Grape Pomace: Antioxidant Activity, Potential Effect Against Hypertension and Metabolites Characterization after Intake


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Materials
2.3. Animals for In Vivo Experiences
2.4. Samples Preparation
2.4.1. Grape Pomace Samples Preparation
2.4.2. Sample Collection from In Vivo Experiments
2.5. Analytical Methods
2.5.1. Antioxidant Assays
2.5.2. Blood Pressure Measurement
2.5.3. Extraction of Phenolics from Tissues
2.5.4. Extraction of Phenolics from Plasmas
2.5.5. Urine Analysis
2.5.6. HPLC-PDA-ESI-MSn Analysis of Procyanidin Metabolites
2.5.7. Statistical Analyses
3. Results and Discussion
3.1. Grape Pomace Seed and Skin Antioxidant Activities Evaluation
3.2. In Vivo Results
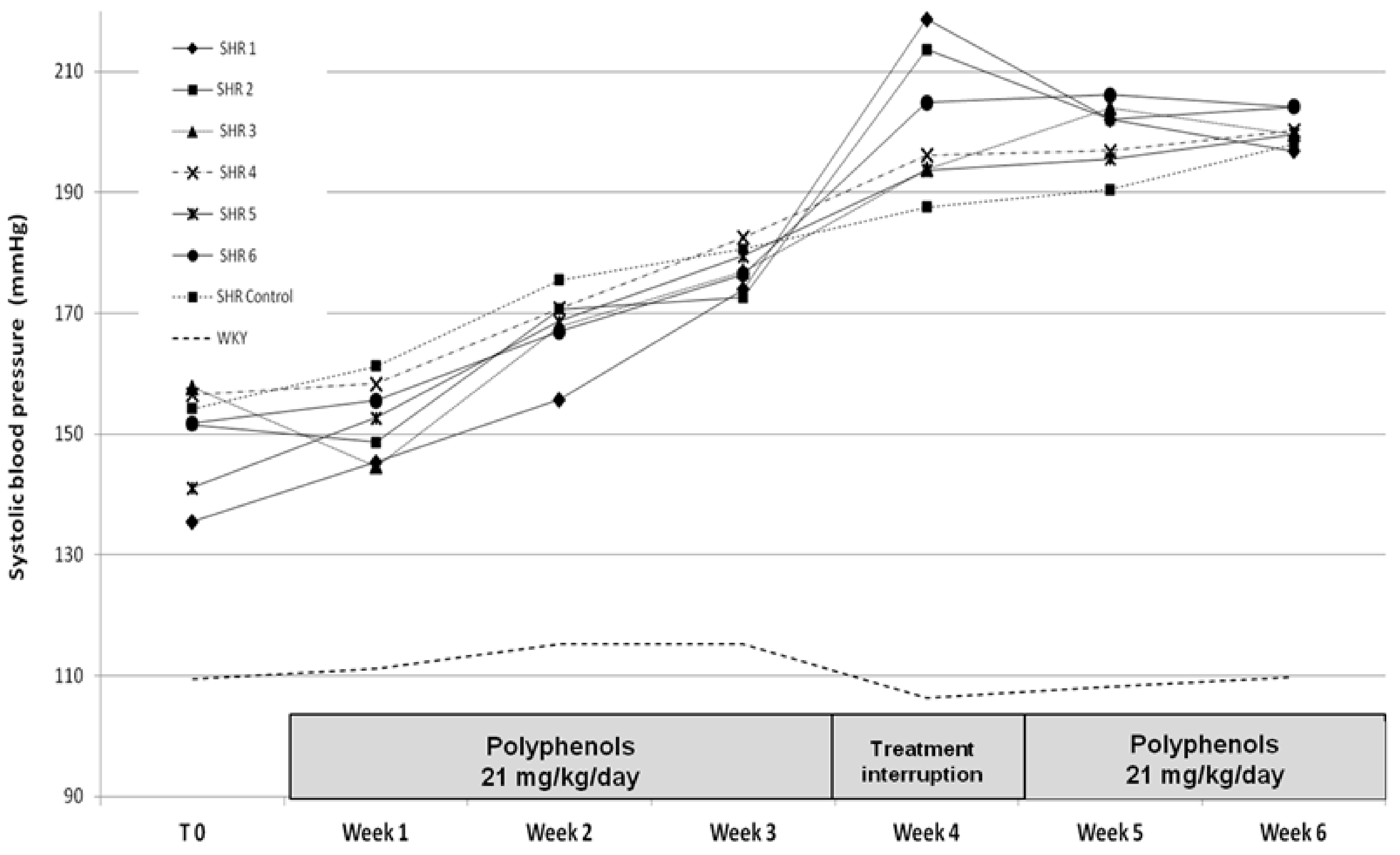
3.2.1. Systolic Blood Pressure Results
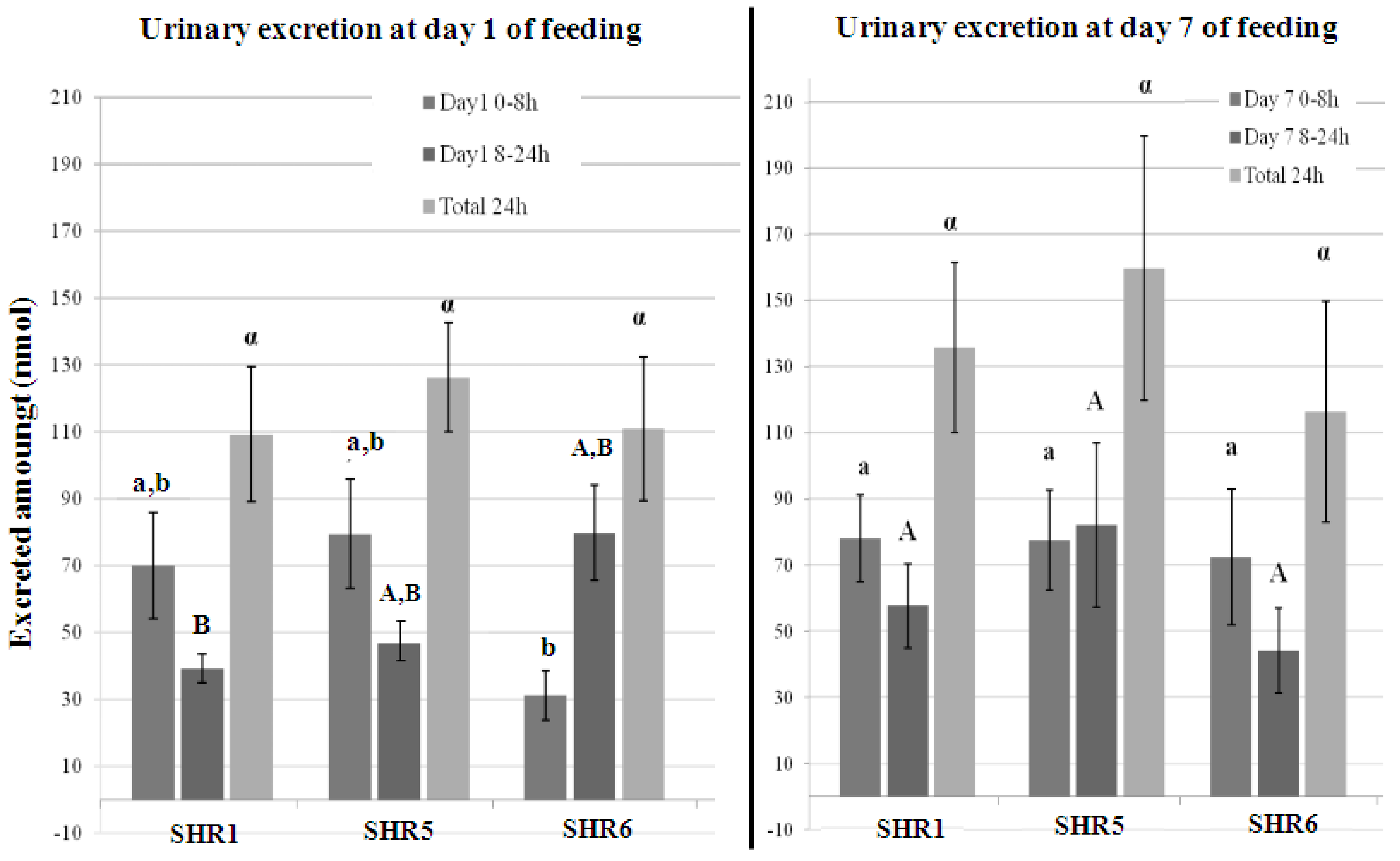
3.2.2. Quantification of Polyphenolic Metabolites in Urine
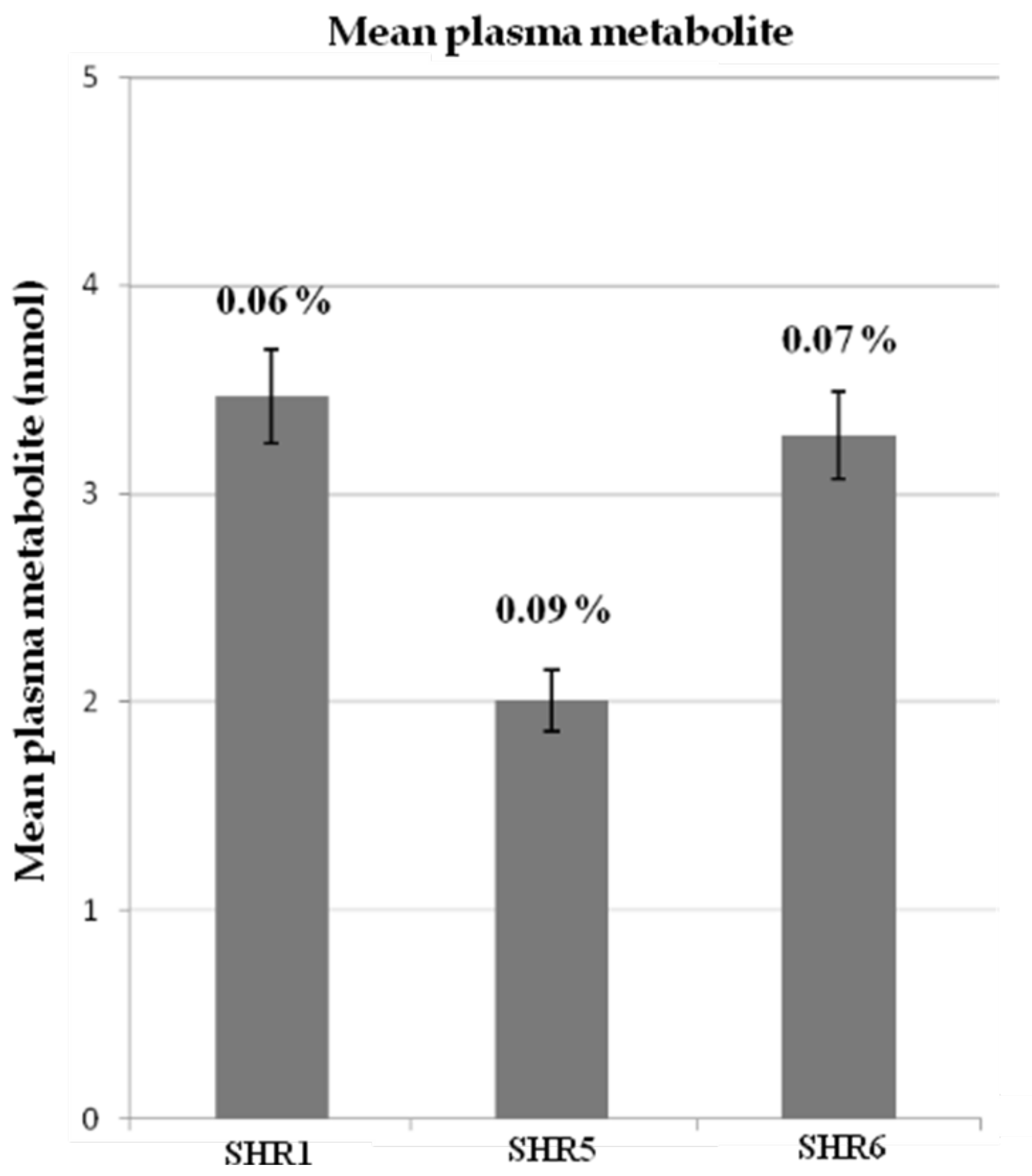
3.2.3. HPLC-ESI-MSn Analysis of SHR Rat Plasma
3.2.4. HPLC-ESI-MSn Analysis of SHR Rat Tissues (Heart, Liver and Kidneys)
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weisshaar, B.; Jenkins, G.I. Phenylpropanoid biosynthesis and its regulation. Curr. Opin. Plant Biol. 1998, 3, 251–257. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Biosynthesis if flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Jonhson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81, 215S–217S. [Google Scholar] [CrossRef] [PubMed]
- Renaud, S.; de Lorgeril, M. Wine, alcohol, platelets, and the French paradox for coronary heart disease. Lancet 1992, 339, 1523–1526. [Google Scholar] [CrossRef]
- Cordova, A.C.; Jackson, L.S.; Berke-Schlessel, D.W.; Sumpio, B.E. The cardiovascular protective effect of red wine. J. Am. Coll. Surg. 2005, 200, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Dohadwala, M.M.; Vita, J.A. Grapes and cardiovascular disease. J. Nutr. 2009, 139, 1788S–1793S. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Franchini, M.; Favaloro, E.J.; Targher, G. Moderate red wine consumption and cardiovascular disease risk: Beyond the French paradox. Semin. Thromb. Hemost. 2010, 36, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Rasines-Perea, Z.; Teissedre, P.L. Grape Polyphenols’ Effects in Human Cardiovascular Diseases and Diabetes. Molecules 2017, 22, 68. [Google Scholar] [CrossRef] [PubMed]
- Brenn, T. The Tromsø heart study: Alcoholic beverages and coronary risk factors. J. Epidemiol. Community Health 1986, 40, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Van de Wiel, A.; de Lange, D.W. Cardiovascular risk is more related to drinking pattern than to the type of alcoholic drinks. Neth. J. Med. 2008, 66, 467–473. [Google Scholar] [PubMed]
- Bulpitt, C.J.; Shipley, M.J.; Semmence, A. The contribution of a moderate intake of alcohol to the presence of hypertension. J. Hypertens. 1987, 5, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Zilkens, R.R.; Burke, V.; Hodgson, J.M.; Barden, A.; Beilin, L.J.; Puddey, I.B. Red wine and beer elevate blood pressure in normotensive men. Hypertension 2005, 45, 874–879. [Google Scholar] [CrossRef] [PubMed]
- Hassellund, S.S.; Flaa, A.; Sandvik, L.; Kjeldsen, S.E.; Rostrup, M. Effects of anthocyanins on blood pressure and stress reactivity: A double-blind randomized placebo-controlled crossover study. J. Hum. Hypertens. 2012, 26, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Spaak, J.; Merlocco, A.C.; Soleas, G.J.; Tomlinson, G.; Morris, B.L.; Picton, P.; Notarius, C.F.; Chan, C.T.; Floras, J.S. Dose-related effects of red wine and alcohol on hemodynamics, sympathetic nerve activity, and arterial diameter. Am. J. Physiol. 2008, 294, H605–H612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, A.P.; Sutherland, W.H.; McCormick, M.P.; Yeoman, D.J.; de Jong, S.A.; Williams, M.J. Effects of white and red wine on endothelial function in subjects with coronary artery disease. Int. Med. J. 2004, 34, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Haque, A.M.; Hashimoto, M.; Katakura, M.; Tanabe, Y.; Hara, Y.; Shido, O. Long-term administration of green tea catechins improves spatial cognition learning ability in rats. J. Nutr. 2006, 136, 1043–1047. [Google Scholar] [CrossRef] [PubMed]
- Naissides, M.; Pal, S.; Mamo, J.C.; James, A.P.; Dhaliwal, S. The effect of chronic consumption of red wine polyphenols on vascular function in postmenopausal women. Eur. J. Clin. Nutr. 2006, 60, 740–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosso, G.; Godos, J.; Lamuela-Raventos, R.; Ray, S.; Micek, A.; Pajak, A.; Sciacca, S.; D’Orazio, N.; Del Rio, D.; Galvano, F. A comprehensive meta-analysis on dietary flavonoid and lignan intake and cancer risk: Level of evidence and limitations. Mol. Nutr. Food Res. 2017, 61, 1600930. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jiménez, J.; Hubert, J.; Hooper, L.; Cassidy, A.; Manach, C.; Williamson, G.; Scalbert, A. Urinary metabolites as biomarkers of polyphenol intake in humans: A systematic review. Am. J. Clin. Nutr. 2010, 92, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Llobera, A.; Cañellas, J. Dietary fibre content and antioxidant activity of Mano Negro red grape (Vitis vinifera): Pomace and stem. Food Chem. 2007, 101, 659–666. [Google Scholar] [CrossRef]
- Laufenberg, G.; Kunz, B.; Nystroem, M. Transformation of vegetable waste into value added products: (A) the upgrading concept; (B) practical implementations. Bioresour. Technol. 2003, 87, 167–198. [Google Scholar] [CrossRef]
- Tarascou, I.; Barathieu, K.; André, Y.; Pianet, I.; Dufourc, E.J.; Fouquet, E. An improved synthesis of procyanidin dimers: Regio- and stereocontrol of the interflavan bond. Eur. J. Org. Chem. 2006, 23, 5367–5377. [Google Scholar] [CrossRef]
- Ky, I.; Lorrain, B.; Kolbas, N.; Crozier, A.; Teissedre, P.L. Wine by-products: Phenolic characterization and antioxidant activity evaluation of grape and grape pomace from six different French grape varieties. Molecules 2014, 19, 482–506. [Google Scholar] [CrossRef] [PubMed]
- Ky, I.; Teissedre, P.L. Characterization of Mediterranean grape pomace seed and skin extracts: Polyphenolic content and antioxidant activity. Molecules 2015, 20, 2190–2207. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, J.I.; Momma, T.Y.; Kuhnle, G.K.; Keen, C.L.; Schroeter, H. Structurally related (−)-epicatechin metabolites in humans: Assessment using de novo chemically synthesized authentic standards. Free Radic. Biol. Med. 2012, 52, 1403–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Niki, E. Antioxidant capacity: Which capacity and how to assess it? J. Berry Res. 2011, 1, 169–176. [Google Scholar] [CrossRef]
- Chiva-Blanch, G.; Visioli, F. Polyphenols and health: Moving beyond antioxidants. J. Berry Res. 2012, 2, 63–71. [Google Scholar] [CrossRef]
- Al-Awwadi, N.A.; Bornet, A.; Azay, J.; Araiz, C.; Delbosc, S.; Cristol, J.P.; Linck, N.; Cros, G.; Teissedre, P.L. Red wine polyphenols alone or in association with ethanol prevent hypertension, cardiac hypertrophy, and production of reactive oxygen species in the insulin-resistant fructose-fed rat. J. Agric. Food Chem. 2004, 52, 5593–5597. [Google Scholar] [CrossRef] [PubMed]
- Bravo, L.; Herrera, M.D.; Marhuenda, E.; Perez-Guerrero, C. Cardiovascular effects of lovastatin in normotensive and spontaneously hypertensive rats. Gen. Pharmacol. 1998, 30, 331–336. [Google Scholar] [CrossRef]
- Mukai, Y.; Sato, S. Polyphenol-containing azuki bean (Vigna angularis) seed coats attenuate vascular oxidative stress and inflammation in spontaneously hypertensive rats. J. Nutr. Biochem. 2011, 22, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, M.; Sánchez, D.; Muguerza, B.; Moulay, L.; Laghi, S.; Miguel, M.; Aleixandre, A. The blood pressure effect and related plasma levels of flavan-3-ols in spontaneously hypertensive rats. Food Chem. 2010, 122, 3479–3489. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Sanada, T.; Ojima, M.; Kanagawa, R.; Nishikawa, K.; Inada, Y. Combined effects of the angiotensin II antagoninst candesartatn cilexetil (TCV-116) and other classes of antihypertensive drugs in spontaneously hypertensive rats. Hypertens. Res. 1996, 19, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.C.; Jhou, K.Y.; Tseng, C.Y. Antihypertensive effect on mulberry leaf aqueous extract containing γ-aminobutyric acid in spontaneously hypertensive rats. Food Chem. 2012, 132, 1796–1801. [Google Scholar] [CrossRef]
- Tsang, C.; Auger, C.; Mullen, W.; Bornet, A.; Rouanet, J.M.; Crozier, A.; Teissedre, P.L. The absorption, metabolism and excretion of flavan-3-ols and procyanidins following the ingestion of a grape seed extract by rats. Br. J. Nutr. 2005, 94, 170–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touriño, S.; Fuguet, E.; Vinardell, M.P.; Cascante, M.; Torres, J.L. Phenolic metabolites of grape antioxidant dietary fiber in rat urine. J. Agric. Food Chem. 2009, 57, 11418–11426. [Google Scholar] [CrossRef] [PubMed]
- Serra, A.; Macià, A.; Romero, M.P.; Salvado, M.J.; Bustos, M.; Fernandez-Larrea, J.; Motilva, M.J. Determination of procyanidins and their metabolites in plasma samples by improved liquid chromatography-tandem mass spectrometry. J. Chromatogr. B 2009, 877, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Day, A.J.; Gee, J.M.; DuPont, M.S.; Johnson, I.T.; Williamson, G. Absorption of quercetin-3-glucoside and quercetin-4′-glucoside in the rat small intestine: The role of lactase phlorizin hydrolase and the sodium-dependent glucose transporter. Biochem. Pharmacol. 2003, 65, 1199–1206. [Google Scholar] [CrossRef]
- Llorach, R.; Garrido, I.; Monagas, M.; Urpi-Sarda, M.; Tulipani, S.; Bartolome, B.; Andres-Lacueva, C. Metabolomics study of human urinary metabolome modifications after intake of almond (Prunus dulcis (Mill.) D.A. Webb) skin polyphenols. J. Proteome Res. 2010, 9, 5859–5867. [Google Scholar] [CrossRef] [PubMed]
- Donovan, J.L.; Kasim-Karakas, S.; German, J.B.; Waterhouse, A.L. Urinary excretion of catechin metabolites by human subjects after red wine consumption. Br. J. Nutr. 2002, 87, 31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auger, C.; Mullen, W.; Hara, Y.; Crozier, A. Bioavailability of polyphenon E flavan-3-ols in humans with an Ileostomy. J. Nutr. 2008, 138, 1535–1542. [Google Scholar] [CrossRef] [PubMed]
- Urpi-Sarda, M.; Garrido, I.; Monagas, M.; Gomez-Cordoves, C.; Medina-Remon, A.; Andres La cueva, C.; Bartolome, B. Profile of plasma and urine metabolites after the intake of almond [Prunus dulcis (Mill.) D.A. Webb] polyphenols in humans. J. Agric. Food Chem. 2009, 57, 10134–10142. [Google Scholar] [CrossRef] [PubMed]
- Van der Hooft, J.J.; de Vos, R.C.; Mihaleva, V.; Bino, R.J.; Ridder, L.; de Roo, N.; Jacobs, D.M.; van Duynhoven, J.P.; Vervoort, J. Structural elucidation and quantification of phenolic conjugates present in human urine after tea intake. Anal. Chem. 2012, 84, 7263–7271. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| ORAC 2 | FRAP 2 | ABTS•+ 2 | DPPH 2 | ||
|---|---|---|---|---|---|
| Seeds-EAQ | GRE1 1 | 1466.4 ± 29.6 a | 0.63 ± 0.02 a | 1203.2 ± 24.1 a | 410.8 ± 43.3 a |
| SYR1 1 | 2230.7 ± 101.7 b | 1.33 ± 0.08 c | 2432.6 ± 56.0 c | 1037.1 ± 64.0 b | |
| CAR 1 | 2058.6 ± 85.1 b | 1.06 ± 0.08 b | 1948.8 ± 61.1 b | 1050.6 ± 30.1 b | |
| Seeds-EA70 | GRE1 1 | 1926.7 ± 108.6 a | 1.28 ± 0.01 a | 2813.2 ± 90.0 a | 1277.6 ± 54.7 a |
| SYR1 1 | 2614.0 ± 150.9 a | 1.45 ± 0.16 a | 3601.2 ± 88.6 b | 1685.9 ± 130.7 b | |
| CAR 1 | 2332.9 ± 91.9 a | 1.20 ± 0.06 a | 3495.6 ± 66.4 b | 1536.8 ± 38.9 b |
| ORAC 2 | FRAP 2 | ABTS•+ 2 | DPPH 2 | ||
|---|---|---|---|---|---|
| Skins-EAQ | GRE2 1 | 1190.7 ± 183.6 ab | 0.56 ± 0.01 c | 934.1 ± 11.9 b | 99.5 ± 10.8 a |
| SYR1 1 | 1345.9 ± 19.2 ab | 0.88 ± 0.01 e | 1428,0 ± 54.8 c | 690.3 ± 147.0 bc | |
| SYR2 1 | 1066,0 ± 84.2 a | 0.14 ± 0.02 a | 668.3 ± 30.0 a | 263.9 ± 71.5 ab | |
| CAR 1 | 1077.8 ± 60.2 a | 0.67 ± 0.02 d | 1048.8 ± 101.6 b | 591.0 ± 85.6 abc | |
| MOU 1 | 1033.8 ± 77.6 a | 0.32 ± 0.01 b | 965.6 ± 16.6 b | 279.4 ± 61.7 ab | |
| ALI 1 | 1714.6 ± 14.8 b | 1.13 ± 0.00 f | 1760.1 ± 91.0 d | 1057.1 ± 45.2 c | |
| Skins-EA70 | GRE2 1 | 1828.3 ± 40.4 bc | 1.32 ± 0.03 c | 2612.1 ± 130.9 a | 877,0 ± 74.3 a |
| SYR1 1 | 1912.6 ± 6.1 bc | 1.52 ± 0.05 d | 2614.5 ± 10.4 a | 1391.7 ± 37.2 bc | |
| SYR2 1 | 1701.8 ± 88.3 bc | 0.94 ± 0.03 a | 2010.6 ± 147.0 a | 1164.9 ± 55.6 ab | |
| CAR 1 | 1238.4 ± 11.1 a | 1.34 ± 0.03 c | 2555.9 ± 146.0 a | 1075.4 ± 46.2 ab | |
| MOU 1 | 2070.0 ± 60.6 c | 1.03 ± 0.02 ab | 2674.8 ± 187.3 a | 833.3 ± 26.4 a | |
| ALI 1 | 1628.5 ± 82.6 b | 1.13 ± 0.01 b | 1923.4 ± 87.0 a | 1749.3 ± 112.7 c |
| Σof Structurally Related Metabolites(Day 1, 0–8 h) | SHR1 | SHR5 | SHR6 | Control |
| (Epi)catechin-O-glucuronide | 1.0 ± 0.3 * | 2.0 ± 0.2 * | 1.2 ± 0.5 * | 0.1. ± 0.0 |
| O-Methyl-(epi)catechin-O-glucuronide | 8.5 ± 1.5 * | 3.5 ± 0.4 | 6.1 ± 2.7 | 3.5 ± 1.0 |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-glucuronide | 1.0 ± 0.1 * | 1.2 ± 0.4 | 0.5 ± 0.3 | 0.4 ± 0.2 |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-sulphate | 1.5 ± 0.3 | 4.4 ± 2.9 | 1.2 ± 0.4 | 1.1 ± 0.3 |
| 5-(Hydroxyphenyl)-γ-hydroxyvaleric acid-O-sulphate | 2.5 ± 0.6 * | 3.5 ± 1.3 | 2.0 ± 0.5 * | 0.9 ± 0.3 |
| 5-(Phenyl)-γ-hydroxyvaleric acid-O-sulphate | 6.9 ± 1.3 * | 7.6 ± 1.6 | 6.1 ± 3.7 | 3.1 ± 0.7 |
| Isoferulic acid-4-O-sulfate | 157.0 ± 35.5 * | 75.0 ± 15.6* | 82.8 ± 19.9 | 82.2 ± 19.9 |
| Σof Structurally Related Metabolites(Day 1, 8–24 h) | SHR1 | SHR5 | SHR6 | Control |
| (Epi)catechin-O-glucuronide | 1.1 ± 0.2 | 1.8 ± 0.6 | 1.4 ± 0.3 | 1.0 ± 0.3 |
| O-Methyl-(epi)catechin-O-glucuronide | 3.8 ± 0.7 | 6.9 ± 2.1 * | 7.1 ± 1.3 * | 2.7 ± 1.0 |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-glucuronide | 0.2 ± 0.0 | 0.4 ± 0.1 | 0.4 ± 0.1 * | 0.2 ± 0.0 |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-sulphate | 1.3 ± 0.2 | 1.8 ± 0.6 | 1.6 ± 0.2 | 1.3 ± 0.3 |
| 5-(Hydroxyphenyl)-γ-hydroxyvaleric acid-O-sulphate | 1.0 ± 0.1 | 2.6 ± 0.9 * | 2.2 ± 0.4 * | 0.7 ± 0.1 |
| 5-(Phenyl)-γ-hydroxyvaleric acid-O-sulphate | 12.7 ± 1.8 | 14.4 ± 2.5 | 15.8 ± 1.1 * | 8.7 ± 0.9 |
| Isoferulic acid-4-O-sulfate | 80.4 ± 13.6 | 99.7 ± 16.1 | 111.5 ± 25.7 * | 74.2 ± 4.3 |
| Σof Structurally Related Metabolites(Day 7, 0–8 h) | SHR1 | SHR5 | SHR6 | Control |
| (Epi)catechin-O-glucuronide | 1.6 ± 1.0 | 3.1 ± 1.9 | 2.9 ± 1.0 * | 0.5 ± 0.3 |
| O-Methyl-(epi)catechin-O-glucuronide | 5.7 ± 2.2 | 5.7 ± 2.3 | 5.9 ± 2.1 * | 1.7 ± 0.4 |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-glucuronide | 0.2 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.2 ± 0.1 |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-sulphate | 0.2 ± 0.0 | 0.4 ± 0.2 | 0.5 ± 0.2 | 0.2 ± 0.1 |
| 5-(Hydroxyphenyl)-γ-hydroxyvaleric acid-O-sulphate | 0.4 ± 0.1 | 0.6 ± 0.1 * | 0.9 ± 0.4 | 0.4 ± 0.2 |
| 5-(Phenyl)-γ-hydroxyvaleric acid-O-sulphate | 6.6 ± 1.6 | 6.7 ± 2.6 | 8.4 ± 2.3 | 7.7 ± 2.0 |
| Isoferulic acid-4-O-sulfate | 131.2 ± 25.9 * | 149.2 ± 34.0 * | 137.4 ± 45.8 | 71.8 ± 29.5 |
| Σof Structurally Related Metabolites(Day 7, 8–24 h) | SHR1 | SHR5 | SHR6 | Control |
| (Epi)catechin-O-glucuronide | 1.6 ± 0.4 | 1.6 ± 0.1 | 2.2 ± 0.4 * | 1.1 ± 0.3 |
| O-Methyl-(epi)catechin-O-glucuronide | 8.1 ± 1.8 | 6.2 ± 1.3 | 5.2 ± 0.8 * | 3.7 ± 0.8 |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-glucuronide | 0.5 ± 0.1 | 0.7 ± 0.3 | 0.7 ± 0.1 * | 0.4 ± 0.1 |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-sulphate | 0.8 ± 0.1 | 1.4 ± 0.5 | 1.6 ± 0.3 | 1.1 ± 0.3 |
| 5-(Hydroxyphenyl)-γ-hydroxyvaleric acid-O-sulphate | 1.2 ± 0.4 | 1.8 ± 0.6 | 1.4 ± 0.3 | 1.1 ± 0.3 |
| 5-(Phenyl)-γ-hydroxyvaleric acid-O-sulphate | 13.2 ± 4.2 | 15.4 ± 3.6 | 14.5 ± 4.0 | 12.4 ± 2.8 |
| Isoferulic acid-4-O-sulfate | 209.4 ± 38.6 * | 218.2 ± 60.6 * | 171.5 ± 43.0 | 121.2 ± 20.8 |
| Percentage of Intake Day 1 (%) | Percentage of Intake Day 7 (%) | |||||
|---|---|---|---|---|---|---|
| SHR1 a | SHR5 a | SHR6 a | SHR1 a | SHR5 a | SHR6 a | |
| 0–8 h | 0.83 | 0.83 | 0.51 | 0.94 | 2.10 | 1.03 |
| 8–24 h | 0.19 | 0.74 | 0.51 | 0.58 | 2.31 | 0.39 |
| Total 24 h | 1.02 | 1.57 | 1.02 | 1.52 | 4.41 | 1.43 |
| Metabolites | SHR1 | SHR5 | SHR6 |
|---|---|---|---|
| 5-(Hydroxyphenyl)-γ-valerolactone-O-glucuronide 1 | 0.62 ± 0.06 * | 0.71 ± 0.02 | 0.46 ± 0.01 |
| (Epi)catechin-O-glucuronide 1 | Nd | Nd | 0.25 ± 0.02 * |
| (Epi)catechin-O-glucuronide 2 | 0.34 ± 0.01 * | 0.32 ± 0.03 | 0.39 ± 0.03 * |
| O-Methyl-(epi)catechin-O-glucuronide 1 | 0.24 ± 0.02 * | 0.17 ± 0.02 * | 0.23 ± 0.01 * |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-glucuronide 2 | 0.26 ± 0.01 * | 0.31 ± 0.00 * | 0.22 ± 0.01 * |
| Methyl-(epi)catechin-O-glucuronide 2 | 0.62 ± 0.05 * | 0.41 ± 0.00 | 0.71 ± 0.04 * |
| Methyl-(epi)catechin-O-glucuronide 3 | Nd | Nd | 0.37 ± 0.01 * |
| Di-methyl-(epi)catechin-O-glucuronide 1 | 0.54 ± 0.06 * | 0.86 ± 0.03 * | 0.50 ± 0.02 * |
| 5-(Hydroxyphenyl)-4-hydroxyvaleric acid-O-sulphate 2 | Nd | Nd | Nd |
| Di-methyl-(epi)catechin-O-glucuronide 2 | 0.42 ± 0.04 * | 0.35 ± 0.04 * | 0.19 ± 0.01 * |
| 5-(Hydroxyphenyl)-γ-valerolactone sulphate 1 | 0.42 ± 0.04 * | 0.31 ± 0.05 * | 0.42 ± 0.06 * |
| Mean Tissue Metabolites Levels (Day 7, after 4 h) | SHR1 | SHR5 | SHR6 | |
|---|---|---|---|---|
| Heart | 5-(Hydroxyphenyl)-γ-valerolactone-O-glucuronide | 2.50 ± 0.25 * | 2.64 ± 0.03 * | 2.27 ± 0.08 * |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-sulphate | 0.11 ± 0.00 * | 0.11 ± 0.00 * | Nd | |
| Kidneys | 5-(Hydroxyphenyl)-γ-valerolactone-O-glucuronide | 5.55 ± 0.11 | 6.62 ± 0.08 | 5.23 ± 0.34 |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-sulphate | 0.38 ± 0.00 * | 0.37 ± 0.02 * | 0.45 ± 0.03 * | |
| Liver | 5-(Hydroxyphenyl)-γ-valerolactone-O-glucuronide | 167.19 ± 8.77 | 209.98 ± 12.20 * | 179.26 ± 0.25 |
| 5-(Hydroxyphenyl)-γ-valerolactone-O-sulphate | Nd | Nd | Nd |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasines-Perea, Z.; Ky, I.; Cros, G.; Crozier, A.; Teissedre, P.-L. Grape Pomace: Antioxidant Activity, Potential Effect Against Hypertension and Metabolites Characterization after Intake. Diseases 2018, 6, 60. https://doi.org/10.3390/diseases6030060
Rasines-Perea Z, Ky I, Cros G, Crozier A, Teissedre P-L. Grape Pomace: Antioxidant Activity, Potential Effect Against Hypertension and Metabolites Characterization after Intake. Diseases. 2018; 6(3):60. https://doi.org/10.3390/diseases6030060
Chicago/Turabian StyleRasines-Perea, Zuriñe, Isabelle Ky, Gérard Cros, Alan Crozier, and Pierre-Louis Teissedre. 2018. "Grape Pomace: Antioxidant Activity, Potential Effect Against Hypertension and Metabolites Characterization after Intake" Diseases 6, no. 3: 60. https://doi.org/10.3390/diseases6030060
APA StyleRasines-Perea, Z., Ky, I., Cros, G., Crozier, A., & Teissedre, P.-L. (2018). Grape Pomace: Antioxidant Activity, Potential Effect Against Hypertension and Metabolites Characterization after Intake. Diseases, 6(3), 60. https://doi.org/10.3390/diseases6030060