg, T.; Abraham, D.J.; Leask, A. Expression of integrin β1 by fibroblasts is required for tissue repair in vivo. J. Cell Sci. 2010, 123, 3674–3682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khew, S.T.; Tong, Y.W. The Specific Recognition of a Cell Binding Sequence Derived from Type I Collagen by Hep3B and L929 Cells. Biomacromolecules 2007, 8, 3153–3161. [Google Scholar] [CrossRef] [PubMed]
- Farndale, R.W.; Lisman, T.; Bihan, D.; Hamaia, S.; Smerling, C.S.; Pugh, N.; Konitsiotis, A.; Leitinger, B.; De Groot, P.G.; Jarvis, G.E.; et al. Cell-collagen interactions: The use of peptide Toolkits to investigate collagen-receptor interactions. Biochem. Soc. Trans. 2008, 36, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Schéele, S.; Nyström, A.; Durbeej, M.; Talts, J.F.; Ekblom, M.; Ekblom, P. Laminin isoforms in development and disease. J. Mol. Med. 2007, 85, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Hadjipanayi, E.; Mudera, V.; Brown, R.A. Close dependence of fibroblast proliferation on collagen scaffold matrix stiffness. J. Tissue Eng. Regen. Med. 2009, 3, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Kleinman, H.K.; Weeks, B.S.; Cannon, F.B.; Sweeney, T.M.; Sephel, G.C.; Clement, B.; Zain, M.; Olson, M.O.J.; Jucker, M.; Burrous, B.A. Identification of a 110-kDa nonintegrin cell surface laminin-binding protein which recognizes an a chain neurite-promoting peptide. Arch. Biochem. Biophys. 1991, 290, 320–325. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.J.; Bax, D.V.; Murphy, R.; Best, S.M.; Cameron, R.E. Collagen Film Activation with Nanoscale IKVAV-Capped Dendrimers for Selective Neural Cell Response. Nanomaterials 2021, 11, 1157. https://doi.org/10.3390/nano11051157
Kim JJ, Bax DV, Murphy R, Best SM, Cameron RE. Collagen Film Activation with Nanoscale IKVAV-Capped Dendrimers for Selective Neural Cell Response. Nanomaterials. 2021; 11(5):1157. https://doi.org/10.3390/nano11051157
Chicago/Turabian StyleKim, Jessica J., Daniel V. Bax, Robert Murphy, Serena M. Best, and Ruth E. Cameron. 2021. "Collagen Film Activation with Nanoscale IKVAV-Capped Dendrimers for Selective Neural Cell Response" Nanomaterials 11, no. 5: 1157. https://doi.org/10.3390/nano11051157