
A Comprehensive Ecotoxicity Study of Molybdenum Disulfide Nanosheets versus Bulk form in Soil Organisms

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Species
2.2. Test Medium
2.3. Test Materials—Characterization and Quantification
2.4. Soil Spiking
2.5. Reproduction Tests
2.6. Avoidance Tests
2.7. Comet Assay Technique
2.8. Data Analysis
3. Results and Discussion
3.1. Molybdenum Disulfide Characterization and Quantification
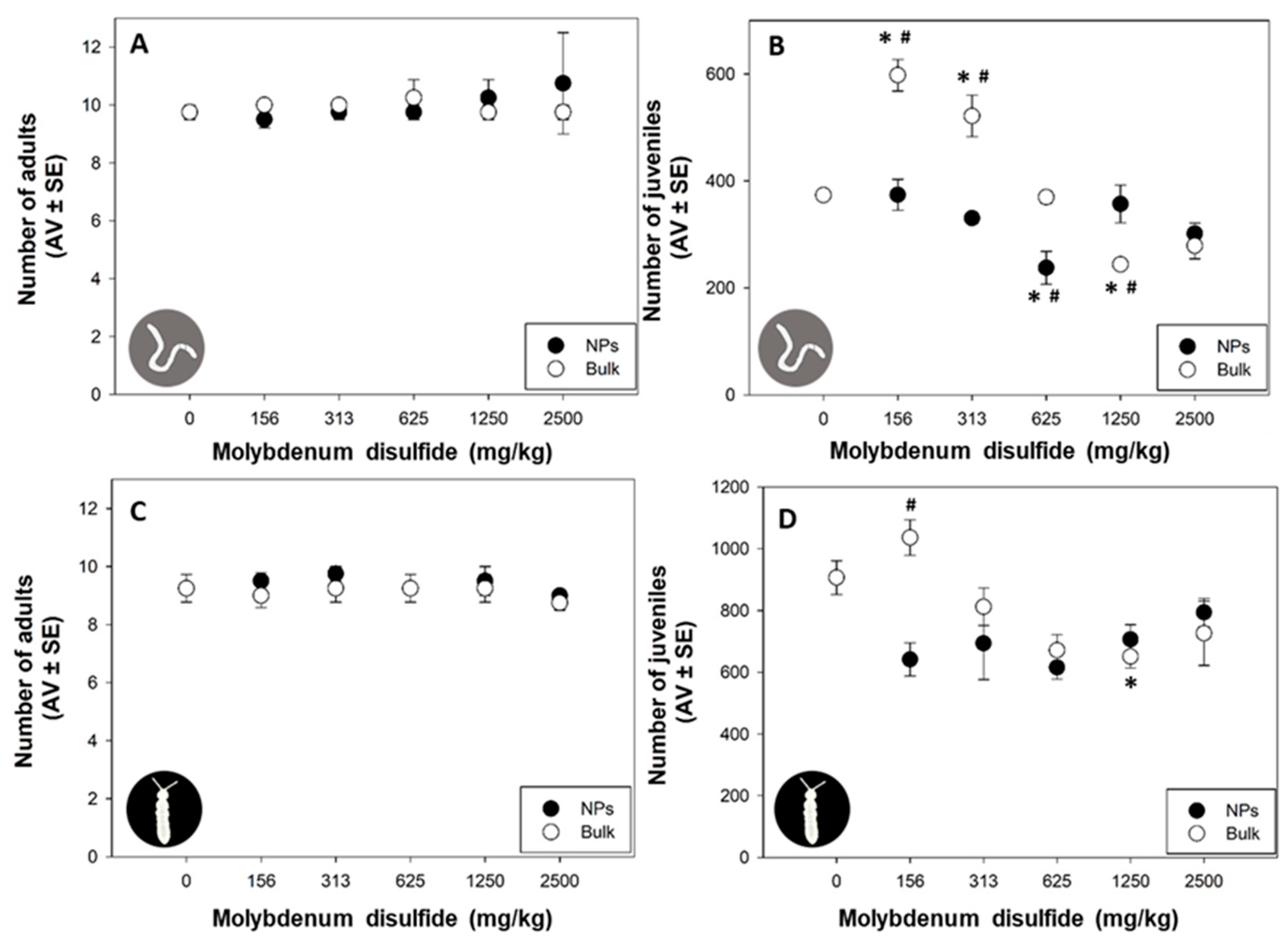
3.2. Adult Survival and Reproduction
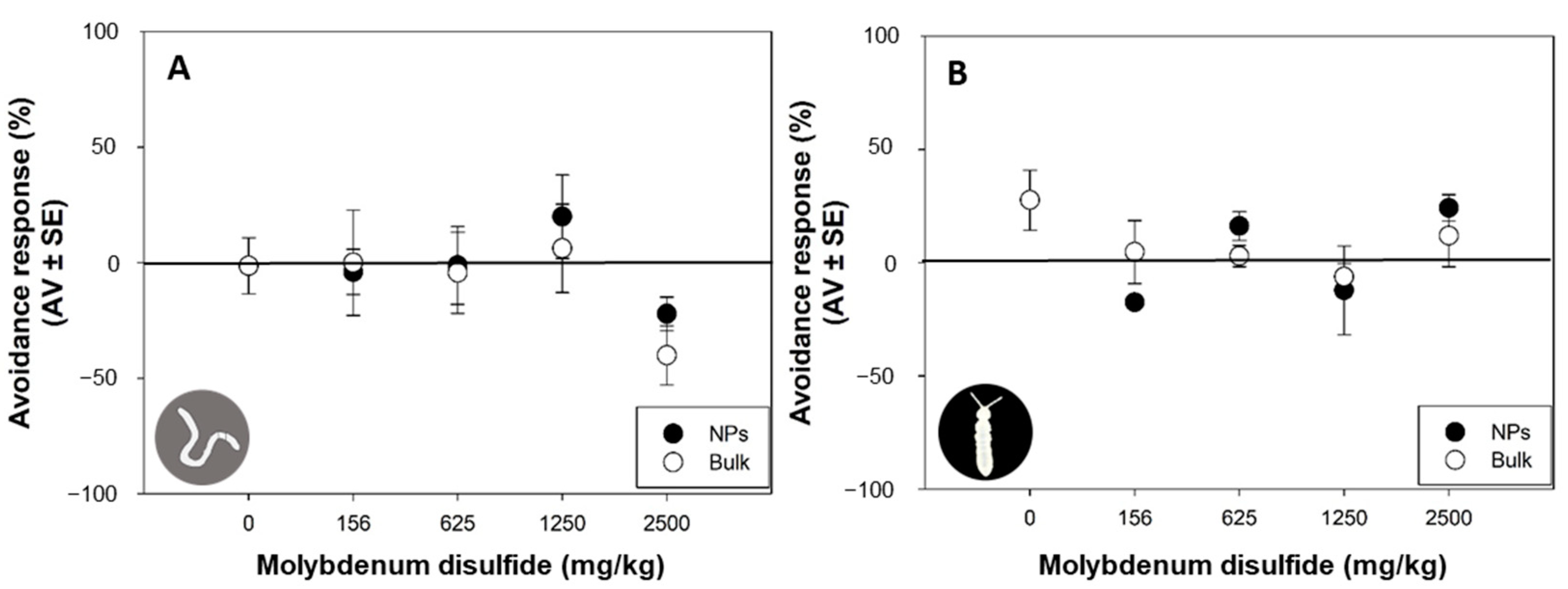
3.3. Avoidance Behavior
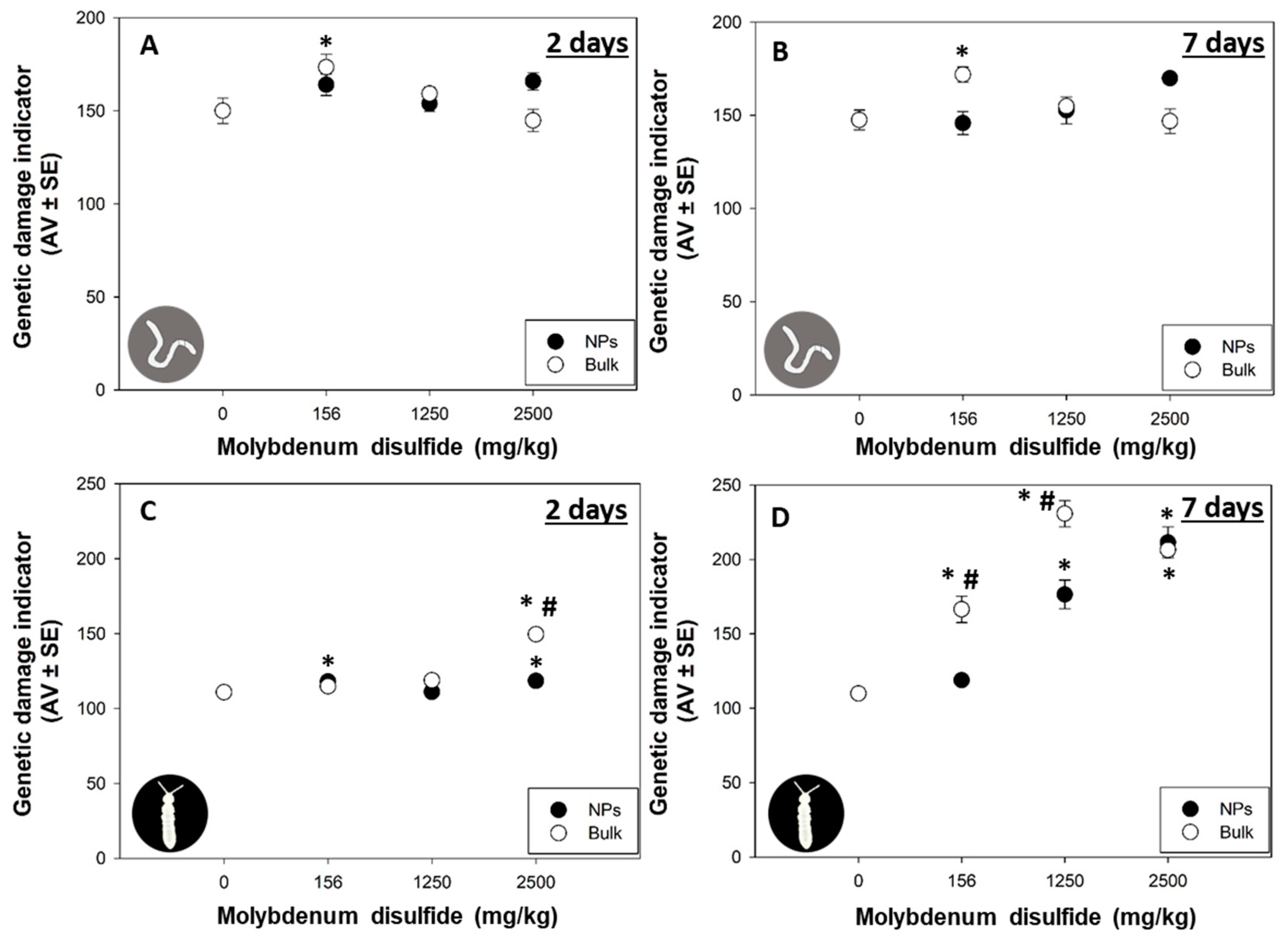
3.4. DNA Damage
3.5. Nanoparticle versus Bulk Effects—An Overview
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, N.; Xiong, G.; Liu, Z. Toxicity of metal-based nanoparticles: Challenges in the nano era. Front. Bioeng. Biotechnol. 2022, 10, 1001572. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.-W.; Alimujiang, A.; Balamurugan, S.; Zheng, J.-W.; Wang, X.; Yang, W.-D.; Cui, J.; Li, H.-Y. Physiological and molecular responses in halotolerant Dunaliella salina exposed to molybdenum disulfide nanoparticles. J. Hazard. Mater. 2021, 404, 124014. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Zhou, Q.; Zhang, X.; Hu, X. Environmental Transformations and Algal Toxicity of Single-Layer Molybdenum Disulfide Regulated by Humic Acid. Environ. Sci. Technol. 2018, 52, 2638–2648. [Google Scholar] [CrossRef] [PubMed]
- Amaral, L.O.; Daniel-da-Silva, A.L. MoS2 and MoS2 Nanocomposites for Adsorption and Photodegradation of Water Pollutants: A Review. Molecules 2022, 27, 6782. [Google Scholar] [CrossRef] [PubMed]
- Sheeja, C.; Ambali, A.; Chacko, L.; Aneesh, P.; Divya, L. MoS2 nanoparticles induce behavioral alteration and oxidative stress mediated cellular toxicity in the social insect Oecophylla smaragdina (Asian weaver ant). J. Hazard. Mater. 2020, 385, 121624. [Google Scholar] [CrossRef]
- Li, Y.; Jin, Q.; Yang, D.; Cui, J. Molybdenum Sulfide Induce Growth Enhancement Effect of Rice (Oryza sativa L.) through Regulating the Synthesis of Chlorophyll and the Expression of Aquaporin Gene. J. Agric. Food Chem. 2018, 66, 4013–4021. [Google Scholar] [CrossRef]
- Wu, B.; Chen, L.; Wu, X.; Hou, H.; Wang, Z.; Liu, S. Differential influence of molybdenum disulfide at the nanometer and micron scales in the intestinal metabolome and microbiome of mice. Environ. Sci. Nano 2019, 6, 1594–1606. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, S.; Tan, X.; Yan, X.; Zhang, W.; Huang, Y.; Ji, R.; White, J.C. Environmental implications of MoS2 nanosheets on rice and associated soil microbial communities. Chemosphere 2022, 291, 133004. [Google Scholar] [CrossRef]
- Asadi-Kavan, Z.; Khavari-Nejad, R.A.; Iranbakhsh, A.; Najafi, F. Influence of Iron Oxide Nanoparticles and Bulk (α-Fe2O3) on Metabolic and Physiological Characteristics of Oenothera biennis L. Plant. Russ. J. Plant Physiol. 2023, 70, 101. [Google Scholar] [CrossRef]
- Santos, F.C.F.; Verweij, R.A.; van Gestel, C.A.M.; Amorim, M.J.B. Toxicokinetics and toxicodynamics of Ag nanomaterials (NM300K) in the soil environment–impact on Enchytraeus crypticus (Oligochaeta). Ecotoxicol. Environ. Saf. 2023, 252, 114599. [Google Scholar] [CrossRef]
- Barreto, A.; Santos, J.; Amorim, M.J.B.; Maria, V.L. Environmental Hazards of Boron and Vanadium Nanoparticles in the Terrestrial Ecosystem—A Case Study with Enchytraeus crypticus. Nanomaterials 2021, 11, 1937. [Google Scholar] [CrossRef] [PubMed]
- Capitão, A.; Santos, J.; Barreto, A.; Amorim, M.J.B.; Maria, V.L. Single and Mixture Toxicity of Boron and Vanadium Nanoparticles in the Soil Annelid Enchytraeus crypticus: A Multi-Biomarker Approach. Nanomaterials 2022, 12, 1478. [Google Scholar] [CrossRef] [PubMed]
- Maria, V.L.; Licha, D.; Scott-Fordsmand, J.J.; Huber, C.G.; Amorim, M.J.B. Multiomics assessment in Enchytraeus crypticus exposed to Ag nanomaterials (Ag NM300K) and ions (AgNO3)—Metabolomics, proteomics (& transcriptomics). Environ. Pollut. 2021, 286, 117571. [Google Scholar] [CrossRef] [PubMed]
- Gomes, S.I.L.; Roca, C.P.; Scott-Fordsmand, J.J.; Amorim, M.J.B. High-throughput transcriptomics: Insights into the pathways involved in (nano) nickel toxicity in a key invertebrate test species. Environ. Pollut. 2019, 245, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Jośko, I.; Krasucka, P.; Skwarek, E.; Oleszczuk, P.; Sheteiwy, M. The co-occurrence of Zn-and Cu-based engineered nanoparticles in soils: The metal extractability vs. toxicity to Folsomia candida. Chemosphere 2022, 287, 132252. [Google Scholar] [CrossRef]
- Guimarães, B.; Gomes, S.I.L.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Impacts of Longer-Term Exposure to AuNPs on Two Soil Ecotoxicological Model Species. Toxics 2022, 10, 153. [Google Scholar] [CrossRef]
- Fischer, J.; Evlanova, A.; Philippe, A.; Filser, J. Soil properties can evoke toxicity of copper oxide nanoparticles towards springtails at low concentrations. Environ. Pollut. 2021, 270, 116084. [Google Scholar] [CrossRef]
- Method 3051A (SW-846); U.S. EPA. Microwave Assisted Acid Digestion of Sediments, Sludges, and Oils (Revision 1). United States Environmental Protection Agency: Washington, DC, USA, 2007.
- OECD Test 220; Enchytraeid Reproduction Test. OECD Guidelines for the Testing of Chemicals, Section 2. OECD Publication: Paris, France, 2016; pp. 1–24.
- OECD Test 232; Collembolan Reproduction Test in Soil. OECD Guidelines for the Testing of Chemicals. OECD Publication: Paris, France, 2016; pp. 1–22.
- ISO 17512-1:2008; ISO Soil Quality—Avoidance Test for Determining the Quality of Soils and Effects of Chemicals on Behaviour—Part 1: Test with Earthworms (Eisenia Fetida and Eisenia Andrei). International Organization for Standardization (ISO): Geneva, Switzerland, 2008; pp. 1–25.
- ISO 17512-2:2011; Soil Quality—Avoidance Test for Testing the Quality of Soils and Effects of Chemicals—Part 2: Test with Collembolans (Folsomia candida). Guideline. International Organization for Standardization (ISO): Geneva, Switzerland, 2011; pp. 1–13.
- Maria, V.L.; Ribeiro, M.J.; Guilherme, S.; Soares, A.M.V.M.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Silver (nano)materials cause genotoxicity in Enchytraeus crypticus, as determined by the comet assay. Environ. Toxicol. Chem. 2018, 37, 184–191. [Google Scholar] [CrossRef]
- Cardoso, D.N.; Silva, A.R.R.; Cruz, A.; Lourenço, J.; Neves, J.; Malheiro, C.; Mendo, S.; Soares, A.M.V.M.; Loureiro, S. The comet assay in Folsomia candida: A suitable approach to assess genotoxicity in collembolans. Environ. Toxicol. Chem. 2017, 36, 2514–2520. [Google Scholar] [CrossRef]
- McCourt, K.M.; Cochran, J.; Abdelbasir, S.M.; Carraway, E.R.; Tzeng, T.-R.J.; Tsyusko, O.V.; Vanegas, D.C. Potential Environmental and Health Implications from the Scaled-Up Production and Disposal of Nanomaterials Used in Biosensors. Biosensors 2022, 12, 1082. [Google Scholar] [CrossRef]
- García-Gómez, C.; Babin, M.; Obrador, A.; Álvarez, J.M.; Fernández, M.D. Toxicity of ZnO Nanoparticles, ZnO Bulk, and ZnCl2 on Earthworms in a Spiked Natural Soil and Toxicological Effects of Leachates on Aquatic Organisms. Arch. Environ. Contam. Toxicol. 2014, 67, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.C.F.; Gomes, S.I.L.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Hazard assessment of nickel nanoparticles in soil—The use of a full life cycle test with Enchytraeus crypticus. Environ. Toxicol. Chem. 2017, 36, 2934–2941. [Google Scholar] [CrossRef]
- Waalewijn-Kool, P.L.; Rupp, S.; Lofts, S.; Svendsen, C.; van Gestel, C.A.M. Effect of soil organic matter content and pH on the toxicity of ZnO nanoparticles to Folsomia candida. Ecotoxicol. Environ. Saf. 2014, 108, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.-W.; Alimujiang, A.; Cui, J.; Chen, T.-T.; Balamurugan, S.; Zheng, J.-W.; Wang, X.; Yang, W.-D.; Li, H.-Y. Molybdenum disulfide nanoparticles concurrently stimulated biomass and β-carotene accumulation in Dunaliella salina. Bioresour. Technol. 2021, 320, 124391. [Google Scholar] [CrossRef] [PubMed]
- García-Gómez, C.; García-Gutiérrez, S.; Obrador, A.; Fernández, M.D. Study of Zn availability, uptake, and effects on earthworms of zinc oxide nanoparticle versus bulk applied to two agricultural soils: Acidic and calcareous. Chemosphere 2020, 239, 124814. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; White, J.C.; He, E.; Van Gestel, C.A.M.; Qiu, H. Surface Defects Regulate the in Vivo Bioenergetic Response of Earthworm Eisenia fetida Coelomocytes to Molybdenum Disulfide Nanosheets. ACS Nano 2023, 17, 2639–2652. [Google Scholar] [CrossRef] [PubMed]
- Van Gestel, C.A.M.; Ortiz, M.D.; Borgman, E.; Verweij, R.A. The bioaccumulation of Molybdenum in the earthworm Eisenia andrei: Influence of soil properties and ageing. Chemosphere 2011, 82, 1614–1619. [Google Scholar] [CrossRef] [PubMed]
- Van Gestel, C.A.M.; McGrath, S.P.; Smolders, E.; Ortiz, M.D.; Borgman, E.; Verweij, R.A.; Buekers, J.; Oorts, K. Effect of long-term equilibration on the toxicity of molybdenum to soil organisms. Environ. Pollut. 2012, 162, 1–7. [Google Scholar] [CrossRef]
- Van Gestel, C.A.M.; Borgman, E.; Verweij, R.A.; Diez Ortiz, M. The influence of soil properties on the toxicity of molybdenum to three species of soil invertebrates. Ecotoxicol. Environ. Saf. 2011, 74, 1–9. [Google Scholar] [CrossRef]
- He, E.; Qiu, R.; Cao, X.; Song, L.; Peijnenburg, W.J.G.M.; Qiu, H. Elucidating Toxicodynamic Differences at the Molecular Scale between ZnO Nanoparticles and ZnCl2 in Enchytraeus crypticus via Nontargeted Metabolomics. Environ. Sci. Technol. 2020, 54, 3487–3498. [Google Scholar] [CrossRef]
- Barreto, A.; Santos, J.; Almeida, L.; Tavares, V.; Pinto, E.; Celeiro, M.; Garcia-Jares, C.; Maria, V.L. First approach to assess the effects of nanoplastics on the soil species Folsomia candida: A mixture design with bisphenol A and diphenhydramine. NanoImpact 2023, 29, 100450. [Google Scholar] [CrossRef] [PubMed]
- Adeel, M.; Shakoor, N.; Ahmad, M.A.; White, J.C.; Jilani, G.; Rui, Y. Bioavailability and toxicity of nanoscale/bulk rare earth oxides in soil: Physiological and ultrastructural alterations in Eisenia fetida. Environ. Sci. Nano 2021, 8, 1654–1666. [Google Scholar] [CrossRef]
- Coleman, J.G.; Johnson, D.R.; Stanley, J.K.; Bednar, A.J.; Weiss, C.A.; Boyd, R.E.; Steevens, J.A. Assessing the fate and effects of nano aluminum oxide in the terrestrial earthworm, Eisenia fetida. Environ. Toxicol. Chem. 2010, 29, 1575–1580. [Google Scholar] [CrossRef] [PubMed]
- Amorim, M.J.B.; Scott-Fordsmand, J.J. Toxicity of copper nanoparticles and CuCl2 salt to Enchytraeus albidus worms: Survival, reproduction and avoidance responses. Environ. Pollut. 2012, 164, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.M.S.; Novais, S.C.; Soares, A.M.V.M.; Amorim, M.J.B. Dimethoate affects cholinesterases in Folsomia candida and their locomotion—False negative results of an avoidance behaviour test. Sci. Total Environ. 2013, 443, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Adeel, M.; Shakoor, N.; Hussain, T.; Azeem, I.; Zhou, P.; Zhang, P.; Hao, Y.; Rinklebe, J.; Rui, Y. Bio-interaction of nano and bulk Lanthanum and Ytterbium oxides in soil system: Biochemical, genetic, and histopathological effects on Eisenia fetida. J. Hazard. Mater. 2021, 415, 125574. [Google Scholar] [CrossRef]
- Abegoda-Liyanage, C.S.; Pathiratne, A. Comparison of Toxicity of Nano and Bulk Titanium Dioxide on Nile Tilapia (Oreochromis niloticus): Acetylcholinesterase Activity Modulation and DNA Damage. Bull. Environ. Contam. Toxicol. 2023, 110, 101. [Google Scholar] [CrossRef]
- Pandit, S.; Karunakaran, S.; Boda, S.K.; Basu, B.; De, M. High Antibacterial Activity of Functionalized Chemically Exfoliated MoS2. ACS Appl. Mater. Interfaces 2016, 8, 31567–31573. [Google Scholar] [CrossRef]
- Srivastav, A.K.; Kumar, M.; Ansari, N.G.; Jain, A.K.; Shankar, J.; Arjaria, N.; Jagdale, P.; Singh, D. A comprehensive toxicity study of zinc oxide nanoparticles versus their bulk in Wistar rats. Hum. Exp. Toxicol. 2016, 35, 1286–1304. [Google Scholar] [CrossRef]
- Bae, M.; Oh, J.K.; Liu, S.; Nagabandi, N.; Yegin, Y.; DeFlorio, W.; Cisneros-Zevallos, L.; Scholar, E.M.A. Nanotoxicity of 2D Molybdenum Disulfide, MoS2, Nanosheets on Beneficial Soil Bacteria, Bacillus cereus and Pseudomonas aeruginosa. Nanomaterials 2021, 11, 1453. [Google Scholar] [CrossRef]
- Abrica-González, P.; Gómez-Arroyo, S.; Jazcilevich-Diamant, A.; Sotelo-López, A.; Flores-Márquez, A.R.; Cortés-Eslava, J. Evaluation of Toxicological Effects of ZnO and CuO Nanoparticles with Taraxacum officinale as Bioindicator. Water Air Soil Pollut. 2023, 234, 443. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nominal Concentrations (mg/kg) | Measured Concentrations (mg/kg) | |
|---|---|---|
| 2D MoS2 NPs | Bulk MoS2 | |
| 0 | 0.25 ± 0.01 | 0.25 ± 0.01 |
| 156 | 156 ± 3.4 | 149 ± 3.4 |
| 313 | 315 ± 5.6 | 306 ± 4.3 |
| 625 | 634 ± 20 | 591 ± 6.2 |
| 1250 | 1247 ± 31 | 1294 ± 34 |
| 2500 | 2505 ± 32 | 2497 ± 72 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, J.; Barreto, A.; Fernandes, C.; Silva, A.R.R.; Cardoso, D.N.; Pinto, E.; Daniel-da-Silva, A.L.; Maria, V.L. A Comprehensive Ecotoxicity Study of Molybdenum Disulfide Nanosheets versus Bulk form in Soil Organisms. Nanomaterials 2023, 13, 3163. https://doi.org/10.3390/nano13243163
Santos J, Barreto A, Fernandes C, Silva ARR, Cardoso DN, Pinto E, Daniel-da-Silva AL, Maria VL. A Comprehensive Ecotoxicity Study of Molybdenum Disulfide Nanosheets versus Bulk form in Soil Organisms. Nanomaterials. 2023; 13(24):3163. https://doi.org/10.3390/nano13243163
Chicago/Turabian StyleSantos, Joana, Angela Barreto, Cristiana Fernandes, Ana Rita R. Silva, Diogo N. Cardoso, Edgar Pinto, Ana L. Daniel-da-Silva, and Vera L. Maria. 2023. "A Comprehensive Ecotoxicity Study of Molybdenum Disulfide Nanosheets versus Bulk form in Soil Organisms" Nanomaterials 13, no. 24: 3163. https://doi.org/10.3390/nano13243163
APA StyleSantos, J., Barreto, A., Fernandes, C., Silva, A. R. R., Cardoso, D. N., Pinto, E., Daniel-da-Silva, A. L., & Maria, V. L. (2023). A Comprehensive Ecotoxicity Study of Molybdenum Disulfide Nanosheets versus Bulk form in Soil Organisms. Nanomaterials, 13(24), 3163. https://doi.org/10.3390/nano13243163