Challenges in Developing a Biochip for Intact Histamine Using Commercial Antibodies



Abstract
:1. Introduction
1.1. Antibodies against Derived Histamine
1.2. Antibodies against Intact Histamine
1.3. Commercial Anti-Histamine Antibodies and Their Use in Immunosensors and Tests
2. Materials and Methods
2.1. Reagents
2.2. Microarray (Biochip) Fabrication
2.2.1. Preparation of Protein-Histamine Conjugates
2.2.2. Spotting of the Probes
2.2.3. Histamine Immunoassays
2.2.4. Detection
3. Results
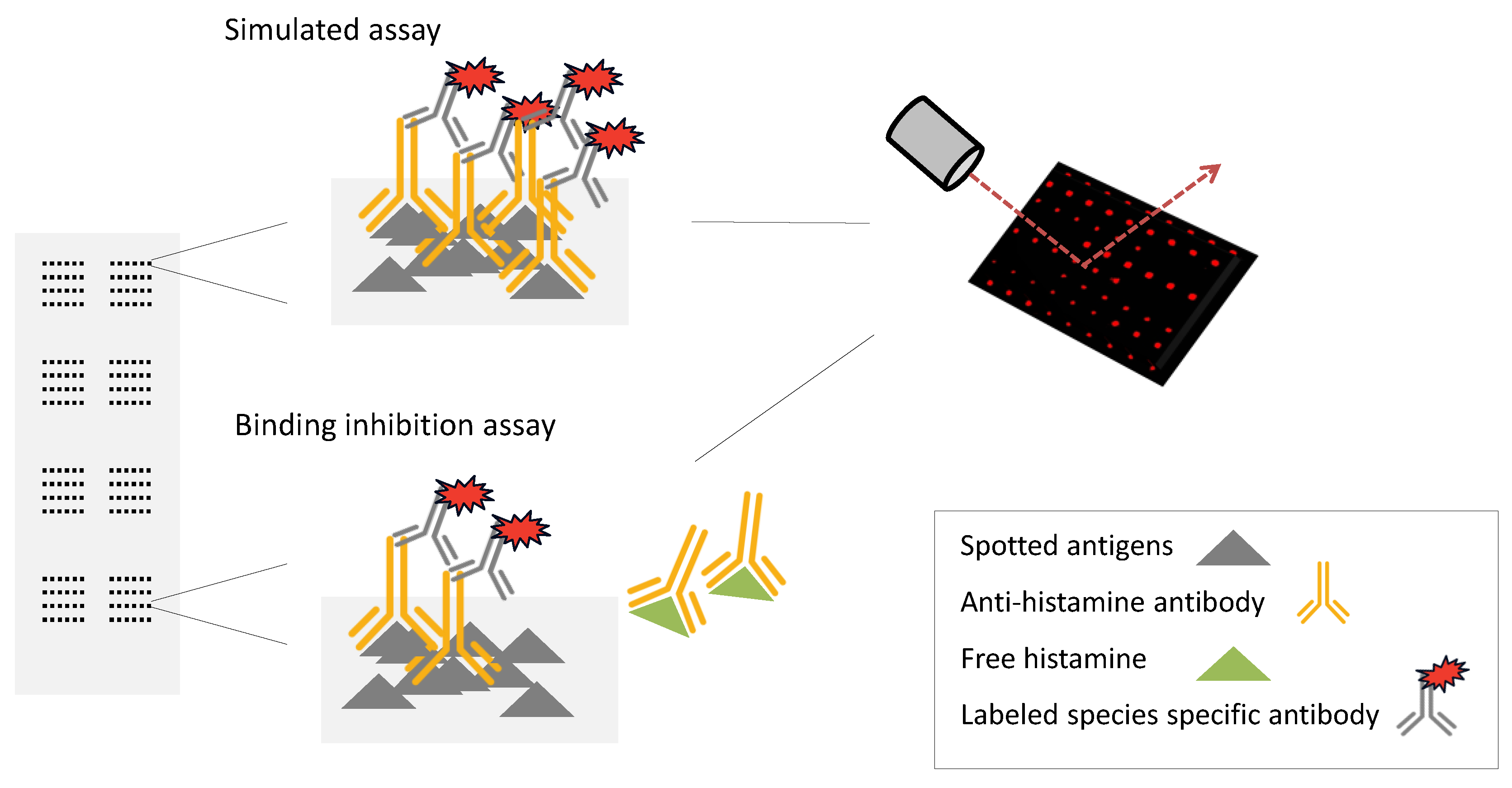
3.1. Assay Format
3.2. Choice of Antibodies
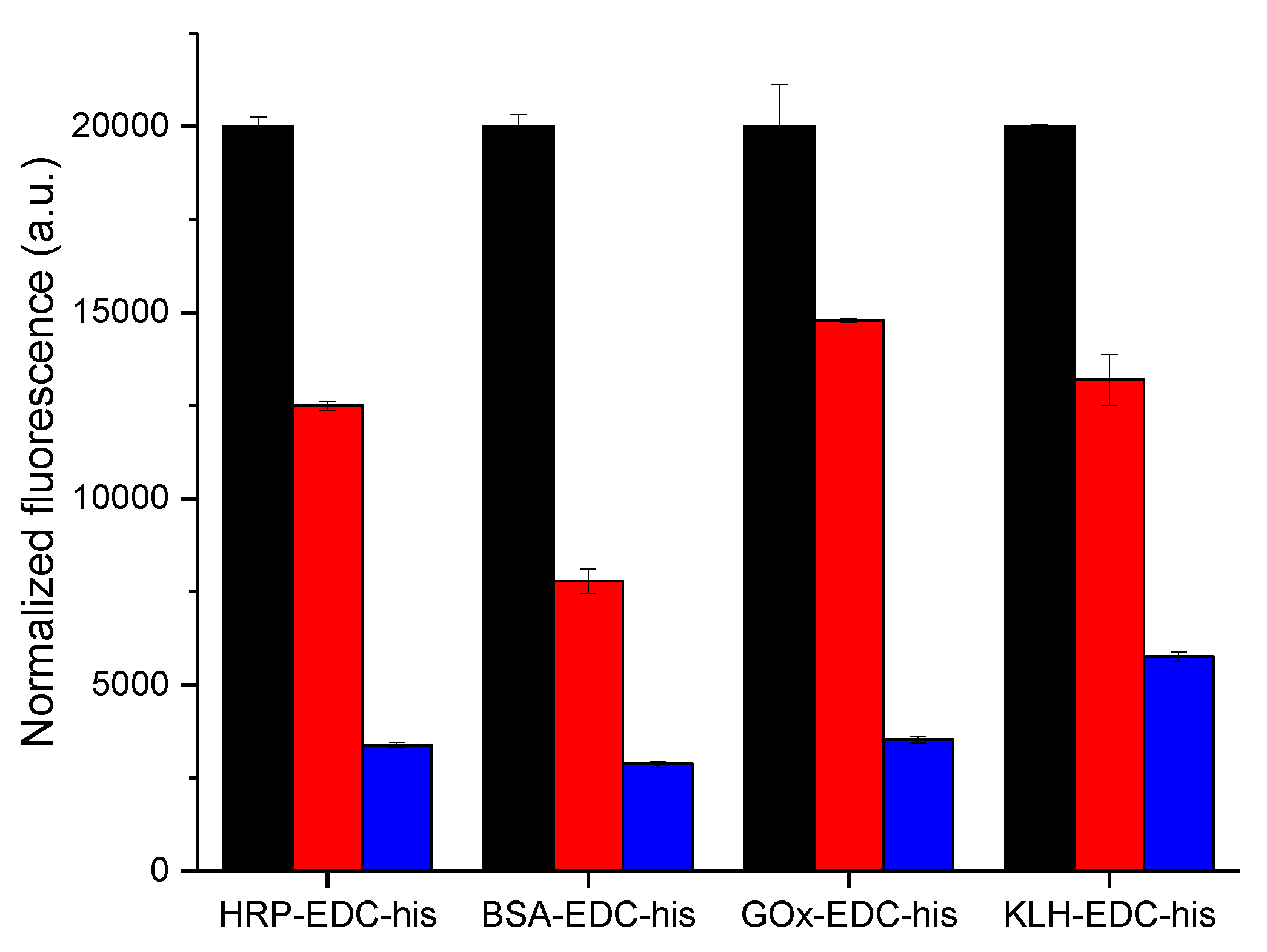
3.3. Specificity of the Antibody Binding in Simulated Assay
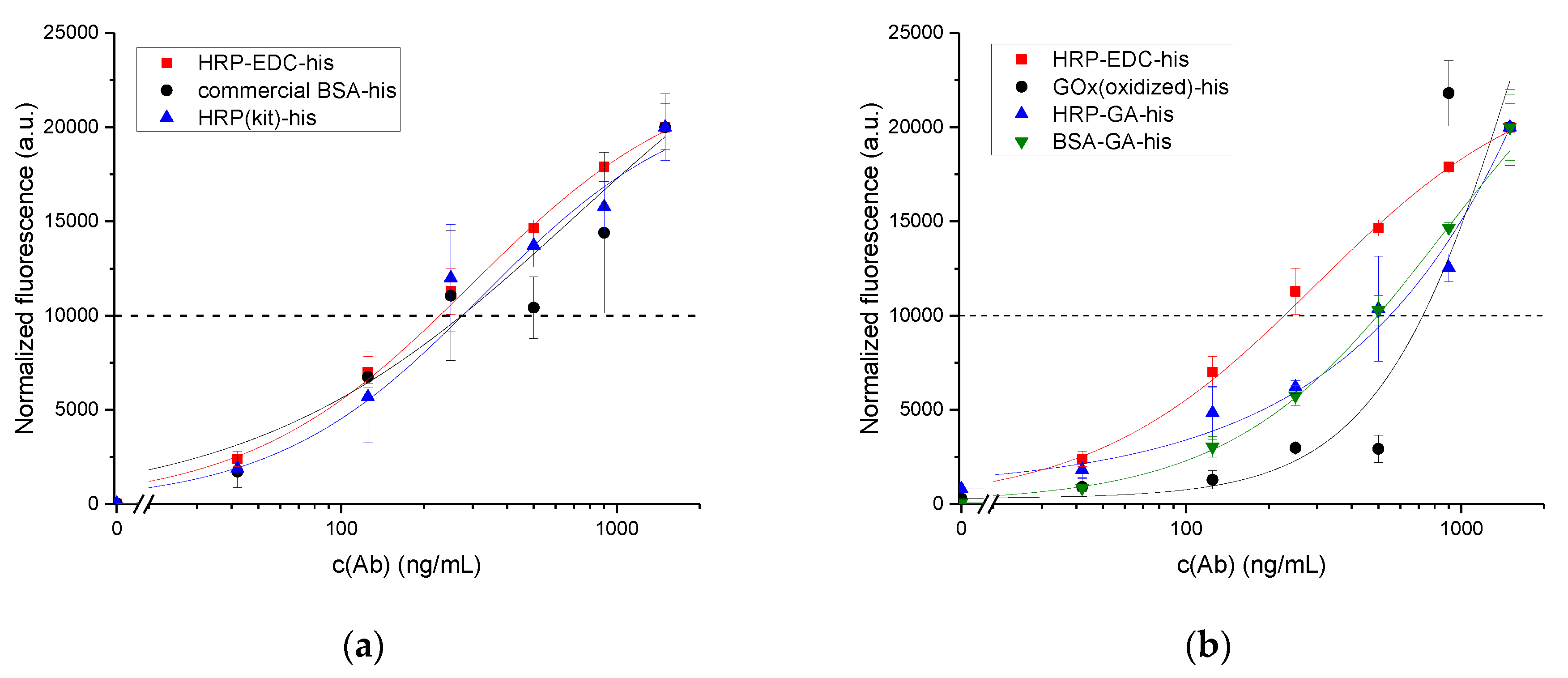
3.4. Binding Inhibition Assay Using Ab1
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Silla Santos, M.H. Biogenic amines: Their importance in foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]
- Food and Drug Administration. Fish and Fishery Products Hazards and Controls Guidance. Fourth edition—April 2011. Available online: http://www.fda.gov/FoodGuidances (accessed on 5 December 2017).
- Morel, A.; Darmon, M.; Delaage, M. Recognition of imidazole and histamine derivatives by monoclonal antibodies. Mol. Immunol. 1990, 27, 995–1000. [Google Scholar] [CrossRef]
- Morel, A.M.; Delaage, M.A. Immunoanalysis of histamine through a novel chemical derivatization. J. Allergy Clin. Immunol. 1988, 82, 646–654. [Google Scholar] [CrossRef]
- Guesdon, J.L.; Chevrier, D.; Mazie, J.C.; David, B.; Avrameas, S. Monoclonal anti-histamine antibody. Preparation, characterization and application to enzyme immunoassay of histamine. J. Immunol. Methods 1986, 87, 69–78. [Google Scholar] [CrossRef]
- Chevrier, D.; Guesdon, J.L.; Mazie, J.C.; Avrameas, S. Enzyme immunoassay for the measurement of histamine. J. Immunol. Methods 1986, 94, 119–125. [Google Scholar] [CrossRef]
- Rauch, P.; Rychetsky, P.; Hochel, I.; Bilek, R.; Guesdon, J. Enzyme immunoassay of histamine in foods. Food Agric. Immunol. 1992, 4, 67–72. [Google Scholar] [CrossRef]
- Serrar, D.; Brebant, R.; Bruneau, S.; Denoyel, G.A. The development of a monoclonal antibody-based ELISA for the determination of histamine in food: Application to fishery products and comparison with the HPLC assay. Food Chem. 1995, 54, 85–91. [Google Scholar] [CrossRef]
- Haydik, I.B. Histamine determination by radioimmunoassay (RIA) [Abstract]. J. Allergy Clin. Immunol. 1983, 71, 152. [Google Scholar] [CrossRef]
- Buckler, R.T.; Dailey, F.A.; Ficalora, J.A.; Gavin, J.J.; Plunkett, G.A. Histamine derivatives, immunogen conjugates and antibodies raised thereto. Patent US5112738, 12 May 1992. [Google Scholar]
- Panula, P.; Yang, H.; Costa, E. Histamine-containing neurons in the rat hypothalamus. Proc. Natl. Acad. Sci. USA 1984, 81, 2572–2576. [Google Scholar] [CrossRef] [PubMed]
- Dohlsten, M.; Hallberg, T.; Håkanson, R. Production and characterization of rabbit antibodies against histamine. Mol. Immunol. 1986, 23, 799–806. [Google Scholar] [CrossRef]
- Mita, H.; Yasueda, H.; Shida, T.; Baba, S. An attempt to produce an antibody of histamine and histamine derivatives. Agents Actions 1984, 14, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, B.J.; Seybold, V.S. Localization of neuronal histamine in rat brain. Neurosci. Lett. 1982, 29, 105–110. [Google Scholar] [CrossRef]
- Johansson, O.; Virtanen, M.; Hilliges, M.; Yang, Q. Histamine immunohistochemistry: A new and highly sensitive method for studying cutaneous mast cells. Histochem. J. 1992, 24, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Schneider, E.; Usleber, E.; Märtlbauer, E. Production and characterization of antibodies against histamine. In Immunoassays for Residue Analysis; ACS Publications: Washington, DC, USA, 1996; pp. 413–420. ISBN 9780841233799. [Google Scholar]
- Aygün, O.; Schneider, E.; Scheuer, R.; Usleber, E.; Gareis, M.; Märtlbauer, E. Comparison of ELISA and HPLC for the determination of histamine in cheese. J. Agric. Food Chem. 1999, 47, 1961–1964. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Xu, Z.-L.; Yang, J.-Y.; Xiao, Z.-L.; Li, Y.-J.; Beier, R.C.; Sun, Y.-M.; Lei, H.-T.; Wang, H.; Shen, Y.-D. Synthesis of novel haptens and development of an enzyme-linked immunosorbent assay for quantification of histamine in foods. J. Agric. Food Chem. 2014, 62, 12299–12308. [Google Scholar] [CrossRef] [PubMed]
- Baker, M. Blame It on the antibodies. Nature 2015, 521, 274–276. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.G. Quality Issues of Research Antibodies. Anal. Chem. Insights 2016, 11, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Baker, M. Antibody anarchy: A call to order. Nature 2015, 527, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Beermann, S.; Seifert, R.; Neumann, D. Commercially available antibodies against human and murine histamine H4-receptor lack specificity. Naunyn. Schmiedebergs. Arch. Pharmacol. 2012, 385, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B. An open letter to our readers on the use of antibodies. J. Comp. Neurol. 2005, 493, 477–478. [Google Scholar] [CrossRef] [PubMed]
- Pradidarcheep, W.; Labruyère, W.T.; Dabhoiwala, N.F.; Lamers, W.H. Lack of specificity of commercially available antisera: Better specifications needed. J. Histochem. Cytochem. 2008, 56, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, A.; Plückthun, A. Reproducibility: Standardize antibodies used in research. Nature 2015, 518, 27–29. [Google Scholar] [CrossRef] [PubMed]
- O’Kennedy, R.; Fitzgerald, S.; Murphy, C. Don’t blame it all on antibodies—The need for exhaustive characterisation, appropriate handling, and addressing the issues that affect specificity. TrAC Trends Anal. Chem. 2017, 89, 53–59. [Google Scholar] [CrossRef]
- Bordeaux, J.; Welsh, A.; Agarwal, S.; Killiam, E.; Baquero, M.; Hanna, J.; Anagnostou, V.; Rimm, D. Antibody validation. Biotechniques 2010, 48, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Roncador, G.; Engel, P.; Maestre, L.; Anderson, A.P.; Cordell, J.L.; Cragg, M.S.; Šerbec, V.Č.; Jones, M.; Lisnic, V.J.; Kremer, L.; Li, D.; Koch-Nolte, F.; Pascual, N.; Rodríguez-Barbosa, J.-I.; Torensma, R.; Turley, H.; Pulford, K.; Banham, A.H. The European antibody network’s practical guide to finding and validating suitable antibodies for research. MAbs 2016, 8, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Voskuil, J.L.A. The challenges with the validation of research antibodies. F1000Research 2017, 6, 161. [Google Scholar] [CrossRef] [PubMed]
- Vasilevsky, N.A.; Brush, M.H.; Paddock, H.; Ponting, L.; Tripathy, S.J.; LaRocca, G.M.; Haendel, M.A. On the reproducibility of science: Unique identification of research resources in the biomedical literature. PeerJ. 2013, 1, e148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Kobayashi, M.; Furui, K.; Soh, N.; Nakano, K.; Imato, T. Surface plasmon resonance immunosensor for histamine based on an indirect competitive immunoreaction. Anal. Chim. Acta 2006, 576, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Adányi, N.; Székács, I.; Szendro, I.; Székács, A. Determination of histamine content in vegetable juices by using direct and competitive immunosensors. Food Agric. Immunol. 2014, 25, 37–41. [Google Scholar] [CrossRef]
- Lim, T.K.; Ohta, H.; Matsunaga, T. Microfabricated on-chip-type electrochemical flow immunoassay system for the detection of histamine released in whole blood samples. Anal. Chem. 2003, 75, 3316–3321. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, J.; Chen, X. Competitive electrochemical immunosensor for the detection of histamine based on horseradish peroxidase initiated deposition of insulating film. J. Electroanal. Chem. 2015, 736, 88–92. [Google Scholar] [CrossRef]
- Köse, S.; Kaklikkaya, N.; Koral, S.; Tufan, B.; Buruk, K.C.; Aydin, F. Commercial test kits and the determination of histamine in traditional (ethnic) fish products-evaluation against an EU accepted HPLC method. Food Chem. 2011, 125, 1490–1497. [Google Scholar] [CrossRef]
- Rogers, P.L.; Staruszkiewicz, W.F. Histamine Test Kit Comparison. J. Aquat. Food Prod. Technol. 2000, 9, 5–17. [Google Scholar] [CrossRef]
- Gogalic, S.; Sauer, U.; Doppler, S.; Heinzel, A.; Perco, P.; Lukas, A.; Simpson, G.; Pandha, H.; Horvath, A.; Preininger, C. Validation of a protein panel for the non-invasive detection of recurrent non-muscle invasive bladder cancer. Biomarkers 2017, 19, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Buchegger, P.; Preininger, C. Four Assay Designs and On-Chip Calibration: Gadgets for a Sepsis Protein Array. Anal. Chem. 2014, 86, 3174–3180. [Google Scholar] [CrossRef] [PubMed]
- Poller, A.M.; Crooks, S.; Preininger, C. Influence of different surface chemistries on the ultrasensitive on-chip detection of enrofloxacin in milk. Sens. Actuators B Chem. 2015, 209, 1077–1083. [Google Scholar] [CrossRef]
- The Antibody Registry. Available online: http://antibodyregistry.org/ (accessed on 14 October 2017).
- Strasser, A.; Dietrich, R.; Usleber, E.; Märtlbauer, E. Immunochemical rapid test for multiresidue analysis of antimicrobial drugs in milk using monoclonal antibodies and hapten-glucose oxidase conjugates. Anal. Chim. Acta 2003, 495, 11–19. [Google Scholar] [CrossRef]
- Fujiwara, K.; Kitagawa, T.; Inoue, Y.; Alonso, G. Monoclonal antibodies against glutaraldehyde-conjugated histamine: Application to immunocytochemistry. Histochem. Cell Biol. 1997, 107, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Romero-Calderón, R.; Uhlenbrock, G.; Borycz, J.; Simon, A.F.; Grygoruk, A.; Yee, S.K.; Shyer, A.; Ackerson, L.C.; Maidment, N.T.; Meinertzhagen, I.A.; et al. A glial variant of the vesicular monoamine transporter is required to store histamine in the Drosophila visual system. PLoS Genet. 2008, 4. [Google Scholar] [CrossRef]
- McCoole, M.D.; Baer, K.N.; Christie, A.E. Histaminergic signaling in the central nervous system of Daphnia and a role for it in the control of phototactic behavior. J. Exp. Biol. 2011, 214, 1773–1782. [Google Scholar] [CrossRef] [PubMed]
- Borycz, J.; Borycz, J.A.; Loubani, M.; Meinertzhagen, I.A. tan and ebony genes regulate a novel pathway for transmitter metabolism at fly photoreceptor terminals. J. Neurosci. 2002, 22, 10549–10557. [Google Scholar] [PubMed]
- Wortberg, M.; Goodrow, M.H.; Gee, S.J.; Hammock, B.D. Immunoassay for Simazine and Atrazine with Low Cross-Reactivity for Propazine. J. Agric. Food Chem. 1996, 44, 2210–2219. [Google Scholar] [CrossRef]
- Holthues, H.; Pfeifer-Fukumura, U.; Sound, I.; Baumann, W. Evaluation of the concept of heterology in a monoclonal antibody-based ELISA utilizing direct hapten linkage to polystyrene microtiter plates. J. Immunol. Methods 2005, 304, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Goodrow, M.H.; Hammock, B.D.; Harrison, R.O. Hapten synthesis, antibody development, and competitive inhibition enzyme immunoassay for s-triazine herbicides. J. Agric. Food Chem. 1990, 38, 990–996. [Google Scholar] [CrossRef]
- Rizner, T.L.; Sasano, H.; Choi, M.H.; Odermatt, A.; Adamski, J. Recommendations for description and validation of antibodies for research use. J. Steroid Biochem. Mol. Biol. 2016, 156, 40–42. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Bandrowski, A.; Carr, S.; Edwards, A.; Ellenberg, J.; Lundberg, E.; Rimm, D.L.; Rodriguez, H.; Hiltke, T.; Snyder, M.; Yamamoto, T. A proposal for validation of antibodies. Nat. Methods 2016, 13. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Retailer | Catalog Number | Lot/Batch | Host | Clonality | Immunogen | Purity | Cross-Reactivity/Specificity | Application | RRID [40] |
|---|---|---|---|---|---|---|---|---|---|---|
| Ab1 | Sigma Aldrich | H7403 | 081M4794, 51K4855 | rabbit | pAb | histamine conjugated to succinylated KLH | affinity isolated antibody | stains histamine-containing cells in fixed sections of rat endocrine cells and mast cells | dot blot, IHC | AB_260077 |
| Ab2 | ImmunoStar | 22939 | 1006001 | rabbit | pAb | synthetic histamine coupled to succinylated KLH with carbodiimide linker | whole serum | staining blocked by pre-absorption of the antiserum with histamine conjugate no cross-reactivity with L-histidine | IHC | AB_572245 |
| Ab3 | Thermo Fischer Scientific | PA1-85547 | OD1701064 | rabbit | pAb | synthetic peptide corresponding to part of the native molecule conjugated to BSA | Protein G purified | detects histamine in many different sample types | ELISA | AB_931244 |
| Ab4 | Biozol Diagnostica Vertried/AbD Serotec | 4970-0010 | 0812 | rabbit | pAb | histamine conjugated to cationised BSA using a 1,4-benzoquinone | affinity chromatography on protein G | specific for histamine | ELISA | AB_619446 |
| Ab5 | Bertin Pharma | A03890 | 0116 | mouse | mAb | histamine-NH-CO-(NH2)6-S-maleinido-BSA | - | histamine 100% 3-methyl histamine, 1-methyl histamine, histidine, serotonin <0.04% | EIA | |
| Ab6 | Merck Millipore | MAB5408 | 2522068 | mouse | mAb | histamine-hexamethylene diisocyanate (HD)-BSA | purified | histamine-HD-BSA 1 1-methyl-histamine-HD-BSA 1 histamine-GA-BSA, histidine-HD-BSA 1/>50 000 | IHC, ICC | AB_177651 |
| Ab2 | Ab3 | Ab4 | Ab5 | Ab6 | Ab1 | Dynamic Range (µg/mL) | IC50 (µg/mL) | |
|---|---|---|---|---|---|---|---|---|
| carbodiimide coupling chemistry | ||||||||
| BSA-EDC-his | + | + | + | – | – | + | 13–106 | 37 |
| HRP-EDC-his | + | – | – | – | – | + | 8–111 | 28 |
| KLH-EDC-his | + | – | – | – | – | + | 11–103 | 48 |
| GOx-EDC-his | + | n/a | n/a | – | n/a | + | 16–98 | 37 |
| pure proteins | ||||||||
| BSA pure | + | – | – | n/a | n/a | – | ||
| HRP pure | + | n/a | n/a | n/a | n/a | – | ||
| KLH pure | + | n/a | n/a | n/a | n/a | – | ||
| GOx pure | – | n/a | n/a | n/a | n/a | – | ||
| aldehyde-based coupling | ||||||||
| HRP(kit)-histamine | – | – | – | – | – | + | 4–89 | 20 |
| GOx(oxidized)-his (acc. Strasser) | – | n/a | n/a | n/a | n/a | + | ||
| HRP(oxidized)-his (acc. Schneider) | n/a | n/a | n/a | n/a | n/a | – | ||
| glutaraldehyde as a cross-linker | ||||||||
| BSA-GA-his | – | n/a | n/a | n/a | n/a | + | 28–191 | 70 |
| HRP-GA-his | n/a | n/a | n/a | n/a | n/a | + | 37–336 | 85 |
| commercial BSA-his conjugate | – | – | + | – | – | + | 4–38 | 14 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattsson, L.; Doppler, S.; Preininger, C. Challenges in Developing a Biochip for Intact Histamine Using Commercial Antibodies. Chemosensors 2017, 5, 33. https://doi.org/10.3390/chemosensors5040033
Mattsson L, Doppler S, Preininger C. Challenges in Developing a Biochip for Intact Histamine Using Commercial Antibodies. Chemosensors. 2017; 5(4):33. https://doi.org/10.3390/chemosensors5040033
Chicago/Turabian StyleMattsson, Leena, Sara Doppler, and Claudia Preininger. 2017. "Challenges in Developing a Biochip for Intact Histamine Using Commercial Antibodies" Chemosensors 5, no. 4: 33. https://doi.org/10.3390/chemosensors5040033
APA StyleMattsson, L., Doppler, S., & Preininger, C. (2017). Challenges in Developing a Biochip for Intact Histamine Using Commercial Antibodies. Chemosensors, 5(4), 33. https://doi.org/10.3390/chemosensors5040033