
Influence of Copper Oxide Nanoparticles on Gene Expression of Birch Clones In Vitro under Stress Caused by Phytopathogens

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Nanoparticles and Nanoparticles Suspensions
2.2. Determination of Antifungal Activity
- where D is the diameter of the fungus colony in the control variant;
- d is the diameter of the NPs-treated fungus colony.
2.3. Cultivation and Artificial Infection
2.4. Evaluation of the Influence of A. alternata and CuO NPs on the Expression of Stress Resistance Genes in Regenerants of Downy Birch
2.4.1. RNA Extraction and Evaluation
2.4.2. cDNA Synthesis and Primers Picking
2.4.3. PCR Analysis and Assessment of the Stress Genes Expression
2.5. Statistical Analysis
3. Results
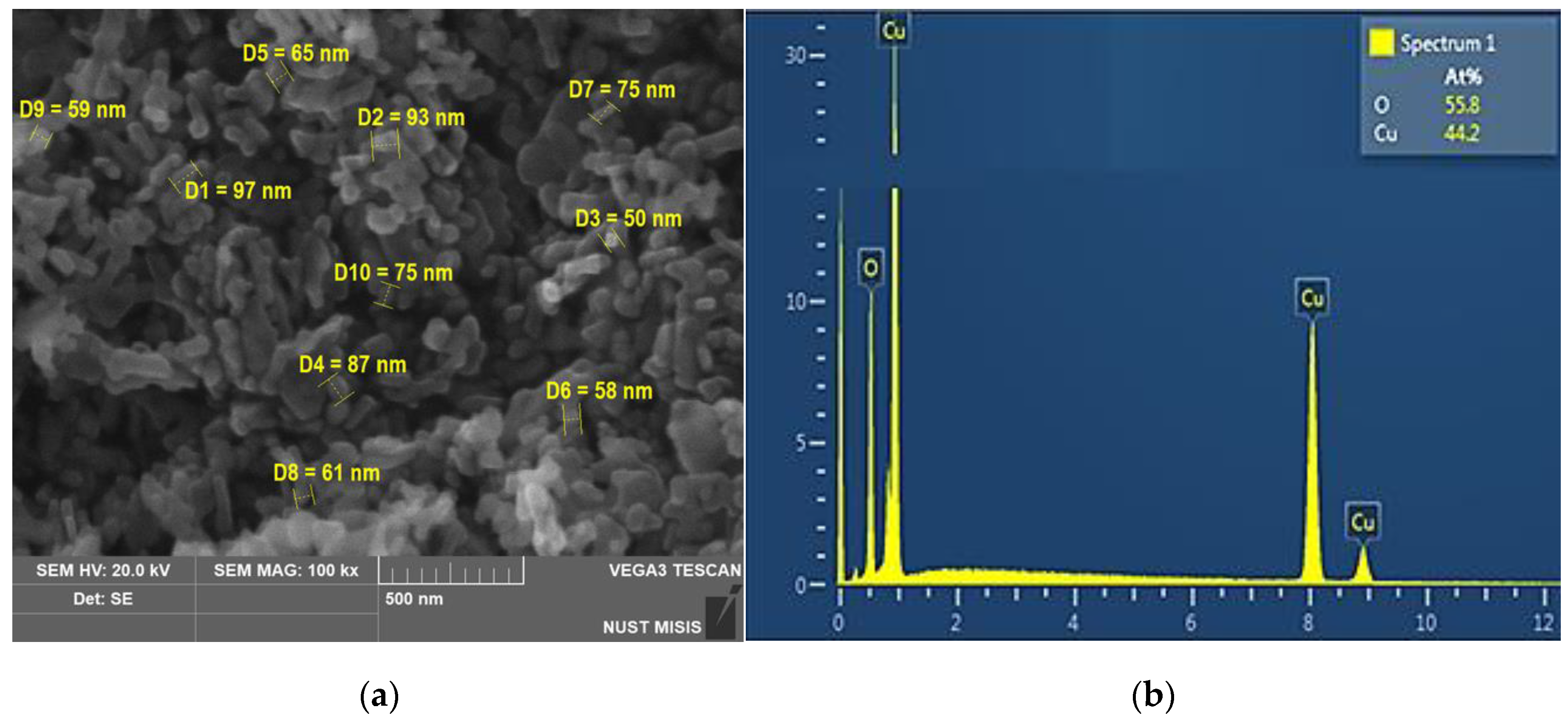
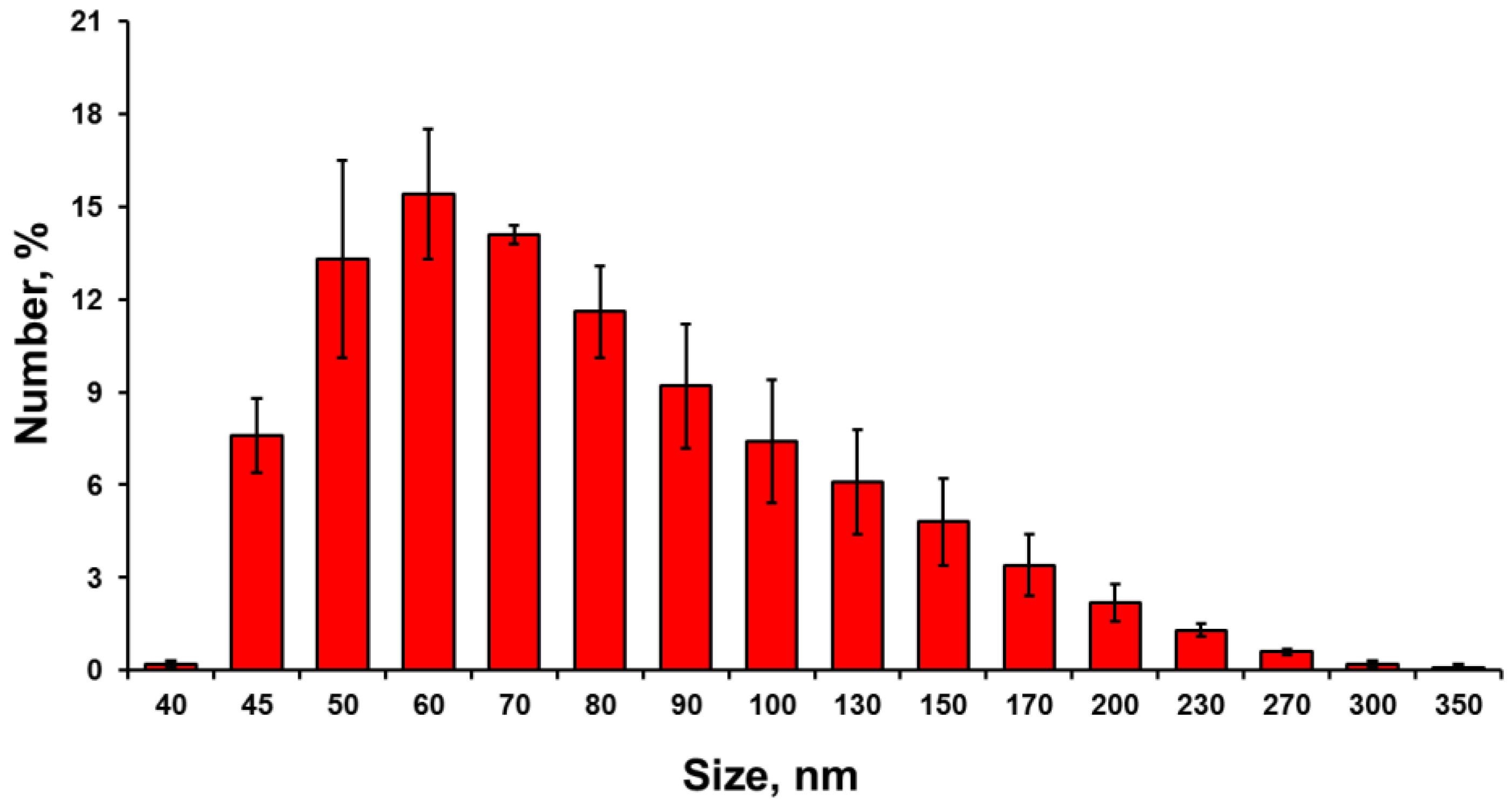
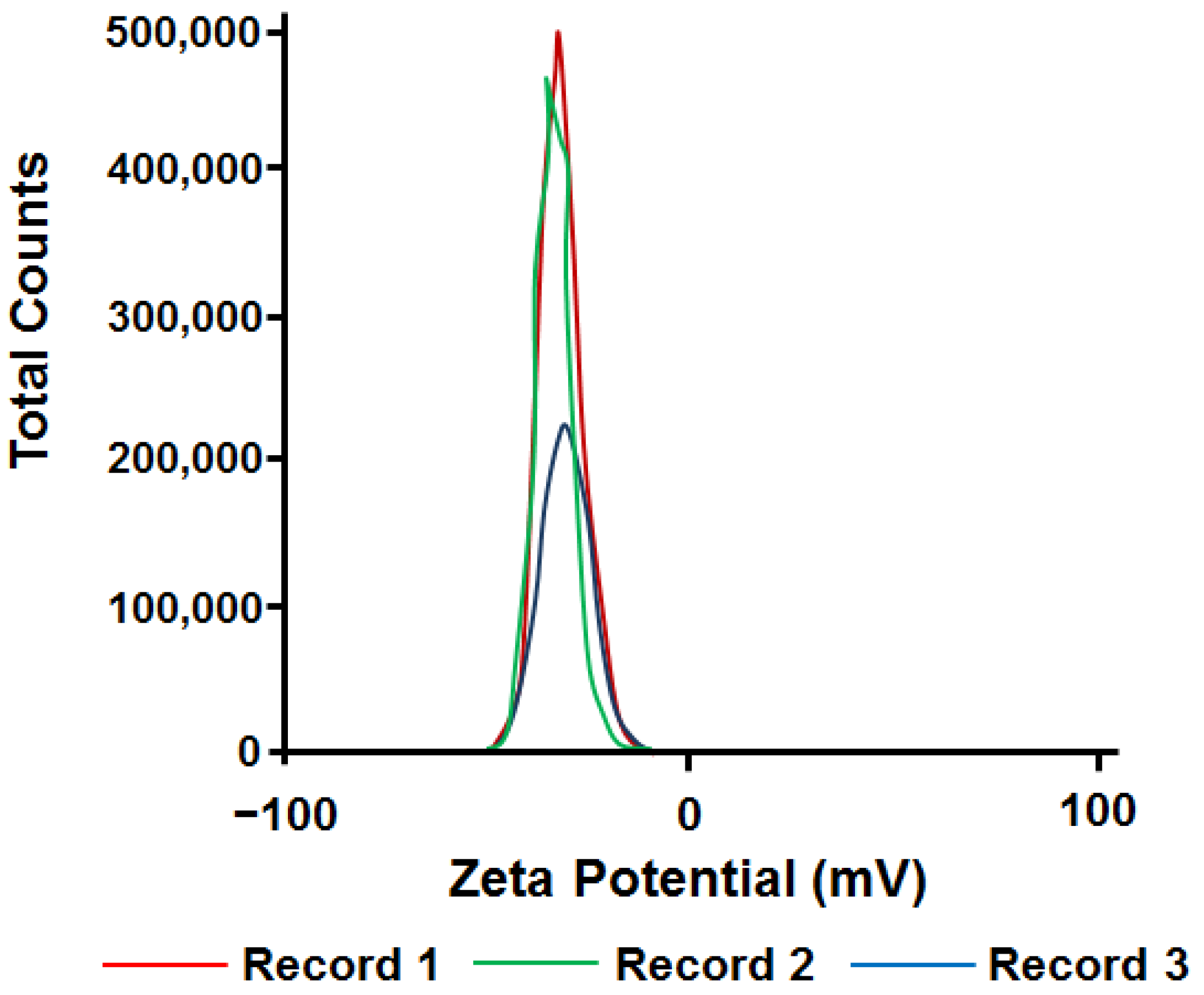
3.1. Nanoparticles Characterization
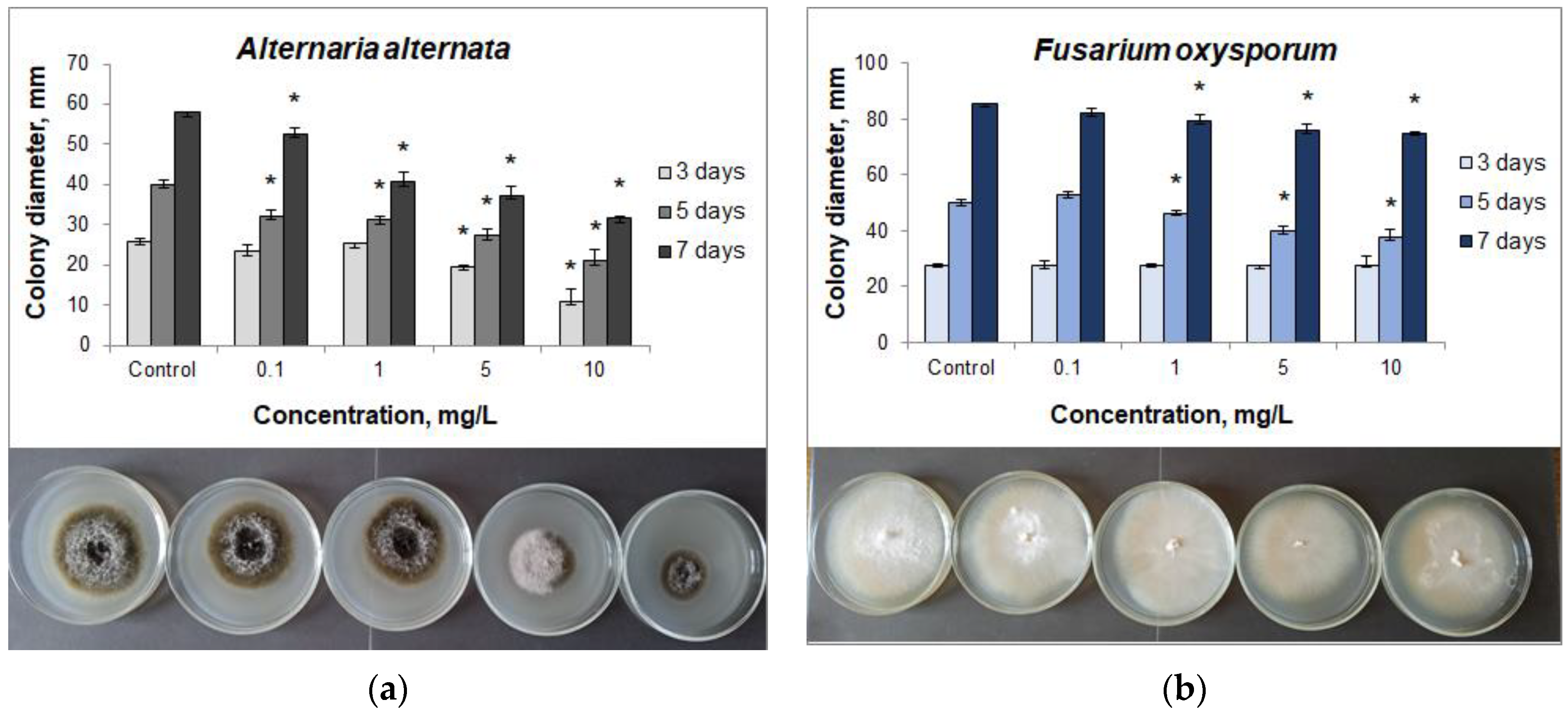
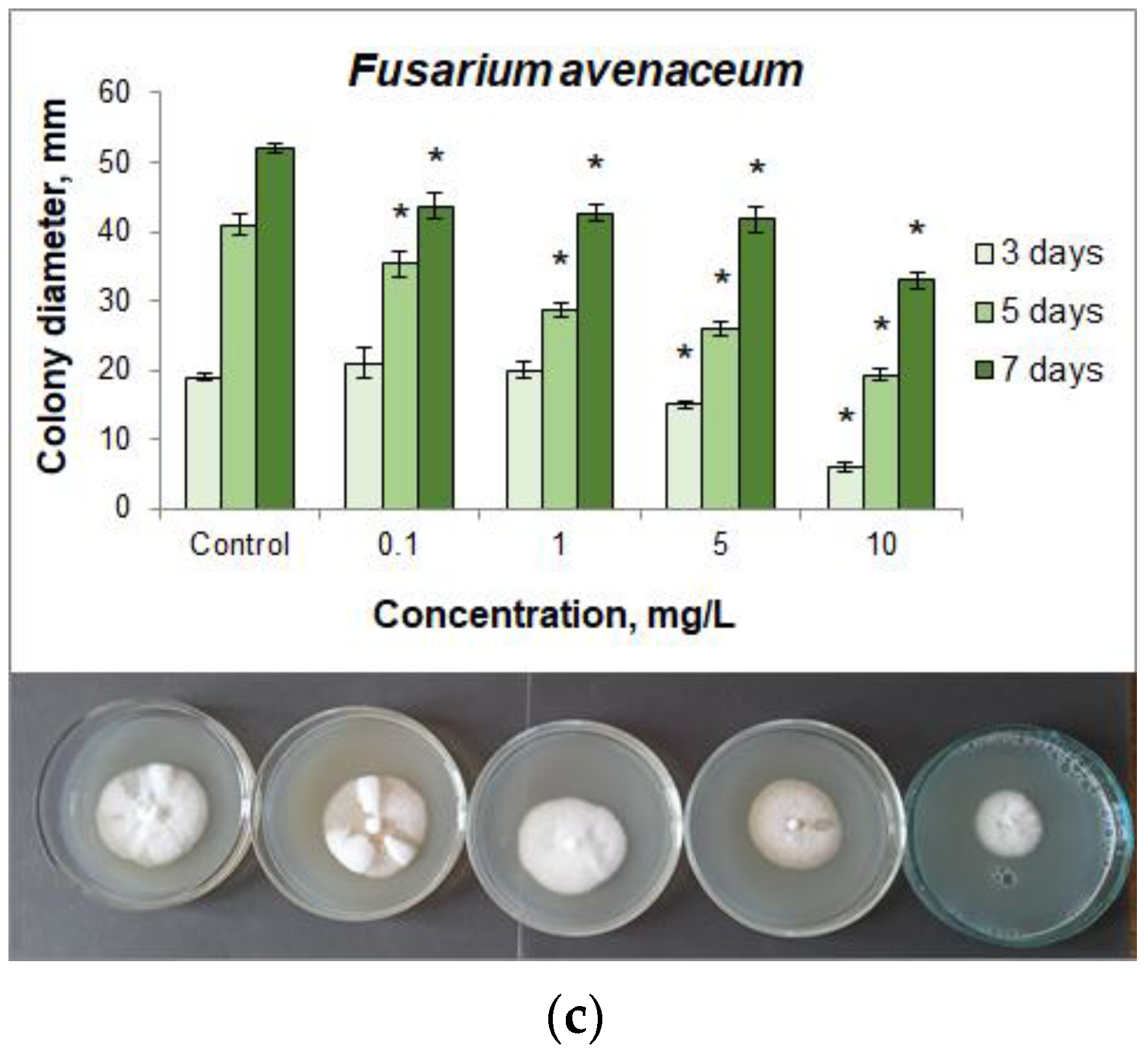
3.2. Fungicidal Action of CuO NPs
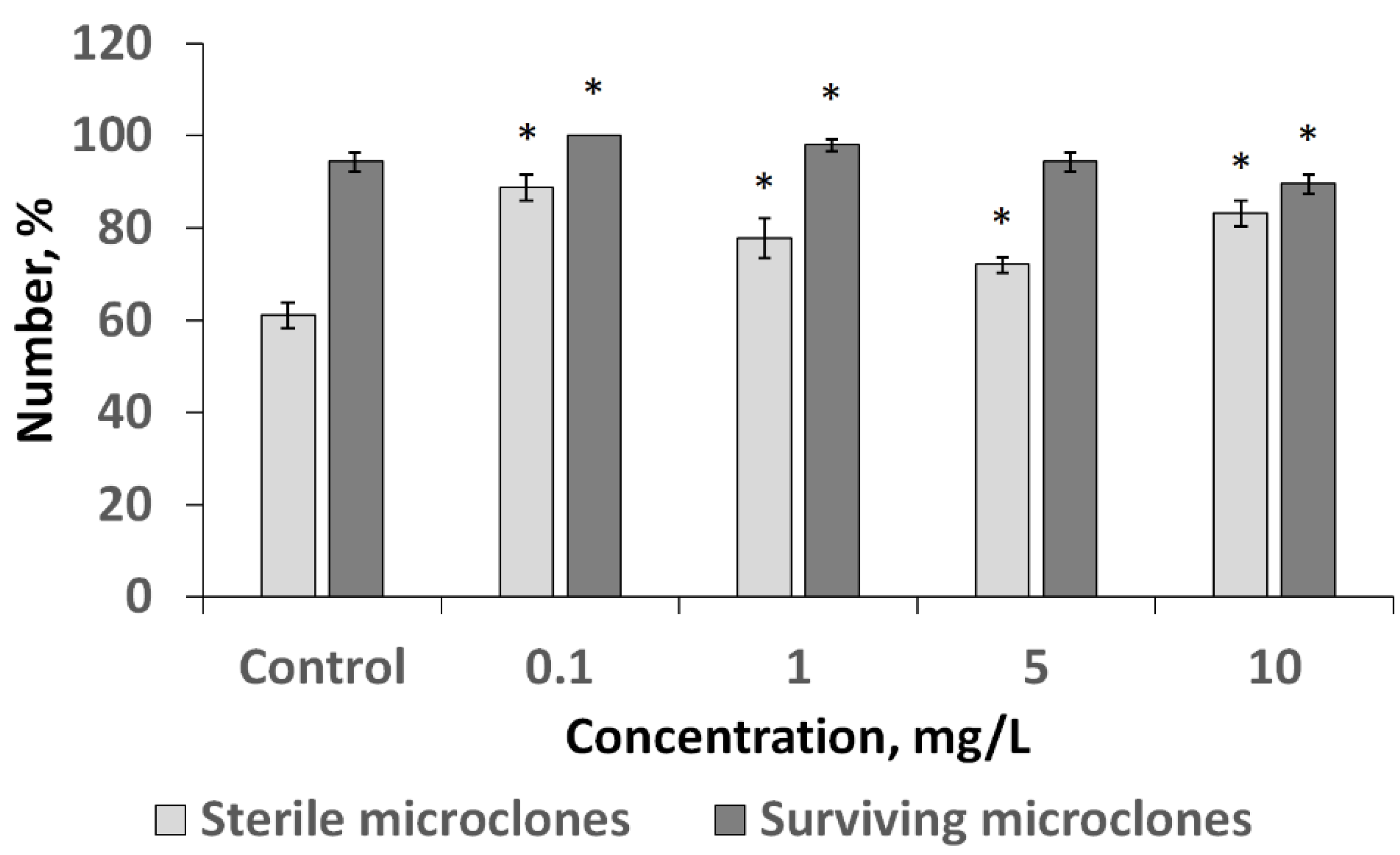
3.3. Influence of CuO NPs on Infected Birch Explants at the Stage of Introduction into Culture
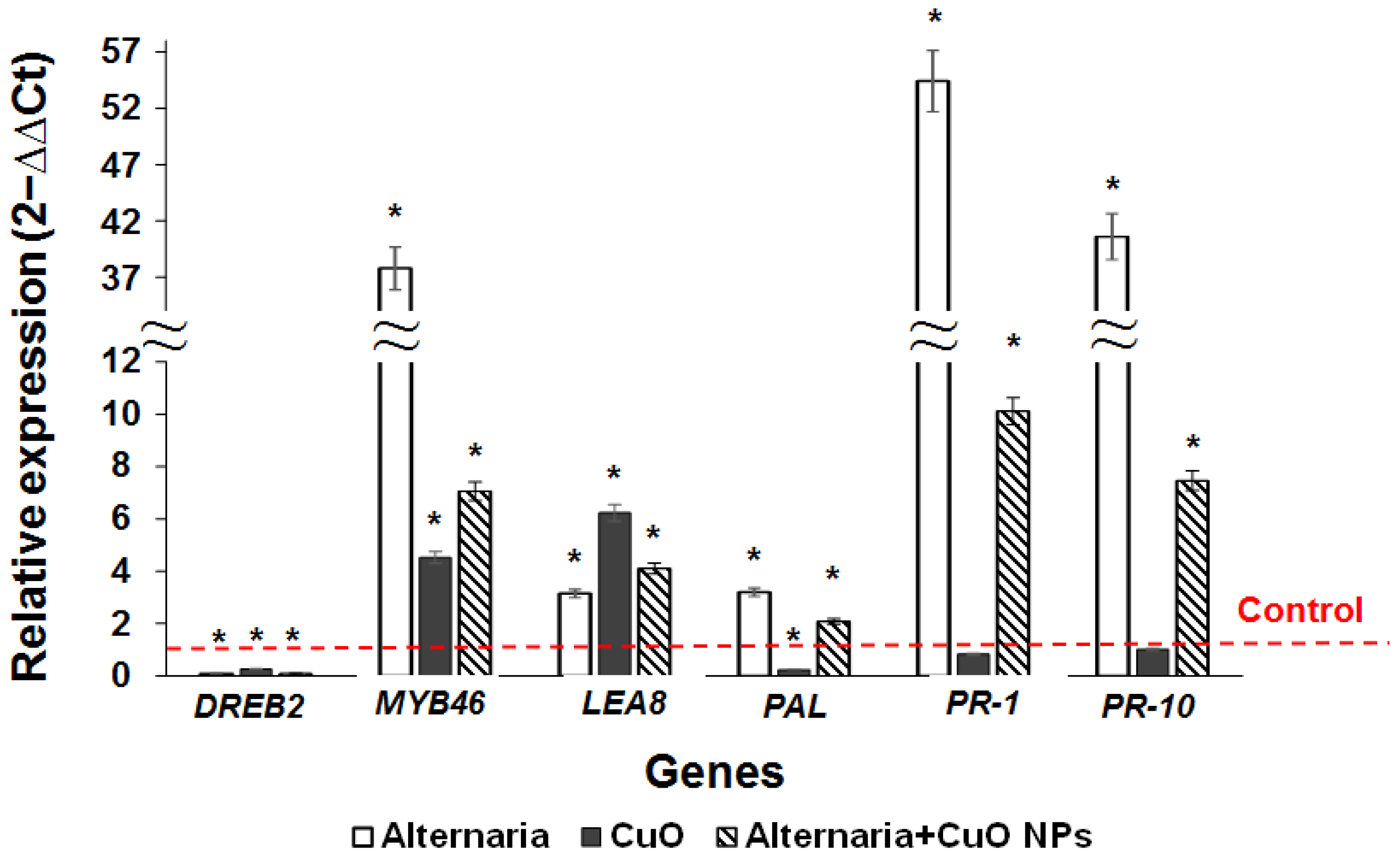
3.4. Influence of A. alternata and CuO NPs on the Expression of Stress Resistance Genes in Regenerants of Downy Birch
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Reed, B.M.; Mentzer, J.; Tanprasert, P.; Yu, X. Internal Bacterial Contamination of Micropropagated Hazelnut: Identification and Antibiotic Treatment. Plant Cell Tissue Organ Cult. 1998, 52, 67–70. [Google Scholar] [CrossRef]
- Pollock, K.; Barfield, D.G.; Shields, R. The toxicity of antibiotics to plant cell cultures. Plant Cell Rep. 1983, 2, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. J. Chem. Biol. 2014, 7, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, P.M.M.; Merfa, M.V.; Takita, M.A.; De Souza, A.A. Persistence in Phytopathogenic Bacteria: Do We Know Enough? Front. Microbiol. 2018, 9, 1099. [Google Scholar] [CrossRef]
- Teow, S.-Y.; Wong, M.M.-T.; Yap, H.-Y.; Peh, S.-C.; Shameli, K. Bactericidal Properties of Plants-Derived Metal and Metal Oxide Nanoparticles (NPs). Molecules 2018, 23, 1366. [Google Scholar] [CrossRef] [Green Version]
- Zakharova, O.; Gusev, A. Photocatalytically Active Zinc Oxide and Titanium Dioxide Nanoparticles in Clonal Micropropagation of Plants: Prospects. Nanotechnol. Russ. 2019, 14, 311–324. [Google Scholar] [CrossRef]
- Ur Rahim, H.; Qaswar, M.; Uddin, M.; Giannini, C.; Herrera, M.L.; Rea, G. Nano-Enable Materials Promoting Sustainability and Resilience in Modern Agriculture. Nanomaterials 2021, 11, 2068. [Google Scholar] [CrossRef]
- Singh, H.; Sharma, A.; Bhardwaj, S.K.; Arya, S.K.; Bhardwaj, N.; Khatri, M. Recent advances in the applications of nano-agrochemicals for sustainable agricultural development. Environ. Sci. Processes Impacts 2021, 23, 213–239. [Google Scholar] [CrossRef]
- Singh, R.P.; Handa, R.; Manchanda, G. Nanoparticles in sustainable agriculture: An emerging opportunity. J. Control. Release 2021, 329, 1234–1248. [Google Scholar] [CrossRef]
- Kim, D.Y.; Kadam, A.; Shinde, S.; Saratale, R.G.; Patra, J.; Ghodake, G. Recent developments in nanotechnology transforming the agricultural sector: A transition replete with opportunities. J. Sci. Food Agric. 2018, 98, 849–864. [Google Scholar] [CrossRef]
- Ali, S.S.; Al-Tohamy, R.; Koutra, E.; Moawad, M.S.; Kornaros, M.; Mustafa, A.M.; Mahmoud, Y.A.G.; Badr, A.; Osman, M.E.H.; Elsamahy, T.; et al. Nanobiotechnological advancements in agriculture and food industry: Applications, nanotoxicity, and future perspectives. Sci. Total Environ. 2021, 792, 148359. [Google Scholar] [CrossRef] [PubMed]
- Ermini, M.L.; Voliani, V. Antimicrobial Nano-Agents: The Copper Age. ACS Nano 2021, 15, 6008–6029. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, K.S.; Husen, A. Current status of plant metabolite-based fabrication of copper/copper oxide nanoparticles and their applications: A review. Biomater. Res. 2020, 24, 11. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.; Sharma, A.; Bachheti, R.K.; Husen, A.; Mishra, V.K. Plant-Mediated Synthesis of Copper Oxide Nanoparticles and Their Biological Applications. In Nanomaterials and Plant Potential; Husen, A., Iqbal, M., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 221–237. [Google Scholar]
- Malandrakis, A.A.; Kavroulakis, N.; Chrysikopoulos, C.V. Use of copper, silver and zinc nanoparticles against foliar and soil-borne plant pathogens. Sci. Total Environ. 2019, 670, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Juárez Maldonado, A.; Ortega-Ortíz, H.; Pérez-Labrada, F.; Cadenas-Pliego, G.; Benavides-Mendoza, A. Cu Nanoparticles absorbed on chitosan hydrogels positively alter morphological, production, and quality characteristics of tomato. J. Appl. Bot. Food Qual. 2016, 89, 183–189. [Google Scholar] [CrossRef]
- Genady, E.; Qaid, E.A.; Fahmy, A. Copper Sulfate Nanoparticales In vitro Applications on Verbena bipinnatifida Nutt. Stimulating Growth and Total Phenolic Content Increasments. Int. J. Pharm. Res. Allied Sci. 2016, 5, 196–202. [Google Scholar]
- Talankova-Sereda, T.; Liapina, K.; Shkopinskij, E.; Ustinov, A.; Kovalyova, A.; Dulnev, P.; Kucenko, N. The Influence of Cu и Co Nanoparticles on Growth Characteristics and Biochemical Structure of Mentha Longifolia In Vitro. In Nanophysics, Nanophotonics, Surface Studies, and Applications; Springer: Cham, Switzerland, 2016; Volume 183, pp. 427–436. [Google Scholar]
- Rivero-Montejo, S.d.J.; Vargas-Hernandez, M.; Torres-Pacheco, I. Nanoparticles as Novel Elicitors to Improve Bioactive Compounds in Plants. Agriculture 2021, 11, 134. [Google Scholar] [CrossRef]
- Naseer, I.; Javad, S.; Singh, A.; Maqsood, S.; Iqbal, S.; Jabeen, K. Alleviation Mechanism of Drought Stress in Plants Using Metal Nanoparticles—A Perspective Analysis. In Sustainable Agriculture Reviews 53: Nanoparticles: A New Tool to Enhance Stress Tolerance; Faizan, M., Hayat, S., Yu, F., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 115–149. [Google Scholar]
- Marslin, G.; Sheeba, C.J.; Franklin, G. Nanoparticles Alter Secondary Metabolism in Plants via ROS Burst. Front. Plant Sci. 2017, 8, 832. [Google Scholar] [CrossRef] [Green Version]
- Chun, S.C.; Chandrasekaran, M. Chitosan and chitosan nanoparticles induced expression of pathogenesis-related proteins genes enhances biotic stress tolerance in tomato. Int. J. Biol. Macromol. 2019, 125, 948–954. [Google Scholar] [CrossRef]
- Zeeshan, M.; Hu, Y.X.; Afridi, M.S.; Ahmad, B.; Ahmad, S.; Muhammad, I.; Hale, B.; Iqbal, A.; Farooq, S.; Wu, H.Y.; et al. Interplay of ZnONPs and/or SeNPs induces arsenic tolerance in soybean by regulation of antioxidants pool, WRKY genes, and expression of arsenic transporters. Environ. Exp. Bot. 2022, 195, 104783. [Google Scholar] [CrossRef]
- Yang, K.-Y.; Doxey, S.; McLean, J.E.; Britt, D.; Watson, A.; Qassy, D.A.; Jacobson, A.; Anderson, A.J. Remodeling of root morphology by CuO and ZnO nanoparticles: Effects on drought tolerance for plants colonized by a beneficial pseudomonad. Botany 2018, 96, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Linh, T.M.; Mai, N.C.; Hoe, P.T.; Lien, L.Q.; Ban, N.K.; Hien, L.T.T.; Chau, N.H.; Van, N.T. Metal-Based Nanoparticles Enhance Drought Tolerance in Soybean. J. Nanomater. 2020, 2020, 4056563. [Google Scholar] [CrossRef]
- Saleh, A.M.; Hassan, Y.M.; Selim, S.; AbdElgawad, H. NiO-nanoparticles induce reduced phytotoxic hazards in wheat (Triticum aestivum L.) grown under future climate CO2. Chemosphere 2019, 220, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- García-Sánchez, S.; Gala, M.; Žoldák, G. Nanoimpact in Plants: Lessons from the Transcriptome. Plants 2021, 10, 751. [Google Scholar] [CrossRef]
- Mosa, K.A.; El-Naggar, M.; Ramamoorthy, K.; Alawadhi, H.; Elnaggar, A.; Wartanian, S.; Ibrahim, E.; Hani, H. Copper Nanoparticles Induced Genotoxicty, Oxidative Stress, and Changes in Superoxide Dismutase (SOD) Gene Expression in Cucumber (Cucumis sativus) Plants. Front. Plant Sci. 2018, 9, 872. [Google Scholar] [CrossRef] [Green Version]
- Jośko, I.; Kusiak, M.; Xing, B.; Oleszczuk, P. Combined effect of nano-CuO and nano-ZnO in plant-related system: From bioavailability in soil to transcriptional regulation of metal homeostasis in barley. J Hazard Mater 2021, 416, 126230. [Google Scholar] [CrossRef]
- Petrova, A.; Plaksenkova, I.; Kokina, I.; Jermaļonoka, M. Effect of Fe3O4 and CuO Nanoparticles on Morphology, Genotoxicity, and miRNA Expression on Different Barley (Hordeum vulgare L.) Genotypes. Sci. World J. 2021, 2021, 6644689. [Google Scholar] [CrossRef]
- Cota-Ruiz, K.; Hernández-Viezcas, J.A.; Varela-Ramírez, A.; Valdés, C.; Núñez-Gastélum, J.A.; Martínez-Martínez, A.; Delgado-Rios, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Toxicity of copper hydroxide nanoparticles, bulk copper hydroxide, and ionic copper to alfalfa plants: A spectroscopic and gene expression study. Environ. Pollut. 2018, 243, 703–712. [Google Scholar] [CrossRef]
- Elmer, W.; De La Torre-Roche, R.; Pagano, L.; Majumdar, S.; Zuverza-Mena, N.; Dimkpa, C.; Gardea-Torresdey, J.; White, J.C. Effect of Metalloid and Metal Oxide Nanoparticles on Fusarium Wilt of Watermelon. Plant Dis. 2018, 102, 1394–1401. [Google Scholar] [CrossRef] [Green Version]
- El-Garhy, H.A.S.; Elsisi, A.A.; Mohamed, S.A.; Morsy, O.M.; Osman, G.; Abdel-Rahman, F.A. Transcriptomic changes in green bean pods against grey mould and white rot diseases via field application of chemical elicitor nanoparticles. IET Nanobiotechnol. 2020, 14, 574–583. [Google Scholar] [CrossRef]
- Nair, P.M.; Chung, I.M. Study on the correlation between copper oxide nanoparticles induced growth suppression and enhanced lignification in Indian mustard (Brassica juncea L.). Ecotoxicol. Environ. Saf. 2015, 113, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Landa, P.; Dytrych, P.; Prerostova, S.; Petrova, S.; Vankova, R.; Vanek, T. Transcriptomic Response of Arabidopsis thaliana Exposed to CuO Nanoparticles, Bulk Material, and Ionic Copper. Environ. Sci. Technol. 2017, 51, 10814–10824. [Google Scholar] [CrossRef] [PubMed]
- Ke, M.; Zhu, Y.; Zhang, M.; Gumai, H.; Zhang, Z.; Xu, J.; Qian, H. Physiological and Molecular Response of Arabidopsis thaliana to CuO Nanoparticle (nCuO) Exposure. Bull. Environ. Contam. Toxicol. 2017, 99, 713–718. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, L.; Zhao, J.; Wang, X.; White, J.C.; Xing, B. CuO Nanoparticle Interaction with Arabidopsis thaliana: Toxicity, Parent-Progeny Transfer, and Gene Expression. Environ. Sci. Technol. 2016, 50, 6008–6016. [Google Scholar] [CrossRef]
- Wang, S.; Liu, H.; Zhang, Y.; Xin, H. The effect of CuO NPs on reactive oxygen species and cell cycle gene expression in roots of rice. Environ. Toxicol. Chem. 2015, 34, 554–561. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, J.; Rui, M.; Yang, L.; Shen, J.; Chu, H.; Song, S.; Chen, Y. OsFTIP7 determines metallic oxide nanoparticles response and tolerance by regulating auxin biosynthesis in rice. J. Hazard. Mater. 2021, 403, 123946. [Google Scholar] [CrossRef]
- Nair, P.M.; Chung, I.M. A mechanistic study on the toxic effect of copper oxide nanoparticles in soybean (Glycine max L.) root development and lignification of root cells. Biol. Trace Elem. Res. 2014, 162, 342–352. [Google Scholar] [CrossRef]
- Servin, A.D.; Pagano, L.; Castillo-Michel, H.; De la Torre-Roche, R.; Hawthorne, J.; Hernandez-Viezcas, J.A.; Loredo-Portales, R.; Majumdar, S.; Gardea-Torresday, J.; Dhankher, O.P.; et al. Weathering in soil increases nanoparticle CuO bioaccumulation within a terrestrial food chain. Nanotoxicology 2017, 11, 98–111. [Google Scholar] [CrossRef]
- Anderson, A.J.; McLean, J.E.; Jacobson, A.R.; Britt, D.W. CuO and ZnO Nanoparticles Modify Interkingdom Cell Signaling Processes Relevant to Crop Production. J. Agric. Food Chem. 2018, 66, 6513–6524. [Google Scholar] [CrossRef]
- Grodetskaya, T.; Fedorova, O.; Evlakov, P.; Baranov, O.; Zakharova, O.; Gusev, A. Effect of Copper Oxide and Silver Nanoparticles on the Development of Tolerance to Alternaria alternata in Poplar in Vitro Clones. Nanobiotechnol. Rep. 2021, 16, 231–238. [Google Scholar] [CrossRef]
- Evlakov, P.M.; Fedorova, O.A.; Grodetskaya, T.A.; Zakharova, O.V.; Gusev, A.A.; Krutyakov, Y.A.; Baranov, O.Y. Influence of Copper Oxide and Silver Nanoparticles on Microclonal Sprouts of Downy Birch (Betula pubescens Ehrh.). Nanotechnol. Russ. 2020, 15, 476–482. [Google Scholar] [CrossRef]
- Zakharova, O.V.; Gusev, A.A.; Muratov, D.S.; Shuklinov, A.V.; Strekalova, N.S.; Matveev, S.M. Titanium Trisulfide Nanoribbons Affect the Downy Birch and Poplar × Aspen Hybrid in Plant Tissue Culture via the Emission of Hydrogen Sulfide. Forests 2021, 12, 713. [Google Scholar] [CrossRef]
- Grodetskaya, T.; Fedorova, O.; Evlakov, P. Optimized method for RNA extraction from leaves of forest tree species. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 875, p. 012008. [Google Scholar] [CrossRef]
- Ritonga, F.; Ngatia, J.; Song, R.; Farooq, U.; Somadona, S.; Lestari, A.T.; Chen, S. Abiotic stresses induced physiological, biochemical, and molecular changes in Betula platyphylla: A review. Silva Fenn 2021, 55, 24. [Google Scholar] [CrossRef]
- Wen, X.; Wang, J.; Zhang, D.; Wang, Y. A Gene Regulatory Network Controlled by BpERF2 and BpMYB102 in Birch under Drought Conditions. Int. J. Mol. Sci. 2019, 20, 3071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zan, T.; Li, L.; Li, J.; Zhang, L.; Li, X. Genome-wide identification and characterization of late embryogenesis abundant protein-encoding gene family in wheat: Evolution and expression profiles during development and stress. Gene 2020, 736, 144422. [Google Scholar] [CrossRef] [PubMed]
- Moradbeygi, H.; Jamei, R.; Heidari, R.; Darvishzadeh, R. Fe2O3 nanoparticles induced biochemical responses and expression of genes involved in rosmarinic acid biosynthesis pathway in Moldavian balm under salinity stress. Physiol. Plant. 2020, 169, 555–570. [Google Scholar] [CrossRef]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Cao, X.; Ma, C.; Zhang, Z.; Zhao, N.; Ali, A.; Hou, T.; Xiang, Z.; Zhuang, J.; Wu, S.; et al. Potential Applications and Antifungal Activities of Engineered Nanomaterials against Gray Mold Disease Agent Botrytis cinerea on Rose Petals. Front. Plant Sci. 2017, 8, 1332. [Google Scholar] [CrossRef] [Green Version]
- El-Abeid, S.E.; Ahmed, Y.; Daròs, J.-A.; Mohamed, M.A. Reduced Graphene Oxide Nanosheet-Decorated Copper Oxide Nanoparticles: A Potent Antifungal Nanocomposite against Fusarium Root Rot and Wilt Diseases of Tomato and Pepper Plants. Nanomaterials 2020, 10, 1001. [Google Scholar] [CrossRef]
- Mehta, M.R.; Mahajan, H.P.; Hivrale, A.U. Green Synthesis of Chitosan Capped-Copper Nano Biocomposites: Synthesis, Characterization, and Biological Activity against Plant Pathogens. BioNanoScience 2021, 11, 417–427. [Google Scholar] [CrossRef]
- Ntasiou, P.; Kaldeli Kerou, A.; Karamanidou, T.; Vlachou, A.; Tziros, G.T.; Tsouknidas, A.; Karaoglanidis, G.S. Synthesis and Characterization of Novel Copper Nanoparticles for the Control of Leaf Spot and Anthracnose Diseases of Olive. Nanomaterials 2021, 11, 1667. [Google Scholar] [CrossRef] [PubMed]
- Studer, A.M.; Limbach, L.K.; Van Duc, L.; Krumeich, F.; Athanassiou, E.K.; Gerber, L.C.; Moch, H.; Stark, W.J. Nanoparticle cytotoxicity depends on intracellular solubility: Comparison of stabilized copper metal and degradable copper oxide nanoparticles. Toxicol. Lett. 2010, 197, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Meghana, S.; Kabra, P.; Chakraborty, S.; Padmavathy, N. Understanding the pathway of antibacterial activity of copper oxide nanoparticles. RSC Adv. 2015, 5, 12293–12299. [Google Scholar] [CrossRef]
- Karlsson, H.L.; Cronholm, P.; Hedberg, Y.; Tornberg, M.; De Battice, L.; Svedhem, S.; Wallinder, I.O. Cell membrane damage and protein interaction induced by copper containing nanoparticles—Importance of the metal release process. Toxicology 2013, 313, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Chakraborty, N.; Chatterjee, A.; Bhattacharjee, A.; Dasgupta, D.; Acharya, K. Green Synthesized Copper Oxide Nanoparticles Ameliorate Defence and Antioxidant Enzymes in Lens culinaris. Nanomaterials 2020, 10, 312. [Google Scholar] [CrossRef] [Green Version]
- Fawke, S.; Doumane, M.; Schornack, S. Oomycete interactions with plants: Infection strategies and resistance principles. Microbiol. Mol. Biol. Rev. 2015, 79, 263–280. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Liu, J.; Ren, Y.; Zhang, L.; Xue, Y.; Zhang, L.; He, J. Phytotoxicity Assessment of Copper Oxide Nanoparticles on the Germination, Early Seedling Growth, and Physiological Responses in Oryza sativa L. Bull. Environ. Contam. Toxicol. 2020, 104, 770–777. [Google Scholar] [CrossRef]
- Ramírez, V.; Agorio, A.; Coego, A.; García-Andrade, J.; Hernández, M.J.; Balaguer, B.; Ouwerkerk, P.B.F.; Zarra, I.; Vera, P. MYB46 Modulates Disease Susceptibility to Botrytis cinerea in Arabidopsis. Plant Physiol. 2011, 155, 1920–1935. [Google Scholar] [CrossRef] [Green Version]
- Mertens, J.; Habibu, A.; Cowan, D. LEA Proteins and the Evolution of the WHy Domain. Appl. Environ. Microbiol. 2018, 84, e00539-18. [Google Scholar] [CrossRef] [Green Version]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid Pathway Engineering: An Emerging Approach towards Plant Defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- Jain, D.; Khurana, J.P. Role of Pathogenesis-Related (PR) Proteins in Plant Defense Mechanism. In Molecular Aspects of Plant-Pathogen Interaction; Singh, A., Singh, I.K., Eds.; Springer: Singapore, 2018; pp. 265–281. [Google Scholar]
- Erpen, L.; Devi, H.S.; Grosser, J.W.; Dutt, M. Potential use of the DREB/ERF, MYB, NAC and WRKY transcription factors to improve abiotic and biotic stress in transgenic plants. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 132, 1–25. [Google Scholar] [CrossRef]
- Chung, I.-M.; Rekha, K.; Venkidasamy, B.; Thiruvengadam, M. Effect of Copper Oxide Nanoparticles on the Physiology, Bioactive Molecules, and Transcriptional Changes in Brassica rapa ssp. rapa Seedlings. Water Air Soil Pollut. 2019, 230, 48. [Google Scholar] [CrossRef]
- Gopalakrishnan Nair, P.M.; Chung, I.M. Biochemical, anatomical and molecular level changes in cucumber (Cucumis sativus) seedlings exposed to copper oxide nanoparticles. Biologia 2015, 70, 1575–1585. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′→3′) |
|---|---|
| DREB2 | F: AGGCAGAGAACATGGGGAAA |
| R: GAAAGTTGAGGCGAGCGTAA | |
| MYB46 | F: GATGTCGCTAGAAATGCCGG |
| R: GTTGTCTGTACGCCCTGG | |
| LEA8 | F: AATGACTTTGACATGGGCGT |
| F: AATGACTTTGACATGGGCGT | |
| PAL | F: CTGTGGCTGCAACGGTTT |
| R: TCAATTTGAGGTCCGAGCCA | |
| PR-1 | F: CCTCAAAGCCCACAATGACG |
| R: TCTCGTCCACCCATAGCTTC | |
| PR-10 | F: GGCCCGGAACCATTAAGAAG |
| R: CCACCCTCGATCAAGCTGTA | |
| GAPDH | F: CAGCCGAAGATGTCAATGCA |
| R: GGCCACTTGTTTGCTACCAA |
| Pathogen | NPs Concentration, mg/L | Level of Inhibition (%) | ||
|---|---|---|---|---|
| 3 Days | 5 Days | 7 Days | ||
| Alternaria alternata | 0.1 | 10.39 | 19.25 | 9.14 |
| 1 | 2.69 | 21.75 | 31.03 | |
| 5 | 24.23 | 31.75 | 35.69 | |
| 10 | 57.69 | 47.50 | 45.34 | |
| Fusarium oxysporum | 0.1 | 1.44 | 0 | 3.52 |
| 1 | 0 | 7.40 | 7.03 | |
| 5 | 1.44 | 20.00 | 10.81 | |
| 10 | 0 | 24.60 | 12.08 | |
| Fusarium avenaceum | 0.1 | 10.53 | 13.90 | 15.96 |
| 1 | 5.26 | 30.00 | 17.88 | |
| 5 | 21.05 | 36.59 | 19.81 | |
| 10 | 68.42 | 52.93 | 36.54 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grodetskaya, T.A.; Evlakov, P.M.; Fedorova, O.A.; Mikhin, V.I.; Zakharova, O.V.; Kolesnikov, E.A.; Evtushenko, N.A.; Gusev, A.A. Influence of Copper Oxide Nanoparticles on Gene Expression of Birch Clones In Vitro under Stress Caused by Phytopathogens. Nanomaterials 2022, 12, 864. https://doi.org/10.3390/nano12050864
Grodetskaya TA, Evlakov PM, Fedorova OA, Mikhin VI, Zakharova OV, Kolesnikov EA, Evtushenko NA, Gusev AA. Influence of Copper Oxide Nanoparticles on Gene Expression of Birch Clones In Vitro under Stress Caused by Phytopathogens. Nanomaterials. 2022; 12(5):864. https://doi.org/10.3390/nano12050864
Chicago/Turabian StyleGrodetskaya, Tatiana A., Peter M. Evlakov, Olga A. Fedorova, Vyacheslav I. Mikhin, Olga V. Zakharova, Evgeny A. Kolesnikov, Nadezhda A. Evtushenko, and Alexander A. Gusev. 2022. "Influence of Copper Oxide Nanoparticles on Gene Expression of Birch Clones In Vitro under Stress Caused by Phytopathogens" Nanomaterials 12, no. 5: 864. https://doi.org/10.3390/nano12050864
APA StyleGrodetskaya, T. A., Evlakov, P. M., Fedorova, O. A., Mikhin, V. I., Zakharova, O. V., Kolesnikov, E. A., Evtushenko, N. A., & Gusev, A. A. (2022). Influence of Copper Oxide Nanoparticles on Gene Expression of Birch Clones In Vitro under Stress Caused by Phytopathogens. Nanomaterials, 12(5), 864. https://doi.org/10.3390/nano12050864