
Indole-3 Carbinol and Diindolylmethane Mitigated β-Amyloid-Induced Neurotoxicity and Acetylcholinesterase Enzyme Activity: In Silico, In Vitro, and Network Pharmacology Study

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. In Silico Docking
2.3. Molecular Dynamics
2.4. Network Pharmacology Studies
2.5. In Vitro Studies
2.5.1. Cholinesterase Inhibition Assay
2.5.2. Amyloid β Inhibition Assay
2.6. Cell Culture Studies
2.7. Measurement of Oxidative Stress
2.8. Statistical Analysis
3. Results
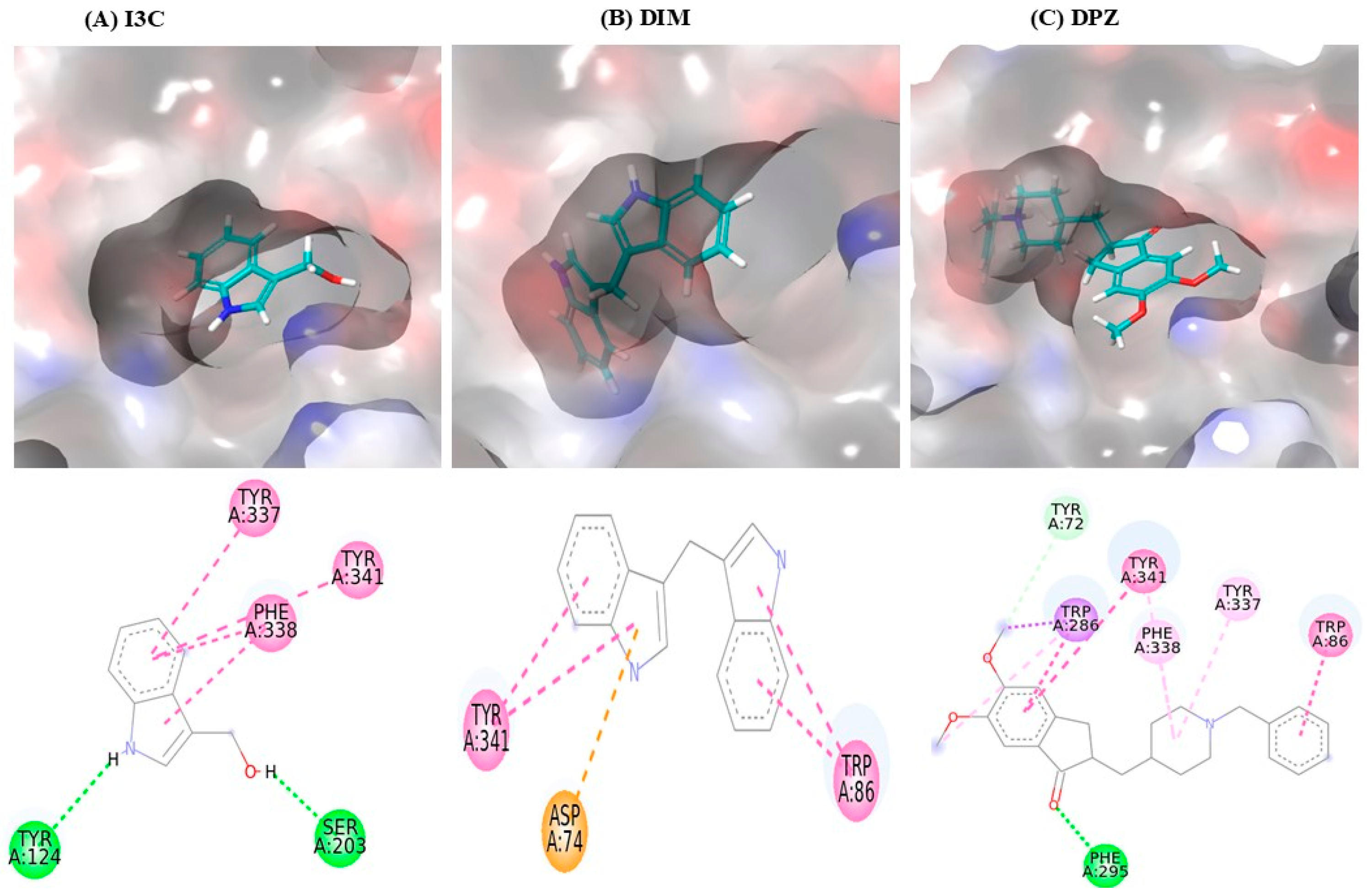
3.1. I3C and DIM Bind to AchE
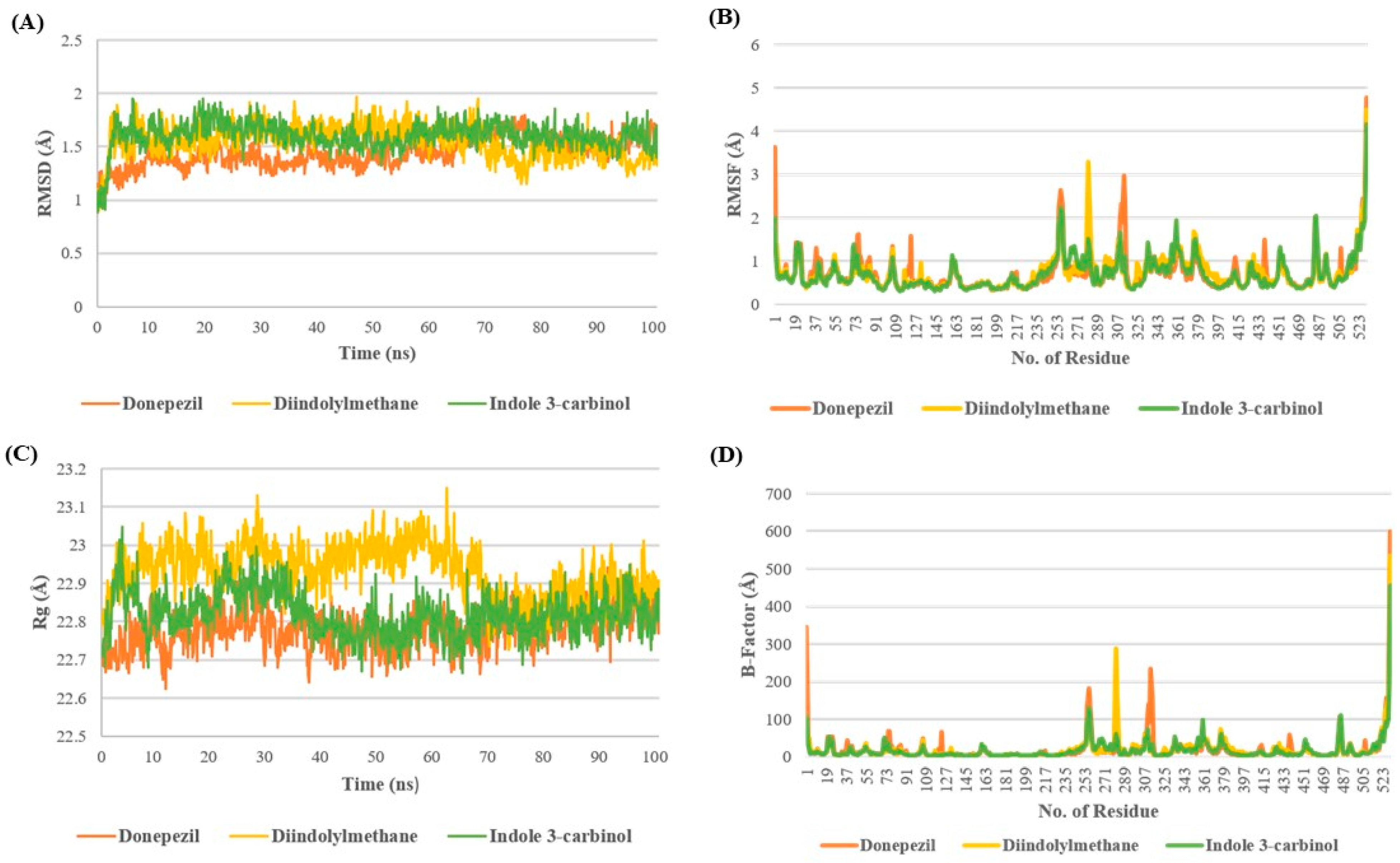
3.2. I3C and DIM Show High Stability with AchE
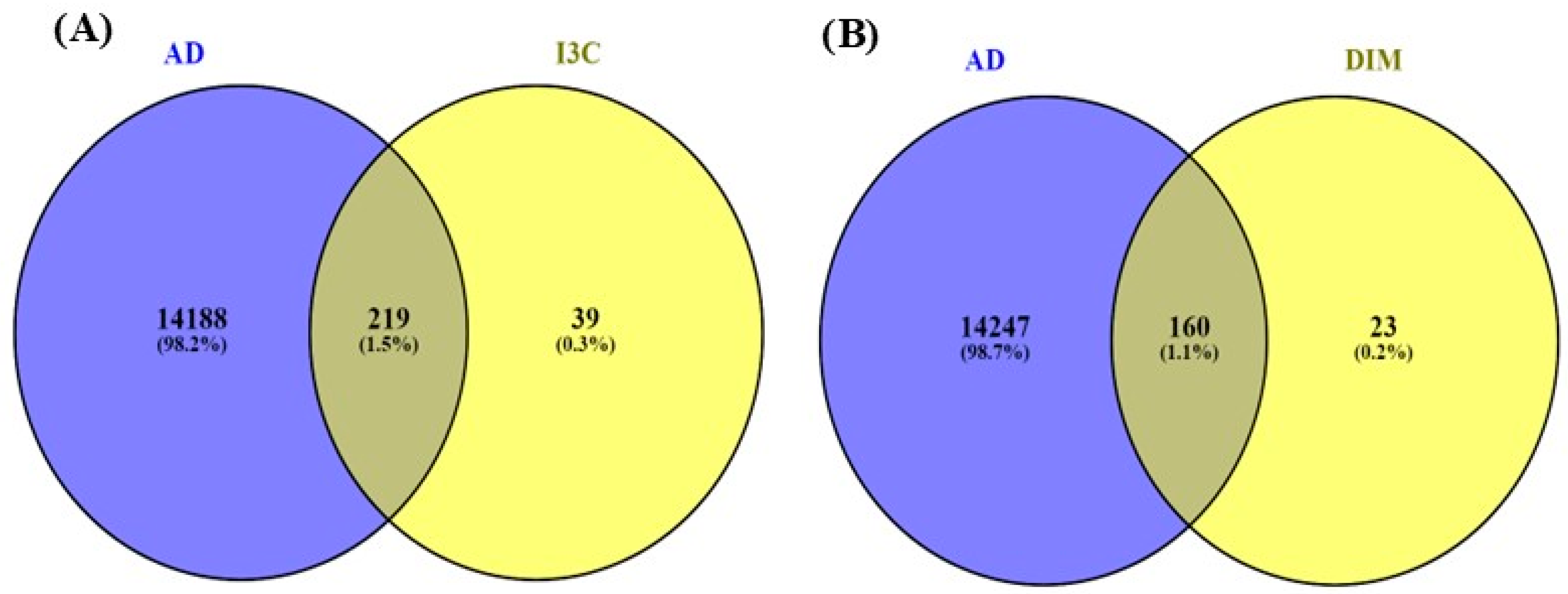
3.3. The Common Genes of I3C and DIM with AD
3.4. Protein–Protein Interaction Analysis
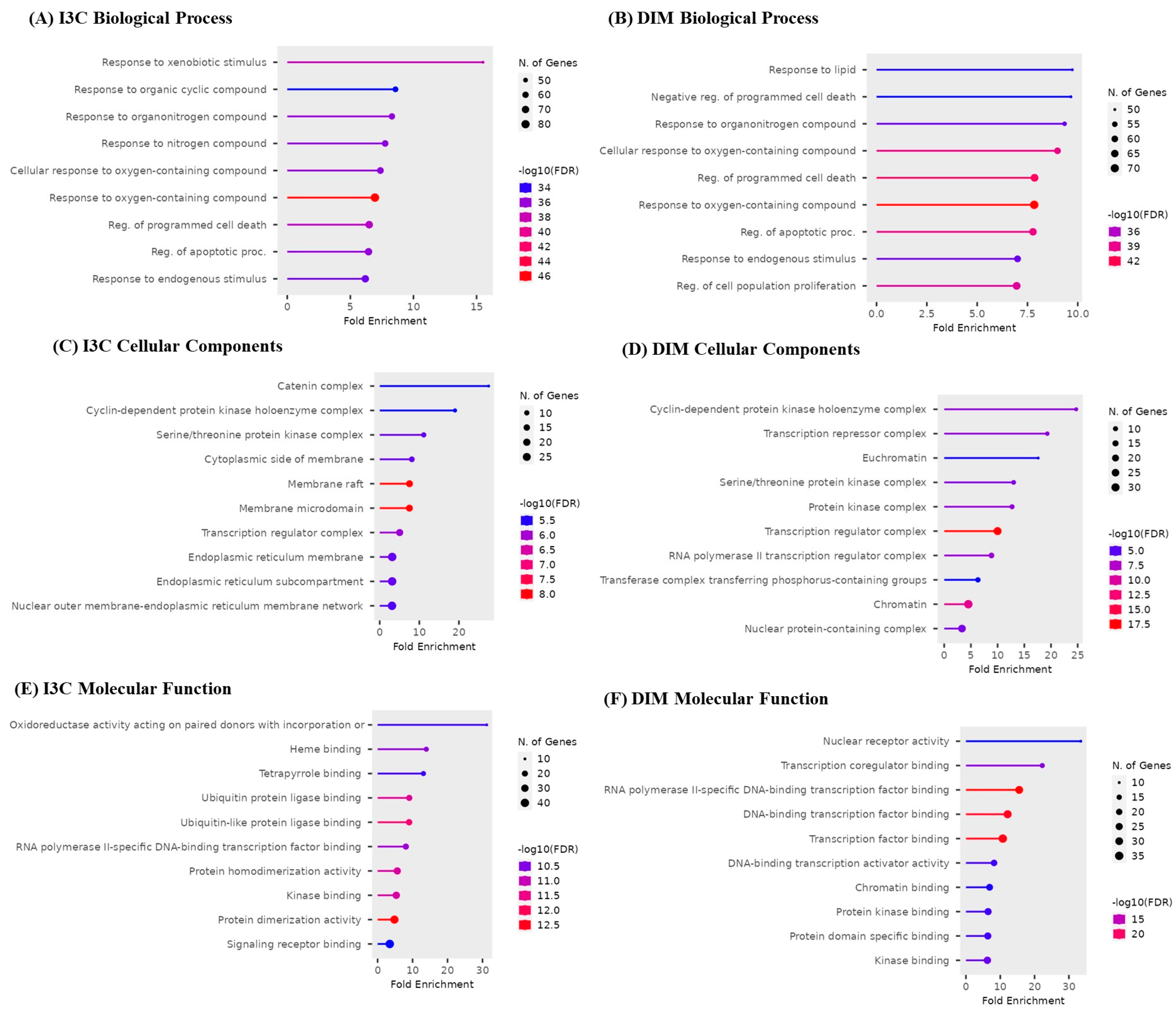
3.5. Gene Enrichment Analysis
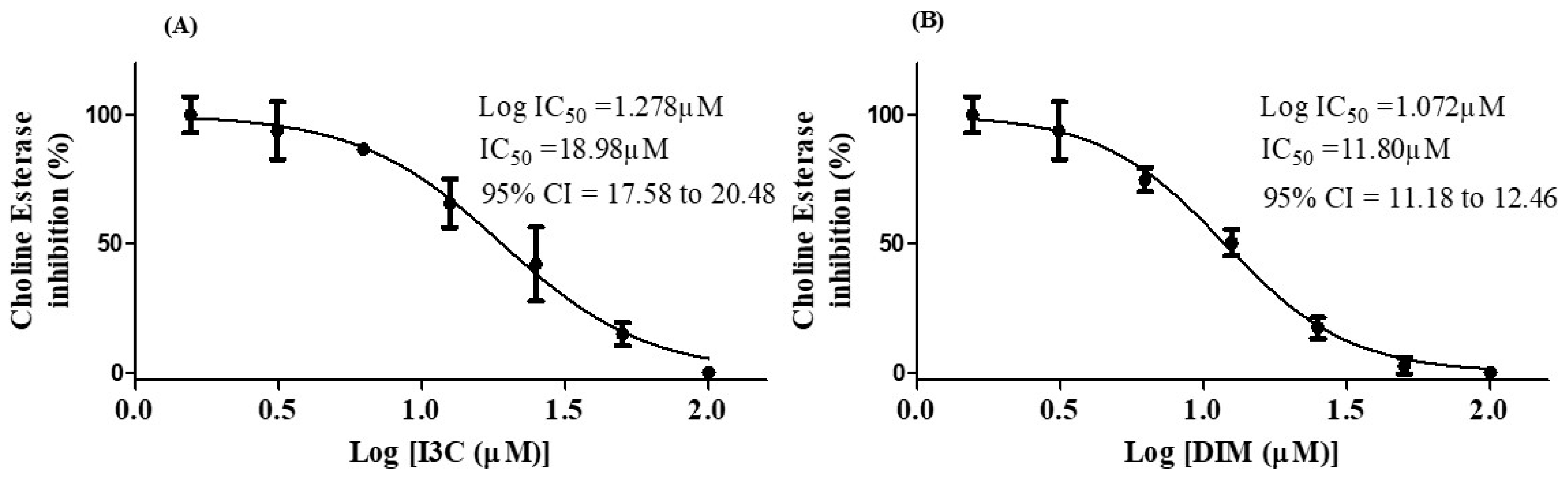
3.6. I3C and DIM Inhibited Choline esterase Enzyme Activity
3.7. I3C and DIM Inhibited Aβ Aggregation
3.8. I3C and DIM Mitigated Aβ Induced Neurotoxicity in SH-S5Y5 Cells
3.9. I3C and DIM Attenuated Oxidative Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nichols, E.; Szoeke, C.E.; Vollset, S.E.; Abbasi, N.; Abd-Allah, F.; Abdela, J.; Murray, C.J. Global, regional, and national burden of Alzheimer’s disease and other dementias, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 88–106. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Feng, X.; Sun, X.; Hou, N.; Han, F.; Liu, Y. Global, regional, and national burden of Alzheimer’s disease and other dementias, 1990–2019. Front. Aging Neurosci. 2022, 14, 937486. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, A. A review on Alzheimer’s disease pathophysiology and its management: An update. Pharmacol. Rep. 2015, 67, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Martorana, A.; Esposito, Z.; Koch, G. Beyond the cholinergic hypothesis: Do current drugs work in Alzheimer’s disease? CNS Neurosci. Ther. 2010, 16, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Bartolini, M.; Bertucci, C.; Cavrini, V.; Andrisano, V. β-Amyloid aggregation induced by human acetylcholinesterase: Inhibition studies. Biochem. Pharmacol. 2003, 65, 407–416. [Google Scholar] [CrossRef]
- Pardo-Moreno, T.; González-Acedo, A.; Rivas-Domínguez, A.; García-Morales, V.; García-Cozar, F.J.; Ramos-Rodríguez, J.J.; Melguizo-Rodríguez, L. Therapeutic approach to Alzheimer’s disease: Current treatments and new perspectives. Pharmaceutics 2022, 14, 1117. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.N.; Srivastava, P.; Sharma, P.; Tripathi, M.K.; Seth, A.; Tripathi, A.; Shrivastava, S.K. Biphenyl-3-oxo-1,2,4-triazine linked piperazine derivatives as potential cholinesterase inhibitors with antioxidant property to improve the learning and memory. Bioorg. Chem. 2019, 85, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Rai, S.N.; Singh, C.; Singh, A.; Singh, M.P.; Singh, B.K. Mitochondrial Dysfunction: A Potential Therapeutic Target to Treat Alzheimer’s Disease. Mol. Neurobiol. 2020, 57, 3075–3088. [Google Scholar] [CrossRef]
- Tripathi, P.N.; Lodhi, A.; Rai, S.N.; Nandi, N.K.; Dumoga, S.; Yadav, P.; Chaudhary, S. Review of Pharmacotherapeutic Targets in Alzheimer’s Disease and Its Management Using Traditional Medicinal Plants. Degener. Neurol. Neuromuscul. Dis. 2024, 14, 47–74. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Ichikawa, H. Molecular targets and anticancer potential of indole-3-carbinol and its derivatives. Cell Cycle 2005, 4, 1201–1215. [Google Scholar] [CrossRef]
- Singh, A.A.; Patil, M.P.; Kang, M.J.; Niyonizigiye, I.; Kim, G.D. Biomedical application of Indole-3-carbinol: A mini-review. Phytochem. Lett. 2021, 41, 49–54. [Google Scholar] [CrossRef]
- Singh, A.A.; Yadav, D.; Khan, F.; Song, M. Indole-3-Carbinol and Its Derivatives as Neuroprotective Modulators. Brain Sci. 2024, 14, 674. [Google Scholar] [CrossRef]
- Ramakrishna, K.; Jain, S.K.; Krishnamurthy, S. Pharmacokinet. Pharmacodyn. Prop. Indole-3-Carbinol Exp. Focal Ischemic Injury. Eur. J. Drug Metab. Pharmacokinet. 2022, 47, 593–605. [Google Scholar]
- Paliwal, P.; Chauhan, G.; Gautam, D.; Dash, D.; Patne, S.C.; Krishnamurthy, S. Indole-3-carbinol improves neurobehavioral symptoms in a cerebral ischemic stroke model. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2018, 391, 613–625. [Google Scholar] [CrossRef]
- Kakarla, R.; Karuturi, P.; Siakabinga, Q.; Kasi Viswanath, M.; Dumala, N.; Guntupalli, C.; Gujjari, L. Current understanding and future directions of cruciferous vegetables and their phytochemicals to combat neurological diseases. Phytother. Res. 2024, 38, 1381–1399. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, K.; Krishnamurthy, S. Indole-3-carbinol ameliorated the neurodevelopmental deficits in neonatal anoxic injury in rats. Int. J. Dev. Neurosci. 2023, 83, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Ma, Y.; Yang, R.; Lu, X.; You, Q.; Ye, T.; Huang, C. Indole-3-carbinol selectively prevents chronic stress-induced depression-but not anxiety-like behaviors via suppressing pro-inflammatory cytokine production and oxido-nitrosative stress in the brain. Front. Pharmacol. 2022, 13, 331. [Google Scholar] [CrossRef]
- El-Naga, R.N.; Ahmed, H.I.; Al Haleem, E.N.A. Effects of indole-3-carbinol on clonidine-induced neurotoxicity in rats: Impact on oxidative stress, inflammation, apoptosis and monoamine levels. Neurotoxicology 2014, 44, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, K.; Sinku, S.; Majumdar, S.; Singh, N.; Gajendra, T.A.; Rani, A.; Krishnamurthy, S. Indole-3-carbinol ameliorated the thioacetamide-induced hepatic encephalopathy in rats. Toxicology 2023, 492, 153542. [Google Scholar] [CrossRef]
- Mohamad, K.A.; El-Naga, R.N.; Wahdan, S.A. Neuroprotective effects of indole-3-carbinol on the rotenone rat model of Parkinson’s disease: Impact of the SIRT1-AMPK signaling pathway. Toxicol. Appl. Pharmacol. 2022, 435, 115853. [Google Scholar] [CrossRef]
- Saini, N.; Akhtar, A.; Chauhan, M.; Dhingra, N.; Sah, S.P. Protective effect of Indole-3-carbinol, an NF-κB inhibitor in experimental paradigm of Parkinson’s disease: In silico and in vivo studies. Brain Behav. Immun. 2020, 90, 108–137. [Google Scholar] [CrossRef]
- Bradlow, H.; Zeligs, M. Diindolylmethane (DIM) spontaneously forms from indole-3-carbinol (I3C) during cell culture experiments. In Vivo 2010, 24, 387–391. [Google Scholar]
- Ramakrishna, K.; Singh, N.; Krishnamurthy, S. Diindolylmethane ameliorates platelet aggregation and thrombosis: In silico, in vitro, and in vivo studies. Eur. J. Pharmacol. 2022, 919, 174812. [Google Scholar] [CrossRef]
- Dhir, N.; Jain, A.; Sharma, A.R.; Prakash, A.; Bhatia, A.; Medhi, B. Neuroprotective effect of 3,3′-diindolylmethane and ɑ-naphthoflavone, aryl hydrocarbon receptor modulators in an experimental model of ischemic stroke. CNS Neurol. Disord. Drug Targets 2022, 21, e180422203743. [Google Scholar] [CrossRef]
- Ramakrishna, K.; Singh, S.K.; Krishnamurthy, S. Diindolylmethane Ameliorates Ischemic Stroke-Induced Brain Injury by Peripheral and Central Mechanisms. Curr. Neurovascular Res. 2022, 19, 462–475. [Google Scholar]
- Rzemieniec, J.; Bratek, E.; Wnuk, A.; Przepiórska, K.; Salińska, E.; Kajta, M. Neuroprotective effect of 3,3′-Diindolylmethane against perinatal asphyxia involves inhibition of the AhR and NMDA signaling and hypermethylation of specific genes. Apoptosis 2020, 25, 747–762. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Kinoshita, K.; Yoshimizu, A.; Kurauchi, Y.; Hisatsune, A.; Seki, T.; Katsuki, H. Laquinimod and 3,3′-diindolylemethane alleviate neuropathological events and neurological deficits in a mouse model of intracerebral hemorrhage. J. Neuroimmunol. 2020, 342, 577195. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Tan, L.; Chen, Y.; Liu, A.; Hong, M.; Peng, Z. DIM mitigates the development of experimental autoimmune encephalomyelitis by maintaining the stability and suppressive function of regulatory T cells. Cell. Immunol. 2020, 358, 104238. [Google Scholar] [CrossRef] [PubMed]
- Madison, C.A.; Kuempel, J.; Albrecht, G.L.; Hillbrick, L.; Jayaraman, A.; Safe, S.; Eitan, S. 3,3′-Diindolylmethane and 1,4-dihydroxy-2-naphthoic acid prevent chronic mild stress induced depressive-like behaviors in female mice. J. Affect. Disord. 2022, 309, 201–210. [Google Scholar] [CrossRef]
- Mattiazzi, J.; Sari MH, M.; de Bastos Brum, T.; Araújo PC, O.; Nadal, J.M.; Farago, P.V.; Cruz, L. 3,3′-Diindolylmethane nanoencapsulation improves its antinociceptive action: Physicochemical and behavioral studies. Colloids Surf. B Biointerfaces 2019, 181, 295–304. [Google Scholar] [CrossRef]
- Mattiazzi, J.; Sari, M.H.M.; Araujo, P.C.O.; Englert, A.V.; Nadal, J.M.; Farago, P.V.; Cruz, L. Ethylcellulose microparticles enhance 3,3′-diindolylmethane anti-hypernociceptive action in an animal model of acute inflammatory pain. J. Microencapsul. 2020, 37, 528–541. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Gutti, G.; Leifeld, J.; Kakarla, R.; Bajad, N.G.; Ganeshpurkar, A.; Kumar, A.; Singh, S.K. Discovery of triazole-bridged aryl adamantane analogs as an intriguing class of multifunctional agents for treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2023, 259, 115670. [Google Scholar] [CrossRef] [PubMed]
- Gutti, G.; Kumar, D.; Paliwal, P.; Ganeshpurkar, A.; Lahre, K.; Kumar, A.; Singh, S.K. Development of pyrazole and spiropyrazoline analogs as multifunctional agents for treatment of Alzheimer’s disease. Bioorg. Chem. 2019, 90, 103080. [Google Scholar] [CrossRef] [PubMed]
- Bhanukiran, K.; Singh, R.; Gajendra, T.A.; Ramakrishna, K.; Singh, S.K.; Krishnamurthy, S.; Hemalatha, S. Vasicinone, a pyrroloquinazoline alkaloid from Adhatoda vasica Nees enhances memory and cognition by inhibiting cholinesterases in Alzheimer’s disease. Phytomed. Plus 2023, 3, 100439. [Google Scholar] [CrossRef]
- Kumar, D.; Gupta, S.K.; Ganeshpurkar, A.; Gutti, G.; Krishnamurthy, S.; Modi, G.; Singh, S.K. Development of Piperazinediones as dual inhibitor for treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2018, 150, 87–101. [Google Scholar] [CrossRef]
- Gutti, G.; Kakarla, R.; Kumar, D.; Beohar, M.; Ganeshpurkar, A.; Kumar, A.; Singh, S.K. Discovery of novel series of 2-substituted benzo[d]oxazol-5-amine derivatives as multi-target directed ligands for the treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2019, 182, 111613. [Google Scholar] [CrossRef]
- Mhillaj, E.; Papi, M.; Paciello, F.; Silvestrini, A.; Rolesi, R.; Palmieri, V.; Mancuso, C. Celecoxib Exerts Neuroprotective Effects in β-Amyloid-Treated SH-SY5Y Cells through the Regulation of Heme Oxygenase-1: Novel Insights for an Old Drug. Front. Cell Dev. Biol. 2020, 8, 561179. [Google Scholar] [CrossRef]
- Singh, M.; Agarwal, V.; Pancham, P.; Jindal, D.; Agarwal, S.; Rai, S.N.; Gupta, V. A Comprehensive Review and Androgen Deprivation Therapy and Its Impact on Alzheimer’s Disease Risk in Older Men with Prostate Cancer. Degener. Neurol. Neuromuscul. Dis. 2024, 14, 33–46. [Google Scholar] [CrossRef]
- Srivastava, P.; Tripathi, P.N.; Sharma, P.; Rai, S.N.; Singh, S.P.; Srivastava, R.K.; Shrivastava, S.K. Design and development of some phenyl benzoxazole derivatives as a potent acetylcholinesterase inhibitor with antioxidant property to enhance learning and memory. Eur. J. Med. Chem. 2019, 163, 116–135. [Google Scholar] [CrossRef]


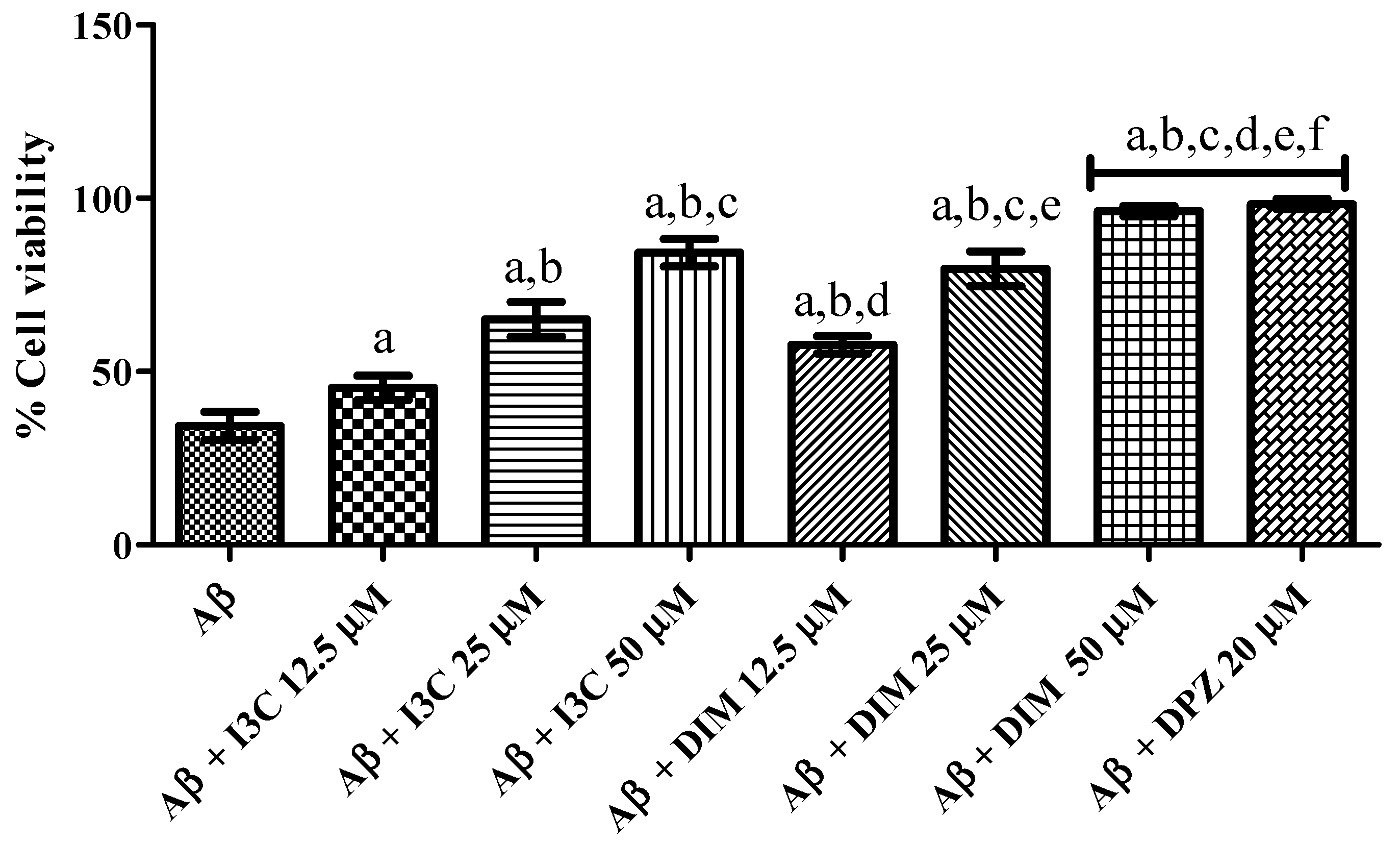


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Ligands Name | Binding Affinity | Interactions | ||||
|---|---|---|---|---|---|---|---|
| Hydrogen Bonds | Pi-Pi Stacking | Pi-Alkyl | Pi-Sigma | Pi-Anion | |||
| 1 | Indole 3-Carbinol | −7.0 | Tyr124, Ser203 | Tyr337, Phe338, Tyr341 | - | - | - |
| 2 | Diindolylmethane | −10.3 | - | Tyr341, Trp86 | - | - | Asp74 |
| 3 | Donepezil (DPZ) | −12.0 | Tyr72, Phe295 | Tyr341, Trp286, Trp86 | Trp286, Tyr341, Phe338, Tyr337 | Trp286 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramakrishna, K.; Karuturi, P.; Siakabinga, Q.; T.A., G.; Krishnamurthy, S.; Singh, S.; Kumari, S.; Kumar, G.S.; Sobhia, M.E.; Rai, S.N. Indole-3 Carbinol and Diindolylmethane Mitigated β-Amyloid-Induced Neurotoxicity and Acetylcholinesterase Enzyme Activity: In Silico, In Vitro, and Network Pharmacology Study. Diseases 2024, 12, 184. https://doi.org/10.3390/diseases12080184
Ramakrishna K, Karuturi P, Siakabinga Q, T.A. G, Krishnamurthy S, Singh S, Kumari S, Kumar GS, Sobhia ME, Rai SN. Indole-3 Carbinol and Diindolylmethane Mitigated β-Amyloid-Induced Neurotoxicity and Acetylcholinesterase Enzyme Activity: In Silico, In Vitro, and Network Pharmacology Study. Diseases. 2024; 12(8):184. https://doi.org/10.3390/diseases12080184
Chicago/Turabian StyleRamakrishna, Kakarla, Praditha Karuturi, Queen Siakabinga, Gajendra T.A., Sairam Krishnamurthy, Shreya Singh, Sonia Kumari, G. Siva Kumar, M. Elizabeth Sobhia, and Sachchida Nand Rai. 2024. "Indole-3 Carbinol and Diindolylmethane Mitigated β-Amyloid-Induced Neurotoxicity and Acetylcholinesterase Enzyme Activity: In Silico, In Vitro, and Network Pharmacology Study" Diseases 12, no. 8: 184. https://doi.org/10.3390/diseases12080184