
Biomass Valorization: Sustainable Methods for the Production of Hemicellulolytic Catalysts from Thermoanaerobacterium thermostercoris strain BUFF

 ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Waste Biomass
2.2. Thermoanaerobacterium thermostercoris Culture Conditions
2.3. Partial Enzyme Purification and Protein Content Determination
2.4. Enzymatic Assays
2.5. Biochemical Characterization of Enzyme Activities
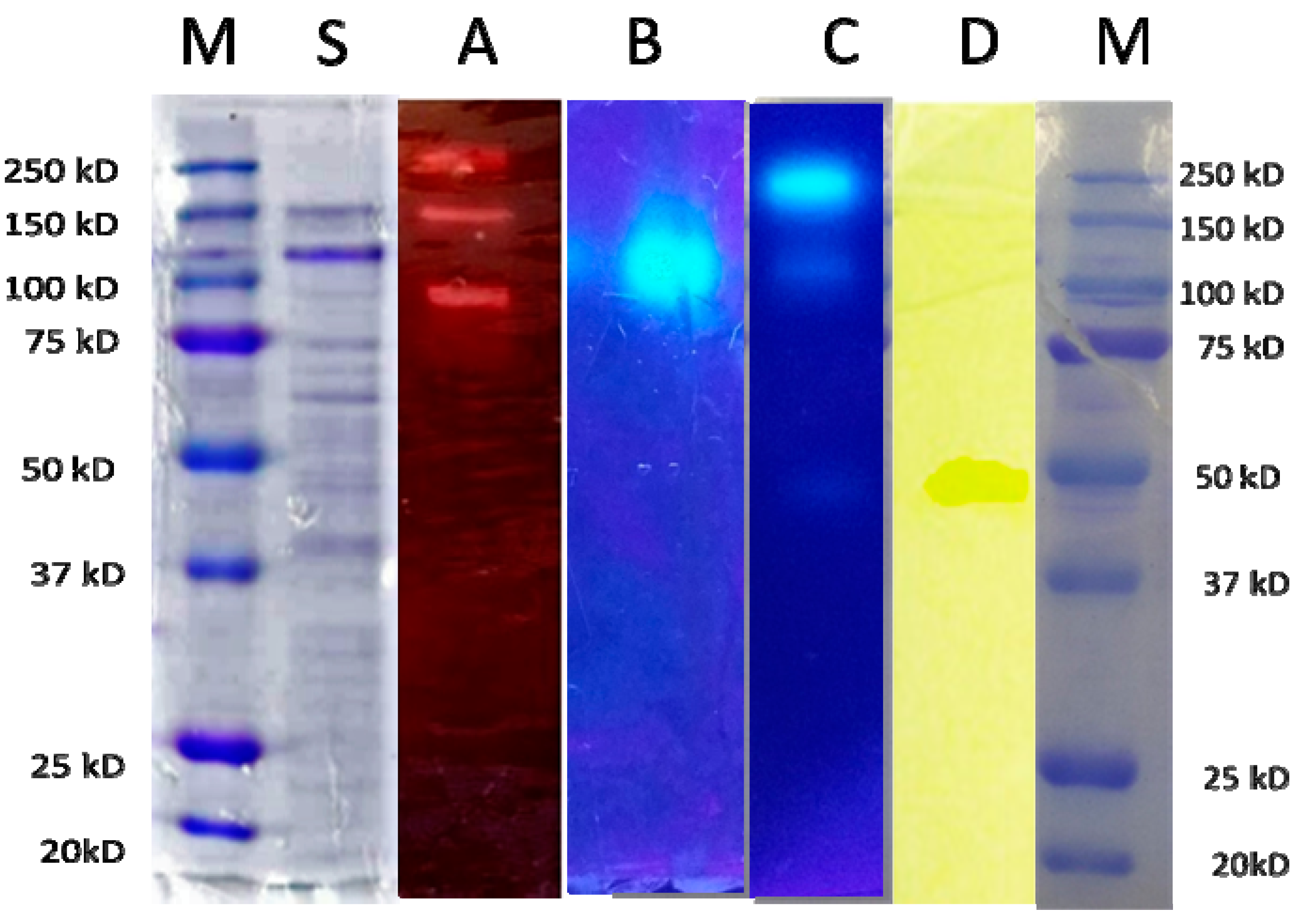
2.6. Electrophoresis and Zymogram
2.7. Ethanol and Hydrogen Production by T. thermostercoris
2.8. Enzymatic Degradation of Hemicellulolytic Extracts from Cynara Cardunculus and Arundo Donax Biomass
3. Results and Discussion
3.1. T. thermostercoris Growth Test on Different Substrates for Hydrogen and Ethanol Production
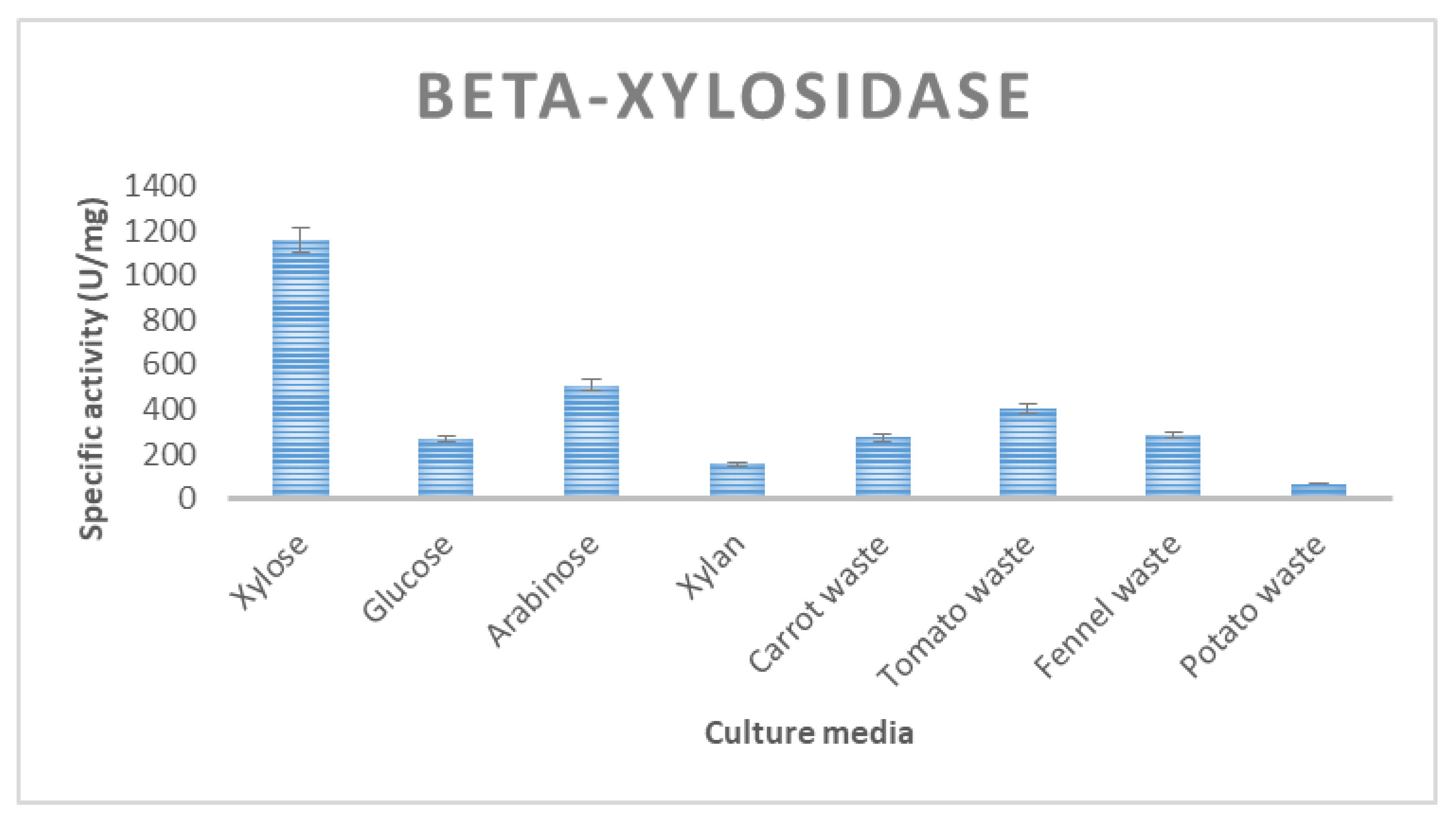
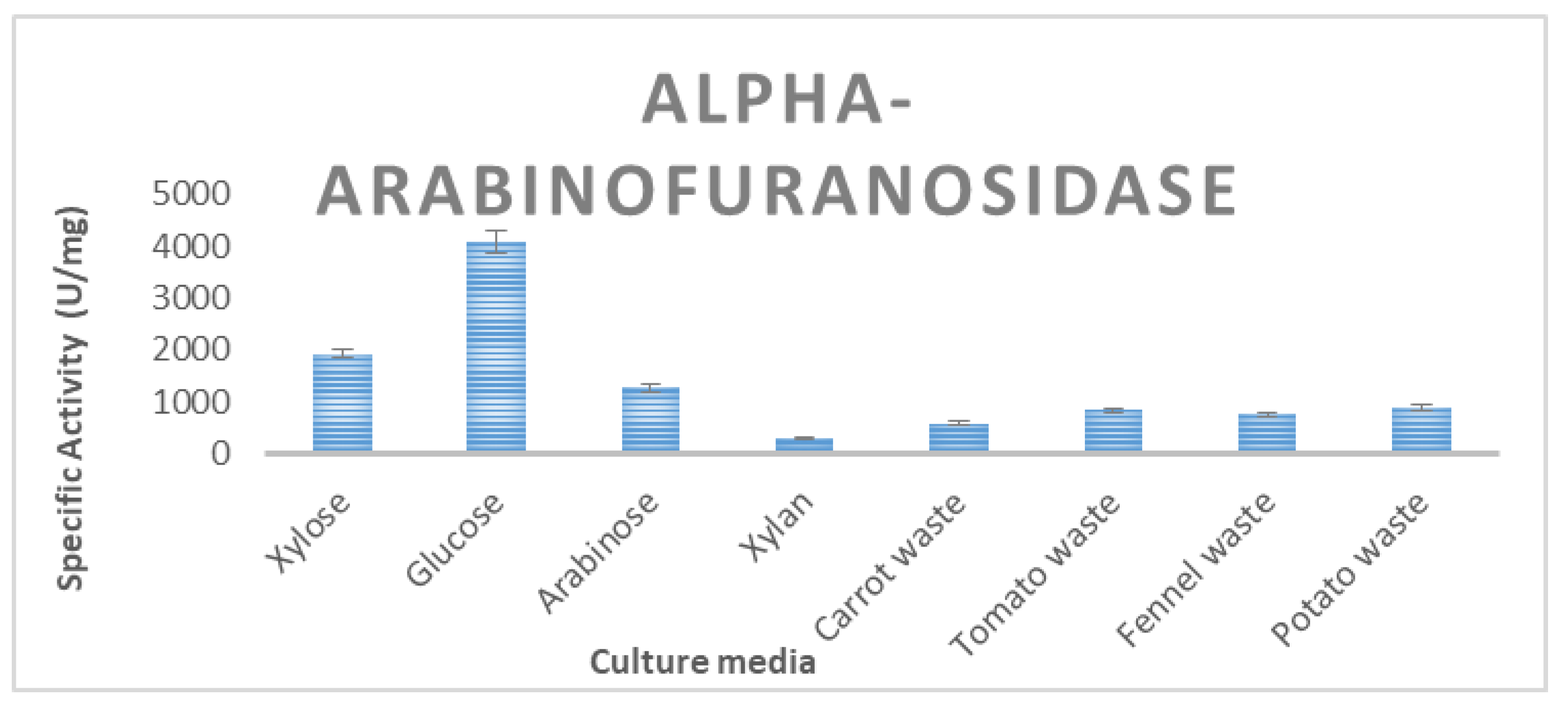
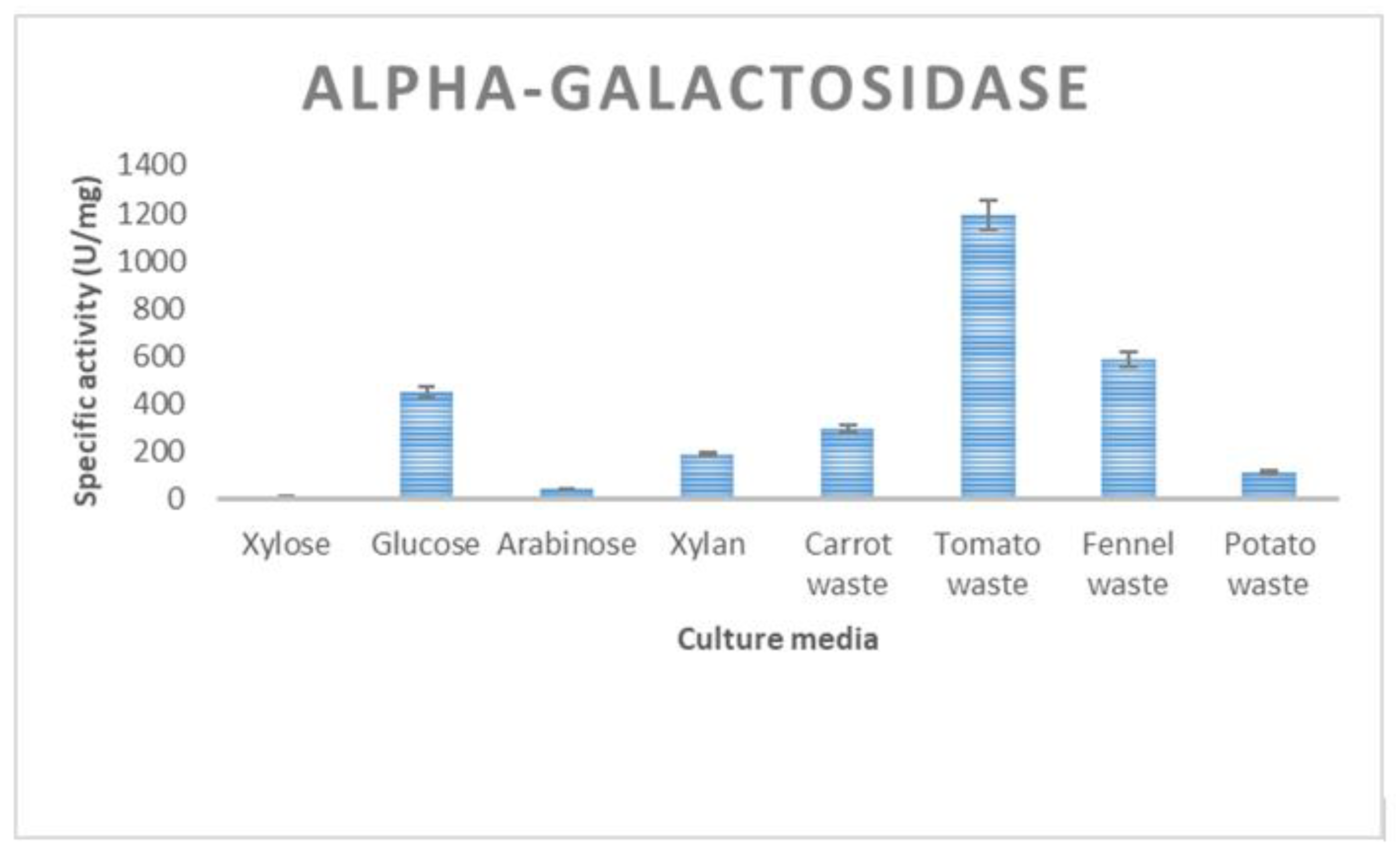
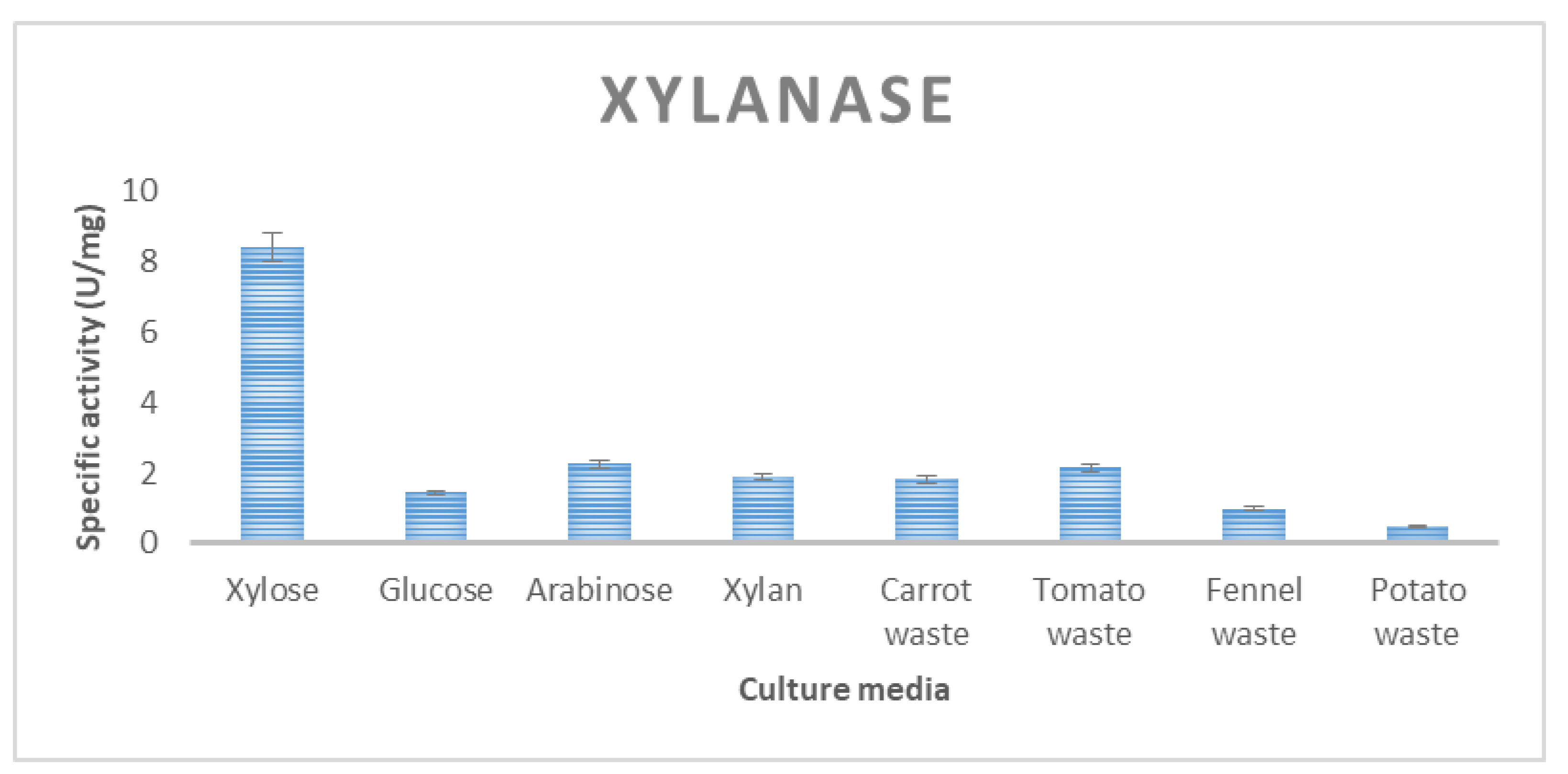
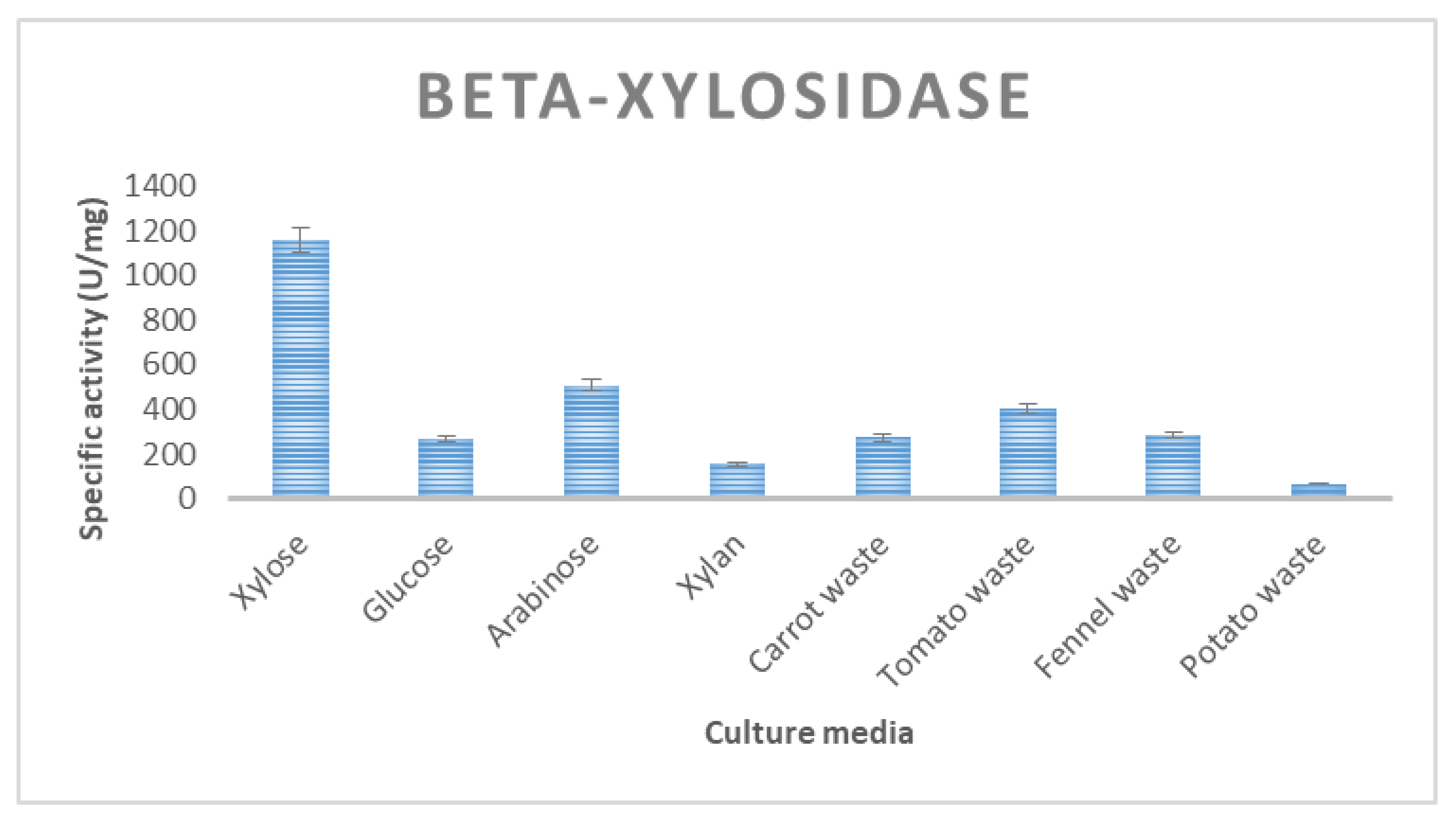
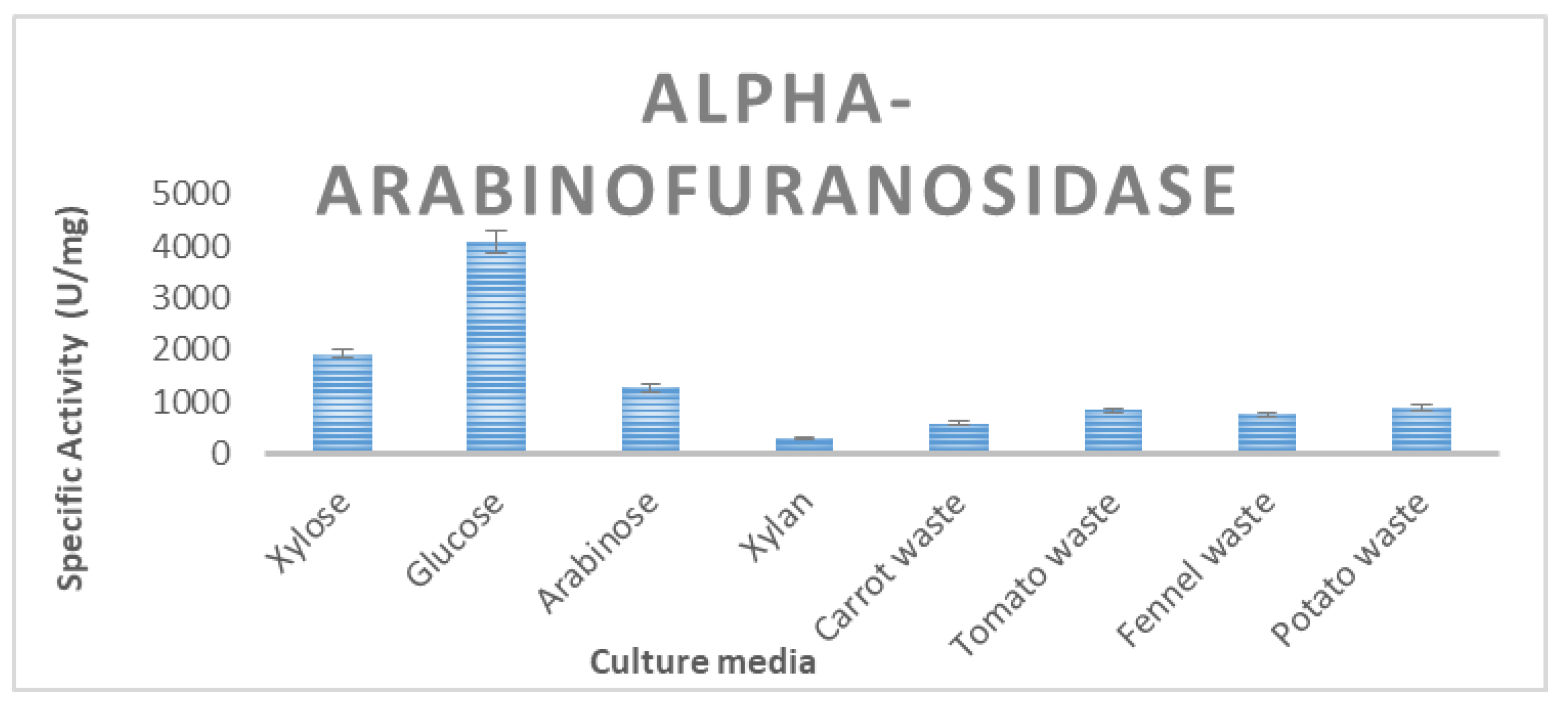
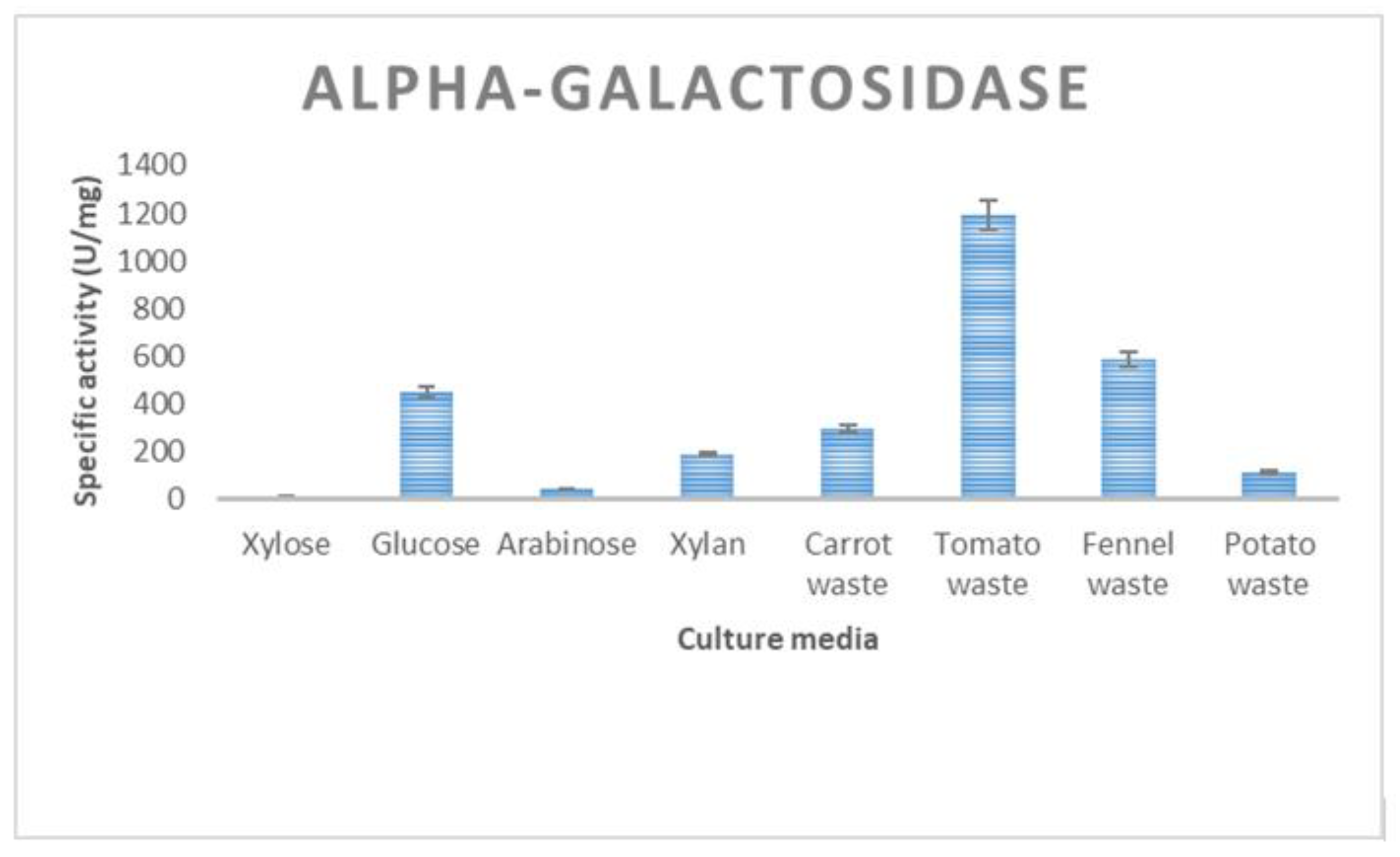
3.2. Enzyme Production of T. thermostercoris in Selected Growth Media
3.3. Bioconversion of Hemicellulose Fractions from Arundo Donax and Cynara Cardunculus
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maina, S.; Kachrimanidou, V.; Koutinas, A. A roadmap towards a circular and sustainable bioeconomy through waste valorization. Curr. Opin. Green Sustain. Chem. 2017, 8, 18–23. [Google Scholar] [CrossRef]
- Caldeira, C.; Vlysidis, A.; Fiore, G.; De Laurentiis, V.; Vignali, G.; Sala, S. Sustainability of food waste biorefinery: A review on valorisation pathways, techno-economic constraints, and environmental assessment. Bioresour. Technol. 2020, 312, 123575. [Google Scholar] [CrossRef] [PubMed]
- Cristóbal, J.; Caldeira, C.; Corrado, S.; Sala, S. Techno-economic and profitability analysis of food waste biorefineries at European level. Bioresour. Technol. 2018, 259, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Liguori, R.; Faraco, V. Biological processes for advancing lignocellulosic waste biorefinery by advocating circular economy. Bioresour. Technol. 2016, 215, 13–20. [Google Scholar] [CrossRef]
- Kamm, B. Biorefineries–their scenarios and challenges. Pure. Appl. Chem. 2014, 86, 821–831. [Google Scholar] [CrossRef]
- Kamm, B.; Kamm, M. Principles of Biorefineries Mini-Review. Appl. Microbiol. Biotechnol. 2004, 64, 137–145. [Google Scholar] [CrossRef]
- Xiong, X.; Yu, I.K.M.; Tsang, D.C.W.; Bolan, N.S.; Sik, O.Y.; Igalavithana, A.D.; Kirkham, M.B.; Kim, K.H.; Vikrant, K. Value-added chemicals from food supply chain wastes: State-of-the-art review and future prospects. Chem. Eng. Sci. 2019, 375, 121983. [Google Scholar] [CrossRef]
- Di Donato, P.; Finore, I.; Poli, A.; Nicolaus, B.; Lama, L. The production of second generation bioethanol: The biotechnology potential of thermophilic bacteria. J. Clean. Prod. 2019, 233, 1410–1417. [Google Scholar] [CrossRef]
- Di Donato, P.; Buono, A.; Poli, A.; Finore, I.; Abbamondi, G.R.; Nicolaus, B.; Lama, L. Exploring Marine Environments for the Identification of Extremophiles and Their Enzymes for Sustainable and Green Bioprocesses. Sustainability 2019, 11, 149. [Google Scholar] [CrossRef] [Green Version]
- Di Donato, P.; Taurisano, V.; Poli, A.; Gomez d’Ayala, G.; Nicolaus, B.; Malinconico, M.; Santagata, G. Vegetable wastes derived polysaccharides as natural eco-friendly plasticizers of sodium alginate. Carbohydr. Polym. 2020, 229, 115427. [Google Scholar] [CrossRef]
- Cimini, D.; Argenzio, O.; D’Ambrosio, S.; Lama, L.; Finore, I.; Finamore, R.; Pepe, O.; Faraco, V.; Schiraldi, C. Production of succinic acid from Basfia succiniciproducens up to the pilot scale from Arundo donax hydrolysate. Bioresour. Technol. 2016, 222, 355–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagliara, V.; Nasso, R.; Di Donato, P.; Finore, I.; Poli, A.; Masullo, M.; Arcone, R. Lemon peel polyphenol extract reduces interleukin-6-induced cell migration, invasiveness, and matrix metalloproteinase-9/2 expression in human gastric adenocarcinoma mkn-28 and ags cell lines. Biomolecules 2019, 9, 833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finore, I.; Poli, A.; Di Donato, P.; Lama, L.; Trincone, A.; Fagnano, M.; Mori, M.; Nicolaus, B.; Tramice, A. The hemicellulose extract from Cynara cardunculus: A source of value-added biomolecules produced by xylanolytic thermozymes. Green Chem. 2016, 18, 2460–2472. [Google Scholar] [CrossRef]
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial Diversity in Extreme Marine Habitats and Their Biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbieta, M.S.; Donati, E.R.; Chan, K.G.; Shahar, S.; Sin, L.L.; Goh, K.M. Thermophiles in the genomic era: Biodiversity, science, and applications. Biotechnol. Adv. 2015, 33, 633–647. [Google Scholar] [CrossRef]
- Bala, A.; Singh, B. Cellulolytic and xylanolytic enzymes of thermophiles for the production of renewable biofuels. Renew. Energy 2019, 136, 1231–1244. [Google Scholar] [CrossRef]
- Kumar, S.; Dangi, A.K.; Shukla, P.; Baishya, D.; Khare, S.K. Thermozymes: Adaptive strategies and tools for their biotechnological applications. Bioresour. Technol. 2019, 278, 372–382. [Google Scholar] [CrossRef]
- Singh, V.; Singh, M.P.; Verma, V.; Singh, P.; Srivastava, R.; Singh, A.K. Characteristics of Cold Adapted Enzyme and Its Comparison with Mesophilic and Thermophilic Counterpart. Cell Mol. Biol. 2016, 62. [Google Scholar] [CrossRef]
- Di Donato, P.; Fiorentino, G.; Anzelmo, G.; Tommonaro, G.; Nicolaus, B.; Poli, A. Re-Use of Vegetable Wastes as Cheap Substrates for Extremophile Biomass Production. Waste Biomass. Valor. 2011, 2, 103–111. [Google Scholar] [CrossRef]
- Barreca, F. Use of giant reed Arundo Donax L. in rural constructions Agricultural. Agric. Eng. Int. CIGR J. 2012, 14, 46–52. [Google Scholar]
- Fernández, J.; Curt, M.D.; Aguado, P.L. Industrial applications of Cynara cardunculus L. for energy and other uses. Ind. Crops. Prod. 2006, 24, 222–229. [Google Scholar] [CrossRef]
- Romano, I.; Di Pasquale, L.; Orlando, P.; Lama, L.; d’Ippolito, G.; Pascual, J.; Gambacorta, A. Thermoanaerobacterium thermostercus sp. nov., a new anaerobic thermophilic hydrogen producing bacterium from buffalo-dung. Extremophiles 2010, 14, 233–240. [Google Scholar] [CrossRef]
- Romano, I.; Di Pasquale, L.; Orlando, P.; Lama, L.; d’Ippolito, G.; Pascual, J.; Gambacorta, A. Thermoanaerobacterium thermostercus corrig. Thermoanaerobacterium thermostercoris. List of new names and new combinations previously effectively, but not validly, published. Int. J. Syst. Evol. Microbiol. 2011, 61, 2563–2565. [Google Scholar] [CrossRef] [Green Version]
- Wolin, E.A.; Wolin, M.J.; Wolfe, R.S. Formation of methane by bacterial extract. J. Biol. Chem. 1963, 238, 2882–2886. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bernfeld, P. Amylases α and β. Methods Enzymol. 1955, 1, 149e58. [Google Scholar] [CrossRef]
- Lama, L.; Calandrelli, V.; Gambacorta, A.; Nicolaus, B. Purification and characterization of thermostable xylanase and β-xylosidase by the thermophilic bacterium Bacillus thermantarcticus. Res. Microbiol. 2004, 155, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Finore, I.; Vigneron, A.; Vincent, W.F.; Leone, L.; Di Donato, P.; Schiano Moriello, A.; Nicolaus, B.; Poli, A. Novel psychrophiles and exopolymers from permafrost thaw lake sediments. Microorganisms 2020, 8, 1282. [Google Scholar] [CrossRef]
- Valkova, N.; Le´pine, F.; Labrie, L.; Dpont, M.; Beaudet, R. Purification and Characterization of PrbA, a New Esterase from Enterobacter cloacae Hydrolyzing the Esters of 4-Hydroxybenzoic Acid (Parabens). J. Biol. Chem. 2003, 278, 12779–12785. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Martínez, T.F.; Alarcón, F.J.; Díaz-López, M.; Moyano, F.J. Improved detection of amylase activity by sodium dodecyl sulphate-polyacrylamide gel electrophoresis with copolymerized starch. Electrophoresis 2000, 21, 2940–2943. [Google Scholar] [CrossRef]
- Lama, L.; Tramice, A.; Finore, I.; Anzelmo, G.; Calandrelli, V.; Pagnotta, E.; Tommonaro, G.; Poli, A.; Di Donato, P.; Nicolaus, B.; et al. Degradative actions of microbial xylanolytic activities on hemicelluloses from rhizome of Arundo donax. AMB Express. 2014, 4, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, C.D. The microbial cell factory. Organic. Biomol. Chem. 2012, 14, 1949–1957. [Google Scholar] [CrossRef]
- Bibra, M.; Rathinam, N.K.; Johnson, G.R.; Sani, R.K. Single pot biovalorization of food waste to ethanol by Geobacillus and Thermoanaerobacter spp. Renew. Energy 2020, 155, 1032–1041. [Google Scholar] [CrossRef]
- Nawirska, A.; Kwasniewska, M. Dietary fibre fractions from fruit and vegetable processing waste. Food Chem. 2005, 91, 221–225. [Google Scholar] [CrossRef]
- Sanchez-Vazquez, S.A.; Hailes, H.C.; Evans, J.R.G. Hydrophobic Polymers from Food Waste: Resources and Synthesis. Polym. Rev. 2013, 53, 627–694. [Google Scholar] [CrossRef]
- Tommonaro, G.; Poli, A.; De Rosa, S.; Nicolaus, B. Tomato derived polysaccharides for biotechnological applications: Chemical and biological approaches. Molecules 2008, 13, 1384–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-E.; Jain, M.K.; Lee, C.; Zeikus, J.G. Taxonomic distinction of saccharolytic thermophilic anaerobes: Description of Thermoanaerobacterium xylanolyticum gen. nov., sp. nov., and Thermoanaerobacterium saccharolyticum gen. nov., sp. nov.; reclassification of Thermoanaerobium brockii, Clostridium thermosulfurogenes, and Clostridium thermohydrosulfiricum ElO0-69 as Thermoanaerobacter brockii comb. nov., Thermoanaerobacterium thermosulfurigenes comb. nov., and Thermoanaerobacter thermohydrosulfuricus comb. nov., respectively; and transfer of Clostridium thermohydrosulfuricum 39E to Thermoanaerobacter ethanolicus. Int. J. Syst. Evol. Microbiol. 1993, 43, 41–51. [Google Scholar]
- Shao, W.; Deblois, S.; Wiegel, J. A high molecular weight, cell-associated xylanase isolated from exponentially growing Thermoanaerobacterium sp. strain JW/SL-YS485. Appl. Environ. Microbiol. 1995, 61, 937–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chimtong, S.; Tachaapaikoon, C.; Pason, P.; Kyu, K.L.; Kosugi, A.; Mori, Y.; Ratanakhanokchai, K. Isolation and characterization of endocellulase-free multienzyme complex from newly isolated Thermoanaerobacterium thermosaccharolyticum strain NOI-1. J. Microbiol. Biotechnol. 2011, 21, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Harnvoravongchai, P.; Singwisut, R.; Ounjai, P.; Aroonnual, A.; Kosiyachinda, P.; Janvilisri, T.; Chankhamhaengdecha, S. Isolation and characterization of thermophilic cellulose and hemicellulose degrading bacterium, Thermoanaerobacterium sp. R63 from tropical dry deciduous forest soil. PLoS ONE 2020, 15, e0236518. [Google Scholar] [CrossRef]
- Wongwilaiwalin, S.; Rattanachomsri, U.; Laothanachareon, T.; Eurwilaichitr, L.; Igarashi, Y.; Champreda, V. Analysis of a thermophilic lignocellulose degrading microbial consortium and multi-species lignocellulolytic enzyme system. Enzyme Microb Technol. 2010, 47, 283–290. [Google Scholar] [CrossRef]
- Bhalla, A.; Bischoff, K.M.; Sani, R.K. Highly thermostable GH39 β-xylosidase from a Geobacillus sp. strain WSUCF1. BMC Biotechnol. 2014, 14, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, W.; Xue, Y.; Wu, A.; Kataeva, I.; Pei, J.; Wu, H.; Wiegel, J. Characterization of a novel beta-xylosidase, XylC, from Thermoanaerobacterium saccharolyticum JW/SL-YS485. Appl. Environ. Microbiol. 2011, 77, 719–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Medium | O.D. λ540 nm | H2 (mL/L/h) | CH2CH2OH (g/L) |
|---|---|---|---|
| TN + xylose | 0.707 ± 0.021 | 20 ± 0.02 | 0.1331 ± 0.08 |
| TN + glucose | 0.853 ± 0.016 | 24.7 ± 0.02 | 0.1433 ± 0.02 |
| TN + arabinose | 0.637 ± 0.013 | 14.8 ± 0.022 | 0.1142 ± 0.01 |
| TN + xylan | 0.980 ± 0.024 | 27 ± 0.05 | 0.1565 ± 0.01 |
| TN + carrot waste | 1.182 ± 0.011 | 31 ± 0.03 | 0.2401 ± 0.02 |
| TN + tomato waste | 0.964 ± 0.029 | 5.3 ± 0.01 | 0.0073 ± 0.0021 |
| TN + fennel waste | 1.154 ± 0.021 | 35 ± 0.05 | 0.1292 ± 0.01 |
| TN + potato waste | 0.475 ± 0.011 | 0.16 ± 0.02 | n.d. |
| Enzyme Activity (U/mg) | Glucose | Carrot | Fennel | Potato | Tomato |
|---|---|---|---|---|---|
| Xylanase | 1.4 ± 0.01 | 1.8 ± 0.1 | 0.9 ± 0.01 | 0.5 ± 0.08 | 2.14 ± 0.8 |
| β-xylosidase | 266 ± 21 | 274 ± 15 | 284 ± 13 | 115 ± 7.2 | 405 ± 15 |
| α-arabinofuranosidase | 4092 ± 34 | 587 ± 13 | 754 ± 8.5 | 881 ± 21 | 831 ± 18 |
| Acetyl-xylan esterase | 4302 ± 28 | 2546 ± 13 | 6019 ± 34 | 5411 ± 91 | 0 |
| α-galactosidase | 485 ± 19 | 335 ± 12 | 588 ± 9 | 168 ± 6 | 1194 ± 31 |
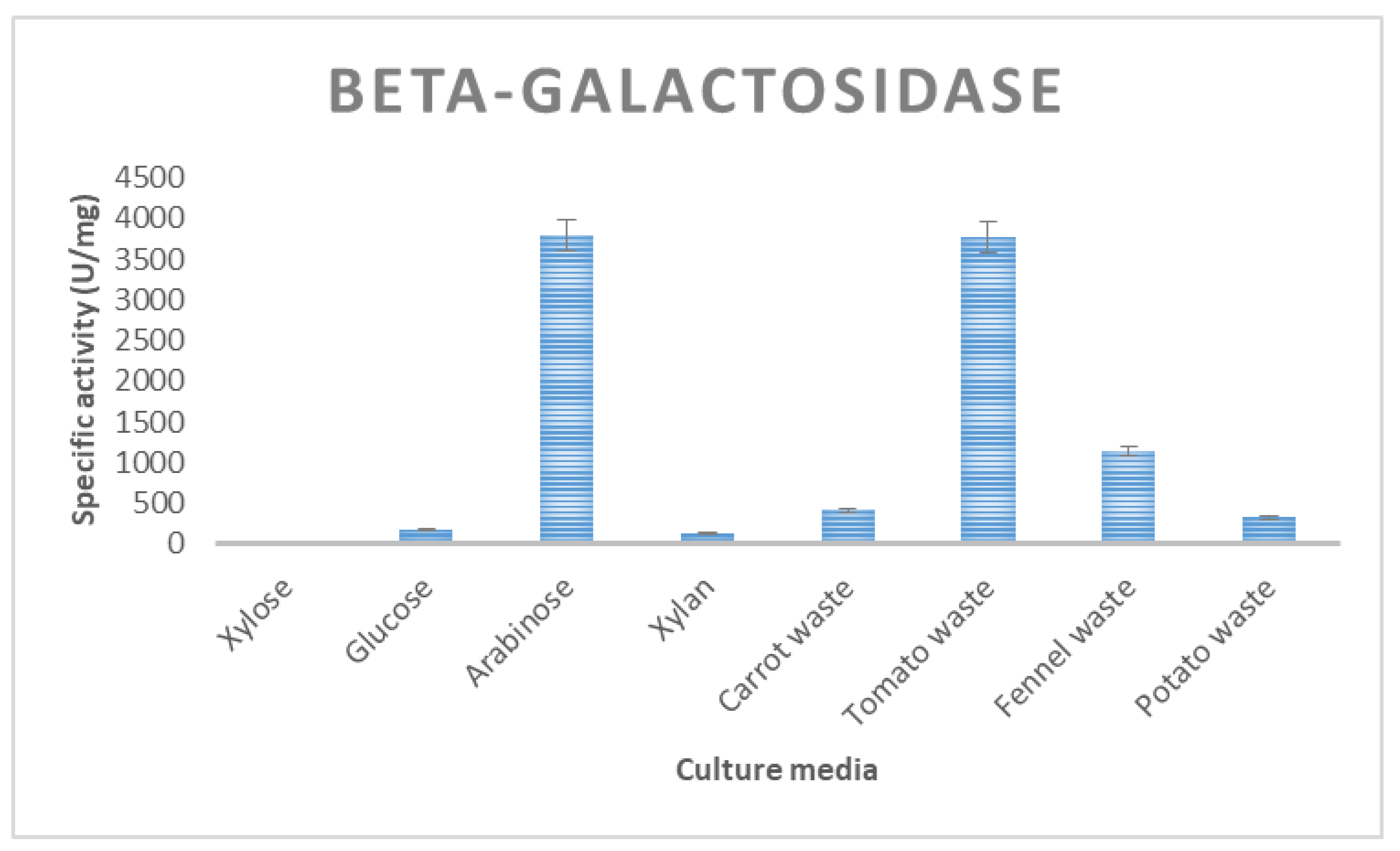
| β-galactosidase | 167 ± 11 | 417 ± 18 | 1139 ± 15 | 322 ± 7 | 3773 ± 43 |
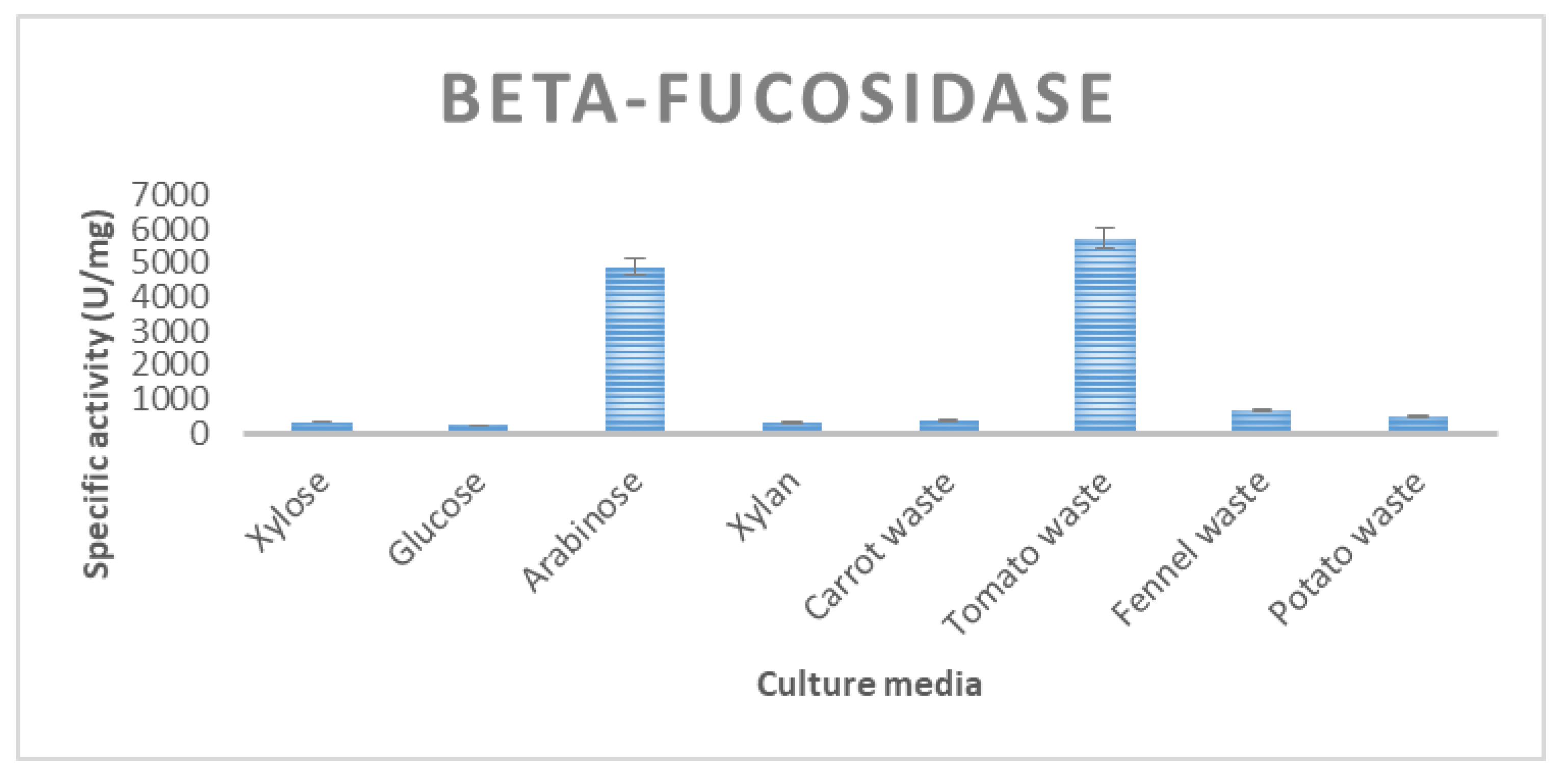
| β-fucosidase | 216 ± 7.6 | 519 ± 23 | 1018 ± 19 | 476 ± 9 | 5725 ± 47 |
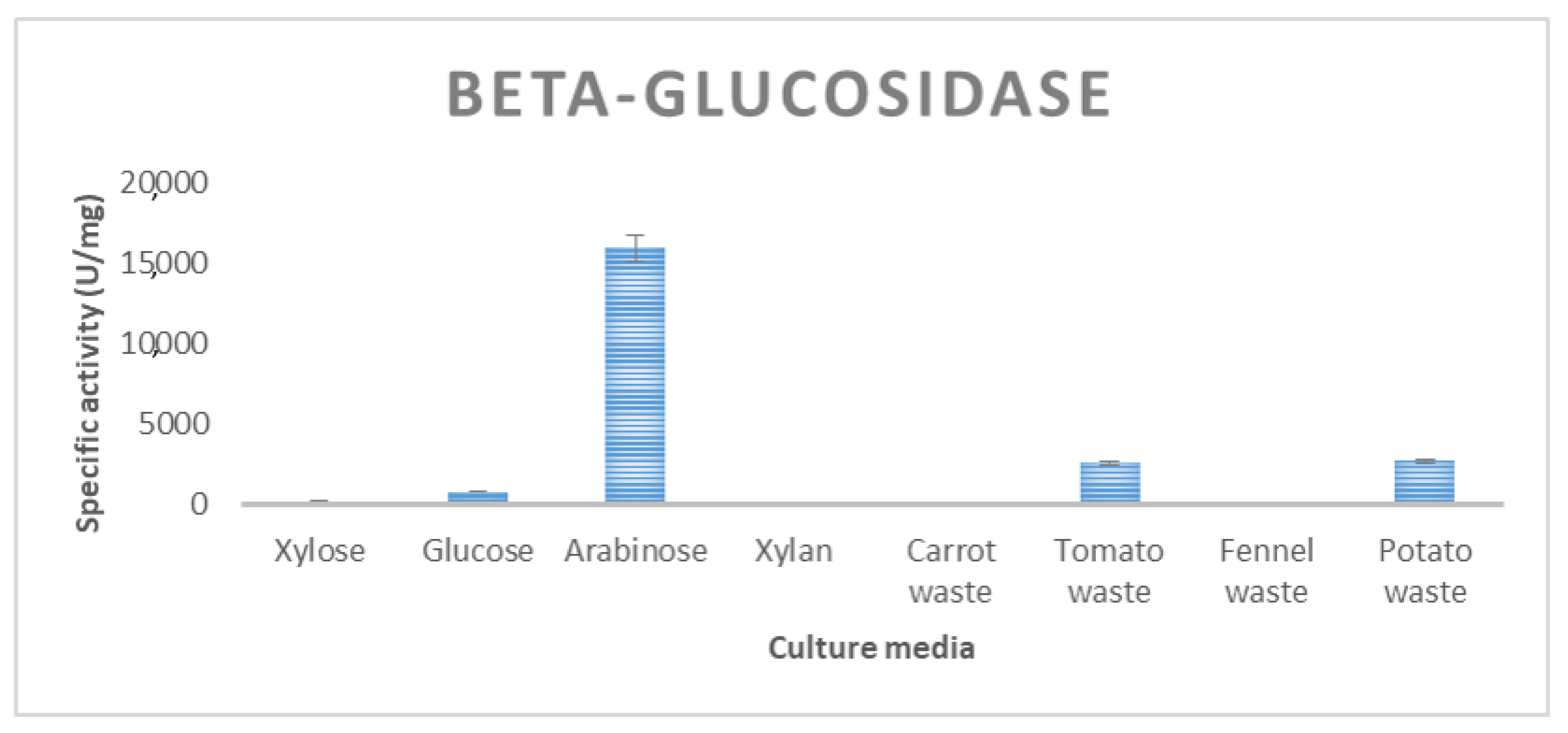
| β-glucosidase | 827 ± 15 | 347 ± 7 | 476 ± 12 | 2714 ± 40 | 2576 ± 33 |
| Enzymes | Optimum Temperature (Range) | Optimum pH (Range) | Molecular Weight (kDa) |
|---|---|---|---|
| Xylanase | 80 °C (60–85 °C) | 6.0 (3.5–6.2) | 250, 150, 100 |
| β-xylosidase | 70 °C (50–80 °C) | 5.6 (4.0–7.0) | 120 |
| α-arabinofuranosidase | 70 °C (60–90 °C) | 5.6 (4.0–9.0) | 250 |
| Acetyl-xylan esterase | 70 °C (60–90 °C) | 3.5 (3.0–6.0) | 50 |
| β-glucosidase | 60 °C (35–75 °C) | 6.5 (5.0–8.5) | n.d. |
| Enzymes | Temperature (°C) | Pre-Incubation Time (min) | Residual Activity (%) | |
|---|---|---|---|---|
| Xylanase | 75 | 240 | 54 * | 50 ** |
| β-xylosidase | 70 | 60 | 47 * | 51 ** |
| α-arabinofuranosidase | 80 | 60 | 45 * | 50 ** |
| Acetyl-xylanesterase | 80 | 10 | 47 * | 44 ** |
| Substrate | RS (Tf) (mg) | RS (%, w/w) |
|---|---|---|
| CK | 5.2 ± 0.016 | 65 |
| AK | 3.84 ± 0.012 | 48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Finore, I.; Romano, I.; Leone, L.; Di Donato, P.; Nicolaus, B.; Poli, A.; Lama, L. Biomass Valorization: Sustainable Methods for the Production of Hemicellulolytic Catalysts from Thermoanaerobacterium thermostercoris strain BUFF. Resources 2021, 10, 115. https://doi.org/10.3390/resources10110115
Finore I, Romano I, Leone L, Di Donato P, Nicolaus B, Poli A, Lama L. Biomass Valorization: Sustainable Methods for the Production of Hemicellulolytic Catalysts from Thermoanaerobacterium thermostercoris strain BUFF. Resources. 2021; 10(11):115. https://doi.org/10.3390/resources10110115
Chicago/Turabian StyleFinore, Ilaria, Ida Romano, Luigi Leone, Paola Di Donato, Barbara Nicolaus, Annarita Poli, and Licia Lama. 2021. "Biomass Valorization: Sustainable Methods for the Production of Hemicellulolytic Catalysts from Thermoanaerobacterium thermostercoris strain BUFF" Resources 10, no. 11: 115. https://doi.org/10.3390/resources10110115
APA StyleFinore, I., Romano, I., Leone, L., Di Donato, P., Nicolaus, B., Poli, A., & Lama, L. (2021). Biomass Valorization: Sustainable Methods for the Production of Hemicellulolytic Catalysts from Thermoanaerobacterium thermostercoris strain BUFF. Resources, 10(11), 115. https://doi.org/10.3390/resources10110115