
Illuminating Histidine-Deficient Intracellular Environments: A Novel Whole-Cell Microbial Fluorescence Sensor

Abstract
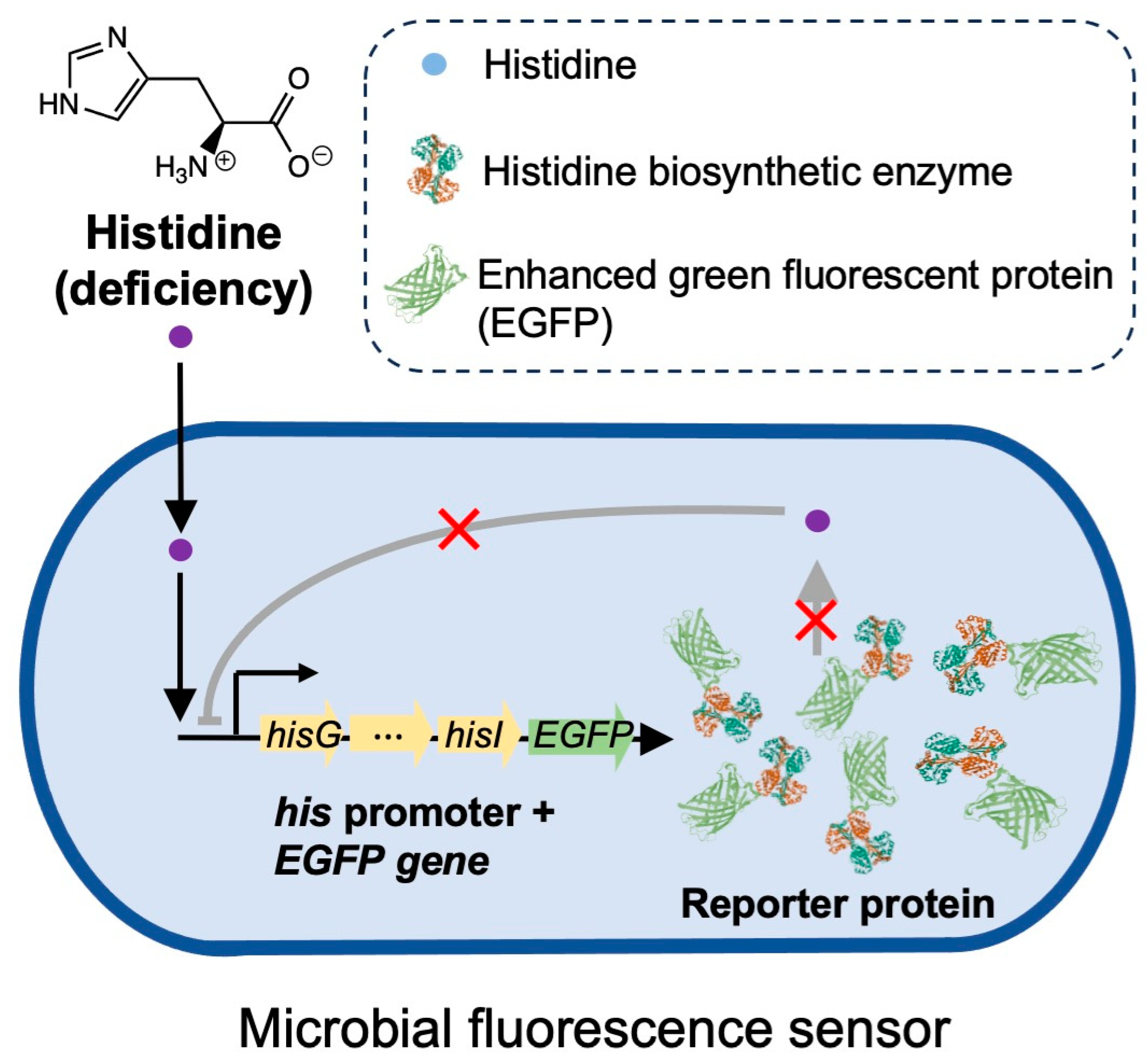
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacteria Cultivation
2.3. Acquisition of Target DNA Fragments by PCR
2.4. Homologous Recombination
2.5. Electrotransformation and Biosensor Construction
2.6. Quantification of EGFP in Bacterial Sensors
2.7. Cell Culture and Bacterial Infection
2.8. Fluorescence Microscopy Imaging
3. Results and Discussion
3.1. Construction of a Bacterial Fluorescence Sensor Responsive to Histidine Deficiency
3.2. Sensitivity and Specificity of the Bacterial Fluorescence Sensors in Response to Changes in Histidine Concentration
3.3. Application of the Bacterial Fluorescence Sensor to Real-Time Monitoring of Histidine Levels within Live Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sundberg, R.J.; Martin, R.B. Interactions of histidine and other imidazole derivatives with transition-metal ions in chemical and biological-systems. Chem. Rev. 1974, 74, 471–517. [Google Scholar] [CrossRef]
- Haas, H.; Panula, P. The role of histamine and the tuberomamillary nucleus in the nervous system. Nat. Rev. Neurosci. 2003, 4, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Ghadimi, H.; Partington, M.W.; Hunter, A. A familial disturbance of histidine metabolism. N. Engl. J. Med. 1961, 265, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, D.J.; Gluzman, I.Y.; Goldberg, D.E. Plasmodium hemozoin formation mediated by histidine-rich proteins. Science 1996, 271, 219–222. [Google Scholar] [CrossRef]
- Engesser, L.; Kluft, C.; Briet, E.; Brommer, E.J.P. Familial elevation of plasma histidine-rich glycoprotein in a family with thrombophilia. Brit. J. Haematol. 1987, 67, 355–358. [Google Scholar] [CrossRef]
- Gerber, D.A. Low free serum histidine concentration in rheumatoid-arthritis—Measure of disease activity. J. Clin. Investig. 1975, 55, 1164–1173. [Google Scholar] [CrossRef]
- Rao, M.L.; Stefan, H.; Scheid, C.; Kuttler, A.D.S.; Froscher, W. Serum amino-acids, liver status, and antiepileptic drug-therapy in epilepsy. Epilepsia 1993, 34, 347–354. [Google Scholar] [CrossRef]
- Watanabe, M.; Suliman, M.E.; Qureshi, A.R.; Garcia-Lopez, E.; Barany, P.; Heimburger, O.; Stenvinkel, P.; Lindholm, B. Consequences of low plasma histidine in chronic kidney disease patients: Associations with inflammation, oxidative stress, and mortality. Am. J. Clin. Nutr. 2008, 87, 1860–1866. [Google Scholar] [CrossRef]
- Saito, H.; Goodnough, L.T.; Boyle, J.M.; Heimburger, N. Reduced histidine-rich glycoprotein levels in plasma of patients with advanced liver-cirrhosis—Possible implications for enhanced fibrinolysis. Am. J. Med. 1982, 73, 179–182. [Google Scholar] [CrossRef]
- Schmid, G.; Fricke, L.; Lange, H.W.; Heidland, A. Intracellular histidine content of various tissues (brain, striated-muscle and liver) in experimental chronic renal-failure. Klin. Wschr. 1977, 55, 583–585. [Google Scholar] [CrossRef]
- Tateda, N.; Matsuhisa, K.; Hasebe, K.; Kitajima, N.; Miura, T. High-performance liquid chromatographic method for rapid and highly sensitive determination of histidine using postcolumn fluorescence detection with o-phthaldialdehyde. J. Chromatogr. B. 1998, 718, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, M.; Nakazawa, T. Determination of pK(a) values of individual histidine residues in proteins using mass spectrometry. Anal. Chem. 2008, 80, 6481–6487. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.B.; Kumar, D.; Madhuri, R.; Tiwari, M.P. Metal ion mediated imprinting for electrochemical enantioselective sensing of L-histidine at trace level. Biosens. Bioelectron. 2011, 28, 117–126. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Sun, M.X. Determination of histamine and histidine by capillary zone electrophoresis with pre-column naphthalene-2,3-dicarboxaldehyde derivatization and fluorescence detection. J. Chromatogr. A 2004, 1040, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Un, H.I.; Wu, S.; Huang, C.B.; Xu, Z.; Xu, L. A naphthalimide-based fluorescent probe for highly selective detection of histidine in aqueous solution and its application in in vivo imaging. Chem. Commun. 2015, 51, 3143–3146. [Google Scholar] [CrossRef]
- Zhu, X.H.; Zhao, T.B.; Nie, Z.; Miao, Z.; Liu, Y.; Yao, S.Z. Nitrogen-doped carbon nanoparticle modulated turn-on fluorescent probes for histidine detection and its imaging in living cells. Nanoscale 2016, 8, 2205–2211. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Abed, M.A.; Lee, N.B.; Wu, X.; Du, G.; Pu, L. Determining the concentration and enantiomeric composition of histidine using one fluorescent probe. Chem. Commun. 2021, 57, 587–590. [Google Scholar] [CrossRef]
- Hu, H.; Gu, Y.; Xu, L.; Zou, Y.; Wang, A.; Tao, R.; Chen, X.; Zhao, Y.; Yang, Y. A genetically encoded toolkit for tracking live-cell histidine dynamics in space and time. Sci. Rep. 2017, 7, 43479. [Google Scholar] [CrossRef]
- Della Corte, D.; van Beek, H.L.; Syberg, F.; Schallmey, M.; Tobola, F.; Cormann, K.U.; Schlicker, C.; Baumann, P.T.; Krumbach, K.; Sokolowsky, S.; et al. Engineering and application of a biosensor with focused ligand specificity. Nat. Commun. 2020, 11, 4851. [Google Scholar] [CrossRef]
- Gonzalez-Escobedo, G.; Marshall, J.M.; Gunn, J.S. Chronic and acute infection of the gall bladder by Salmonella Typhi: Understanding the carrier state. Nat. Rev. Microbiol. 2011, 9, 9–14. [Google Scholar] [CrossRef]
- Mastroeni, P.; Grant, A.; Restif, O.; Maskell, D. A dynamic view of the spread and intracellular distribution of Salmonella enterica. Nat. Rev. Microbiol. 2009, 7, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.E.; Ramos-Montanez, S. Biosynthesis of Histidine. EcoSal. Plus 2009, 3, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Johnston, H.M.; Barnes, W.M.; Chumley, F.G.; Bossi, L.; Roth, J.R. Model for regulation of the histidine operon of salmonella. Proc. Nat. Acad. Sci. India Sect. B-Biol. Sci. 1980, 77, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Kasai, T. Regulation of expression of histidine operon in salmonella-typhimurium. Nature 1974, 249, 523–527. [Google Scholar] [CrossRef]
- Lewis, J.A.; Ames, B.N. Histidine regulation in salmonella-typhimurium.11. percentage of transfer rnahis charges in-vivo and its relation to repression of histidine operon. J. Mol. Biol. 1972, 66, 31–42. [Google Scholar]
- Henry, T.; Garcia-del Portillo, F.; Gorvel, J.P. Identification of Salmonella functions critical for bacterial cell division within eukaryotic cells. Mol. Microbiol. 2005, 56, 252–267. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Y.; Fu, J.; Zhang, B.; Cheng, S.; Wu, M.; Wang, Z.; Jiang, J.; Chang, C.; Liu, X. Salmonella Proteomic Profiling during Infection Distinguishes the Intracellular Environment of Host Cells. Msystems 2019, 4, 10–1128. [Google Scholar] [CrossRef]
- Nagao, T.; Nakayama-Imaohji, H.; Elahi, M.; Tada, A.; Toyonaga, E.; Yamasaki, H.; Okazaki, K.; Miyoshi, H.; Tsuchiya, K.; Kuwahara, T. L histidine augments the oxidative damage against Gram negative bacteria by hydrogen peroxide. Int. J. Mol. Med. 2018, 41, 2847–2854. [Google Scholar] [CrossRef]
- Gomes, T.A.; Zanette, C.M.; Spier, M.R. An overview of cell disruption methods for intracellular biomolecules recovery. Prep. Biochem. Biotechnol. 2020, 50, 635–654. [Google Scholar] [CrossRef]
- Radkov, A.D.; Moe, L.A. Bacterial synthesis of D-amino acids. Appl. Microbiol. Biotechnol. 2014, 98, 5363–5374. [Google Scholar] [CrossRef]
- Cava, F.; de Pedro, M.A.; Lam, H.; Davis, B.M.; Waldor, M.K. Distinct pathways for modification of the bacterial cell wall by non-canonical D-amino acids. EMBO J. 2011, 30, 3442–3453. [Google Scholar] [CrossRef] [PubMed]
- LaRue, T.A.; Spencer, J.F.T. The utilization of imidazoles by yeasts. Can. J. Microbiol. 2011, 13, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Ohl, M.E.; Miller, S.I. Salmonella: A model for bacterial pathogenesis. Annu. Rev. Med. 2001, 52, 259–274. [Google Scholar] [CrossRef]
- Monack, D.M.; Mueller, A.; Falkow, S. Persistent bacterial infections: The interface of the pathogen and the host immune system. Nat. Rev. Microbiol. 2004, 2, 747–765. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Moro, J.; Tomé, D.; Schmidely, P.; Demersay, T.-C.; Azzout-Marniche, D. Histidine: A Systematic Review on Metabolism and Physiological Effects in Human and Different Animal Species. Nutrients 2020, 12, 1414. [Google Scholar] [CrossRef]
- Mills, E.; Petersen, E. Quantification of Microbial Fluorescent Sensors During Live Intracellular Infections. In Bacterial Virulence. Methods in Molecular Biology; Gal-Mor, O., Ed.; Humana: New York, NY, USA, 2022; Volume 2427. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Medium | Components |
|---|---|
| LB | 10 g of tryptone, 5 g of yeast extract, 10 g of NaCl, 15 g of agar (only for LB plate), remaining volume filled to 1 L with ultrapure water, and sterilization at 121 °C for 20 min. |
| M9 | 2 mL 5 × M9 salt solution (Na2HPO4, KH2PO4, NaCl, NH4Cl), 20 μL 1 M MgSO4, 1 μL 1 M CaCl2, 200 μL 20% glucose, remaining volume filled to 10 mL with ultrapure water, and 121 °C sterilization 20 min. |
| SOC | 20 g of tryptone, 5 g of yeast extract, 0.5 g of NaCl, remaining volume filled to 1 L with ultrapure water, sterilized at 121 °C for 20 min, and 5 mL of 2 M MgCl2 aqueous solution added. |
| Primers | ) |
|---|---|
| pSB3313-homo-F | AGAACGCAGAAGCGGTCTGATAAAACAGAATTTG |
| pSB3313-homo-R | GTAGCATAGGGTTTGCAGAATCCCTGCTTC |
| pgex-homo-F | CCTCTGACACATGCAGCTCC |
| pgex-homo-R | GAATTTATGCGGTGTGAAATACCGCAC |
| pSB3313-his-F | TTCTGCAAACCCTATGCTACGCTTTCAATTCCACTC ATACG |
| pSB3313-egfp-R2 | TCAGACCGCTTCTGCGTTCTTTACTTGTACAGCTCGTCCATGC |
| pgex-his-F | ATTTCACACCGCATAAATTCGCTTTCAATTCCACTCATACG |
| pgex-egfp-R2 | GGAGCTGCATGTGTCAGAGGTTACTTGTACAGCTCGTCCATGC |
| R1-his | CCTCGCCCTTGCTCACCATTCTGCGTTCCTCTTTATCC |
| R1-egfp | GGATAAAGAGGAACGCAGAATGGTGAGCAAGGGCGAGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Li, Z.; Zhao, M. Illuminating Histidine-Deficient Intracellular Environments: A Novel Whole-Cell Microbial Fluorescence Sensor. Chemosensors 2023, 11, 515. https://doi.org/10.3390/chemosensors11100515
Li X, Li Z, Zhao M. Illuminating Histidine-Deficient Intracellular Environments: A Novel Whole-Cell Microbial Fluorescence Sensor. Chemosensors. 2023; 11(10):515. https://doi.org/10.3390/chemosensors11100515
Chicago/Turabian StyleLi, Xinyi, Zezhou Li, and Meiping Zhao. 2023. "Illuminating Histidine-Deficient Intracellular Environments: A Novel Whole-Cell Microbial Fluorescence Sensor" Chemosensors 11, no. 10: 515. https://doi.org/10.3390/chemosensors11100515