Season of Birth Impacts the Neonatal Nasopharyngeal Microbiota

 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Methods
2.1. Study Population
2.2. Ethics Statement
2.3. Collection of Nasopharyngeal Samples at 1-Month of Age
2.4. DNA Extraction from Nasopharyngeal Specimens for T-RFLP
2.5. T-RFLP on Nasopharyngeal Specimens from Infants
2.6. Identification of Bacterial Groups with Given TRFs
2.7. Perinatal Risk Factors
2.8. Statistical Analysis
3. Results
3.1. Characteristics of Study Population
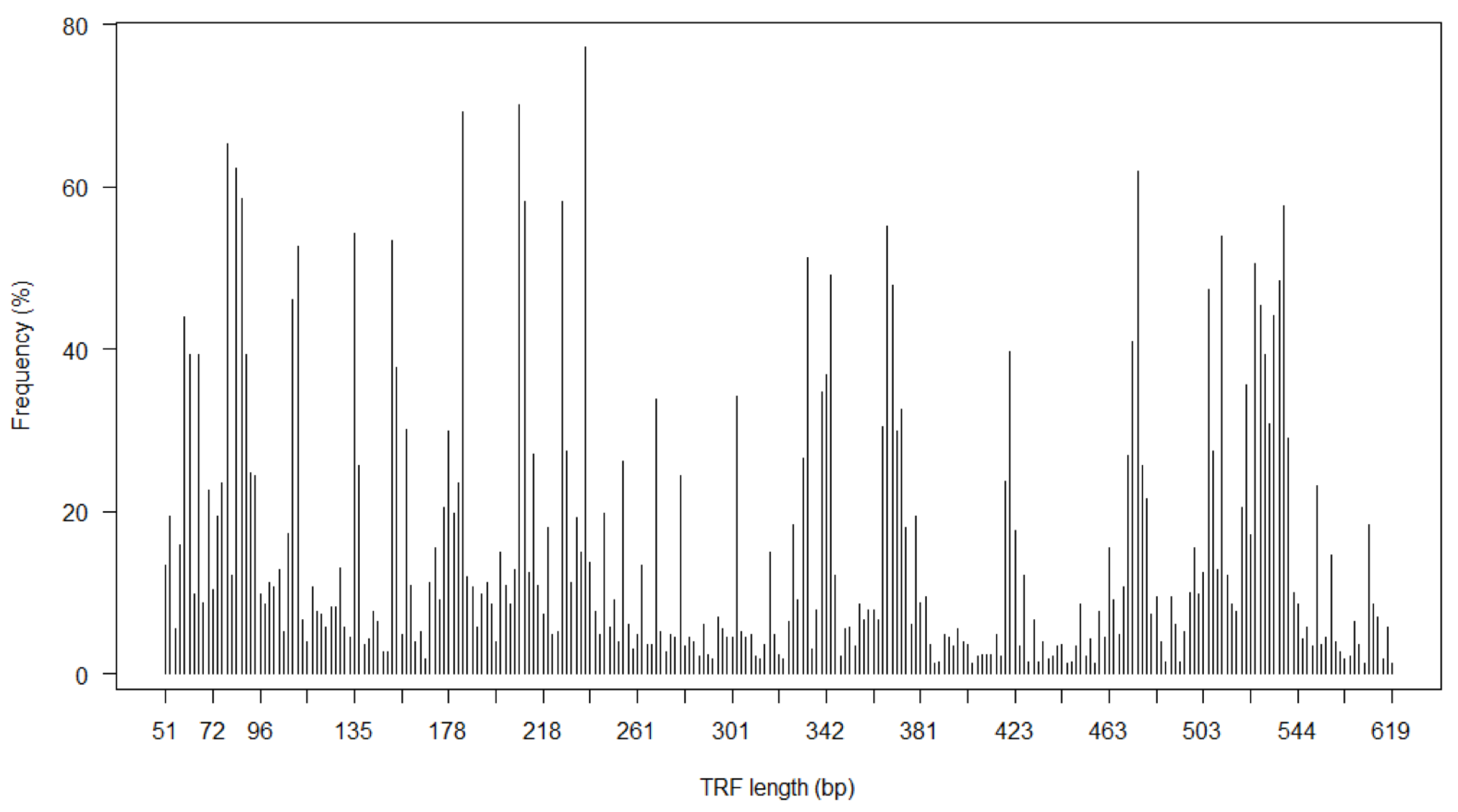
3.2. Nasopharyngeal Bacterial Richness in Asymptomatic Neonates
3.3. Predictors of Neonatal Nasopharyngeal Bacterial Richness
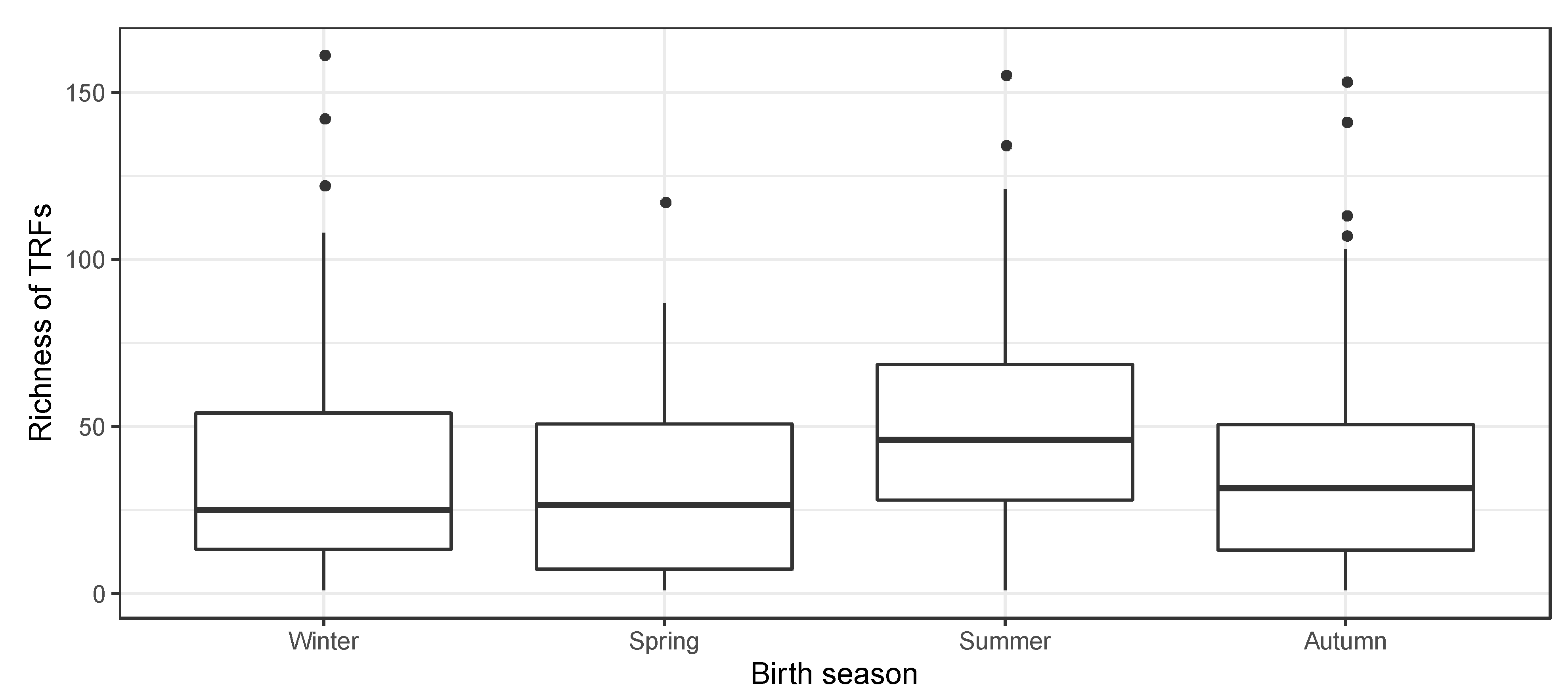
3.4. Season of Birth and Nasopharyngeal Bacterial Richness in Neonates
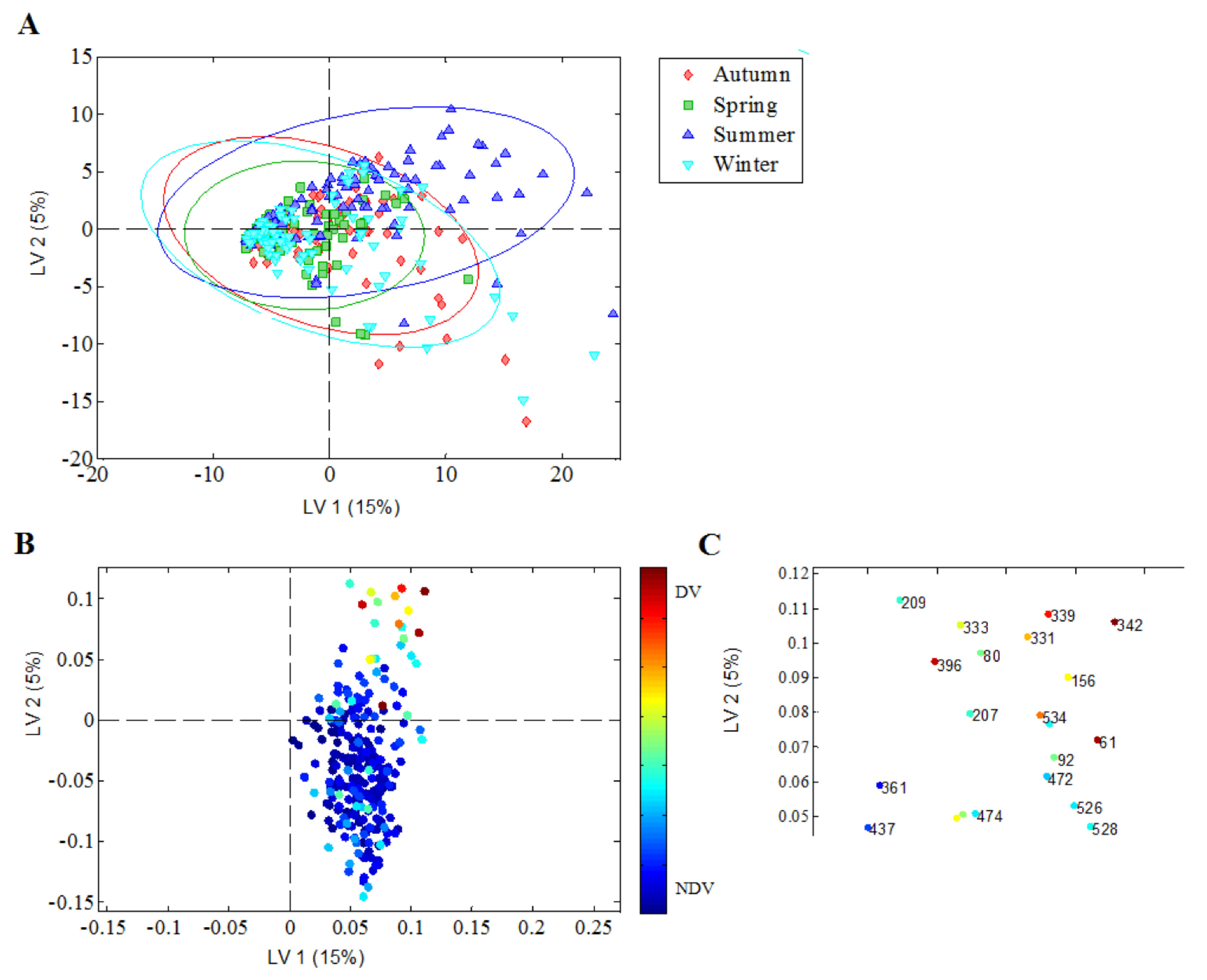
3.5. Discriminatory Nasopharyngeal Bacterial Richness in Relation to Season of Birth
4. Discussion
4.1. Main Findings
4.2. Interpretation
4.3. Strengths and Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Governance
Abbreviations
| COPSAC | Copenhagen Prospective Studies on Asthma in Childhood |
| T-RFLP | terminal restriction fragment length polymorphism |
| TRFs | terminal restriction fragments |
| T-REX | T-RFLP analysis EXpedited |
| PLSDA | Partial Least Square Discriminant Analysis |
| LV | Latent Variable |
References
- Bisgaard, H.; Hermansen, M.N.; Buchvald, F.; Loland, L.; Halkjaer, L.B.; Bønnelykke, K.; Brasholt, M.; Heltberg, A.; Vissing, N.H.; Thorsen, S.V.; et al. Childhood asthma after bacterial colonization of the airway in neonates. N. Engl. J. Med. 2007, 357, 1487–1495. [Google Scholar] [CrossRef]
- Von Linstow, M.-L.; Schønning, K.; Hoegh, A.M.; Sevelsted, A.; Vissing, N.H.; Bisgaard, H. Neonatal Airway Colonization Is Associated with Troublesome Lung Symptoms in Infants. Am. J. Respir. Crit. Care Med. 2013, 188, 1041–1042. [Google Scholar] [CrossRef] [PubMed]
- Vissing, N.H.; Chawes, B.L.K.; Bisgaard, H. Increased risk of pneumonia and bronchiolitis after bacterial colonization of the airways as neonates. Am. J. Respir. Crit. Care Med. 2013, 188, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Følsgaard, N.V.; Schjørring, S.; Chawes, B.L.; Rasmussen, M.A.; Krogfelt, K.A.; Brix, S.; Bisgaard, H. Pathogenic Bacteria Colonizing the Airways in Asymptomatic Neonates Stimulates Topical Inflammatory Mediator Release. Am. J. Respir. Crit. Care Med. 2013, 187, 589–595. [Google Scholar] [CrossRef]
- Thorsen, J.; Rasmussen, M.A.; Waage, J.; Mortensen, M.; Brejnrod, A.; Bønnelykke, K.; Chawes, B.L.; Brix, S.; Sørensen, S.J.; Stokholm, J.; et al. Infant airway microbiota and topical immune perturbations in the origins of childhood asthma. Nat Commun 2019, 10, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, M.S.; Brejnrod, A.D.; Roggenbuck, M.; Abu Al-Soud, W.; Balle, C.; Krogfelt, K.A.; Stokholm, J.; Thorsen, J.; Waage, J.; Rasmussen, M.A.; et al. The developing hypopharyngeal microbiota in early life. Microbiome 2016, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- Horz, H.-P.; Ten Haaf, A.; Kessler, O.; Said Yekta, S.; Seyfarth, I.; Hettlich, M.; Lampert, F.; Küpper, T.; Conrads, G. T-RFLP-based differences in oral microbial communities as risk factor for development of oral diseases under stress. Environ. Microbiol. Rep. 2012, 4, 390–397. [Google Scholar] [CrossRef]
- Brugger, S.D.; Frei, L.; Frey, P.M.; Aebi, S.; Mühlemann, K.; Hilty, M. 16S rRNA terminal restriction fragment length polymorphism for the characterization of the nasopharyngeal microbiota. PLoS ONE 2012, 7, e52241. [Google Scholar] [CrossRef] [Green Version]
- Bisgaard, H.; Li, N.; Bonnelykke, K.; Chawes, B.L.K.; Skov, T.; Paludan-Müller, G.; Stokholm, J.; Smith, B.; Krogfelt, K.A. Reduced diversity of the intestinal microbiota during infancy is associated with increased risk of allergic disease at school age. J. Allergy Clin. Immunol. 2011, 128, 646–652.e1–5. [Google Scholar] [CrossRef]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Boland, M.R.; Shahn, Z.; Madigan, D.; Hripcsak, G.; Tatonetti, N.P. Birth month affects lifetime disease risk: A phenome-wide method. J. Am. Med. Inf. Assoc. 2015, 22, 1042–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.-C.; Yang, K.D.; Wu, M.-T.M.; Wen, Y.-F.; Hsi, E.; Chang, J.-C.; Lin, Y.-M.; Kuo, H.-C.; Chang, W.-P. Close Correlation between Season of Birth and the Prevalence of Bronchial Asthma in a Taiwanese Population. PLoS ONE 2013, 8, e80285. [Google Scholar] [CrossRef] [PubMed]
- Vassallo, M.F.; Banerji, A.; Rudders, S.A.; Clark, S.; Mullins, R.J.; Camargo, C.A. Season of Birth is Associated with Food Allergy in Children. Ann. Allergy Asthma Immunol. 2010, 104, 307–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisgaard, H. The Copenhagen Prospective Study on Asthma in Childhood (COPSAC): Design, rationale, and baseline data from a longitudinal birth cohort study. Ann. Allergy Asthma Immunol. 2004, 93, 381–389. [Google Scholar] [CrossRef]
- Jensen, J.S.; Hansen, H.T.; Lind, K. Isolation of Mycoplasma genitalium strains from the male urethra. J. Clin. Microbiol. 1996, 34, 286–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tully, J.G.; Whitcomb, R.F.; Clark, H.F.; Williamson, D.L. Pathogenic mycoplasmas: Cultivation and vertebrate pathogenicity of a new spiroplasma. Science (New York N. Y.) 1977, 195, 892–894. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Schütte, U.M.E.; Abdo, Z.; Bent, S.J.; Shyu, C.; Williams, C.J.; Pierson, J.D.; Forney, L.J. Advances in the use of terminal restriction fragment length polymorphism (T-RFLP) analysis of 16S rRNA genes to characterize microbial communities. Appl. Microbiol. Biotechnol. 2008, 80, 365–380. [Google Scholar] [CrossRef]
- Fortuna, A.-M.; Marsh, T.L.; Honeycutt, C.W.; Halteman, W.A. Use of primer selection and restriction enzymes to assess bacterial community diversity in an agricultural soil used for potato production via terminal restriction fragment length polymorphism. Appl. Microbiol. Biotechnol. 2011, 91, 1193–1202. [Google Scholar] [CrossRef]
- Culman, S.W.; Bukowski, R.; Gauch, H.G.; Cadillo-Quiroz, H.; Buckley, D.H. T-REX: Software for the processing and analysis of T-RFLP data. Bmc Bioinform. 2009, 10, 171. [Google Scholar] [CrossRef] [Green Version]
- Shyu, C.; Soule, T.; Bent, S.J.; Foster, J.A.; Forney, L.J. MiCA: A web-based tool for the analysis of microbial communities based on terminal-restriction fragment length polymorphisms of 16S and 18S rRNA genes. Microb. Ecol. 2007, 53, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, C.W.; Kitts, C.L. Variation between observed and true Terminal Restriction Fragment length is dependent on true TRF length and purine content. J. Microbiol. Methods 2003, 54, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Wold, S.; Sjostrom, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Bowers, R.M.; Sullivan, A.P.; Costello, E.K.; Collett, J.L.; Knight, R.; Fierer, N. Sources of bacteria in outdoor air across cities in the midwestern United States. Appl. Environ. Microbiol. 2011, 77, 6350–6356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolini, V.; Gandolfi, I.; Ambrosini, R.; Bestetti, G.; Innocente, E.; Rampazzo, G.; Franzetti, A. Temporal variability and effect of environmental variables on airborne bacterial communities in an urban area of Northern Italy. Appl. Microbiol. Biotechnol. 2013, 97, 6561–6570. [Google Scholar] [CrossRef] [Green Version]
- Bogaert, D.; Keijser, B.; Huse, S.; Rossen, J.; Veenhoven, R.; van Gils, E.; Bruin, J.; Montijn, R.; Bonten, M.; Sanders, E. Variability and diversity of nasopharyngeal microbiota in children: A metagenomic analysis. PLoS ONE 2011, 6, e17035. [Google Scholar] [CrossRef] [Green Version]
- Toivonen, L.; Hasegawa, K.; Ajami, N.J.; Celedón, J.C.; Mansbach, J.M.; Petrosino, J.F.; Camargo, C.A. Circulating 25-hydroxyvitamin D, nasopharyngeal microbiota, and bronchiolitis severity. Pediatr. Allergy Immunol. 2018, 29, 877–880. [Google Scholar] [CrossRef]
- Wjst, M.; Dold, S.; Reitmeir, P.; Stiepel, E.; Von Mutius, E. Month of birth and allergic disease at the age of 10. Clin. Exp. Allergy 1992, 22, 1026–1031. [Google Scholar] [CrossRef]
- Wu, P.; Dupont, W.D.; Griffin, M.R.; Carroll, K.N.; Mitchel, E.F.; Gebretsadik, T.; Hartert, T.V. Evidence of a causal role of winter virus infection during infancy in early childhood asthma. Am. J. Respir. Crit. Care Med. 2008, 178, 1123–1129. [Google Scholar] [CrossRef] [Green Version]
- Hilty, M.; Burke, C.; Pedro, H.; Cardenas, P.; Bush, A.; Bossley, C.; Davies, J.; Ervine, A.; Poulter, L.; Pachter, L.; et al. Disordered microbial communities in asthmatic airways. PLoS ONE 2010, 5, e8578. [Google Scholar] [CrossRef] [Green Version]
- Hilty, M.; Qi, W.; Brugger, S.D.; Frei, L.; Agyeman, P.; Frey, P.M.; Aebi, S.; Mühlemann, K. Nasopharyngeal microbiota in infants with acute otitis media. J. Infect. Dis. 2012, 205, 1048–1055. [Google Scholar] [CrossRef] [PubMed]
- Biasucci, G.; Benenati, B.; Morelli, L.; Bessi, E.; Boehm, G. Cesarean delivery may affect the early biodiversity of intestinal bacteria. J. Nutr. 2008, 138, 1796S–1800S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsson, H.E.; Abrahamsson, T.R.; Jenmalm, M.C.; Harris, K.; Quince, C.; Jernberg, C.; Björkstén, B.; Engstrand, L.; Andersson, A.F. Decreased gut microbiota diversity, delayed Bacteroidetes colonisation and reduced Th1 responses in infants delivered by Caesarean section. Gut 2014, 63, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stokholm, J.; Thorsen, J.; Chawes, B.L.; Schjørring, S.; Krogfelt, K.A.; Bønnelykke, K.; Bisgaard, H. Cesarean section changes neonatal gut colonization. J. Allergy Clin. Immunol. 2016, 138, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Ege, M.J.; Mayer, M.; Normand, A.-C.; Genuneit, J.; Cookson, W.O.; Braun-fahrländer, C.; Heederik, D.; Piarroux, R.; von Mutius, E. GABRIELA Transregio 22 Study Group Exposure to environmental microorganisms and childhood asthma. N. Engl. J. Med. 2011, 364, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Sakwinska, O.; Bastic Schmid, V.; Berger, B.; Bruttin, A.; Keitel, K.; Lepage, M.; Moine, D.; Ngom Bru, C.; Brüssow, H.; Gervaix, A. Nasopharyngeal microbiota in healthy children and pneumonia patients. J. Clin. Microbiol. 2014, 52, 1590–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettigrew, M.M.; Laufer, A.S.; Gent, J.F.; Kong, Y.; Fennie, K.P.; Metlay, J.P. Upper respiratory tract microbial communities, acute otitis media pathogens, and antibiotic use in healthy and sick children. Appl. Environ. Microbiol. 2012, 78, 6262–6270. [Google Scholar] [CrossRef] [Green Version]
- Cardenas, P.A.; Cooper, P.J.; Cox, M.J.; Chico, M.; Arias, C.; Moffatt, M.F.; Cookson, W.O. Upper airways microbiota in antibiotic-naïve wheezing and healthy infants from the tropics of rural Ecuador. PLoS ONE 2012, 7, e46803. [Google Scholar] [CrossRef]
- Azad, M.B.; Konya, T.; Maughan, H.; Guttman, D.S.; Field, C.J.; Sears, M.R.; Becker, A.B.; Scott, J.A.; Kozyrskyj, A.L. Infant gut microbiota and the hygiene hypothesis of allergic disease: Impact of household pets and siblings on microbiota composition and diversity. Allergyasthma Clin. Immunol. 2013, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Stokholm, J.; Schjørring, S.; Pedersen, L.; Bischoff, A.L.; Følsgaard, N.; Carson, C.G.; Chawes, B.; Bønnelykke, K.; Mølgaard, A.; Krogfelt, K.A.; et al. Living with cat and dog increases vaginal colonization with E. coli in pregnant women. PLoS ONE 2012, 7, e46226. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Widmer, F. Reliability for detecting composition and changes of microbial communities by T-RFLP genetic profiling. Fems Microbiol. Ecol. 2008, 63, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.T.; Marsh, T.L.; Cheng, H.; Forney, L.J. Characterization of microbial diversity by determining terminal restriction fragment length polymorphisms of genes encoding 16S rRNA. Appl. Environ. Microbiol. 1997, 63, 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, M.; Enkerli, J.; Widmer, F. Residual polymerase activity-induced bias in terminal restriction fragment length polymorphism analysis. Environ. Microbiol. 2007, 9, 555–559. [Google Scholar] [CrossRef]
- Mika, M.; Mack, I.; Korten, I.; Qi, W.; Aebi, S.; Frey, U.; Latzin, P.; Hilty, M. Dynamics of the nasal microbiota in infancy: A prospective cohort study. J. Allergy Clin. Immunol. 2015, 135, 905–912.e11. [Google Scholar] [CrossRef] [Green Version]
- Bisgaard, H.; Vissing, N.H.; Carson, C.G.; Bischof, A.L.; Følsgaard, N.V.; Kreiner-Møller, E.; Chawes, B.; Stokholm, J.; Pedersen, L.; Bjarnadóttir, E.; et al. Deep Phenotyping of the Unselected COPSAC2010 Birth Cohort Study. Clin. Exp. Allergy 2013, 43, 1384–1394. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Variables | No. | No. of Cases (%) | Richness in TRFs Median (IQR) | p-Value 1 | |
|---|---|---|---|---|---|
| Atopic predisposition | |||||
| Maternal asthma 2 | 328 | 328 (100) | - | - | |
| Maternal rhinitis - Yes - No | 327 | 246 (75.2) | 36 (15–55) 34 (14–52) | 0.67 | |
| Maternal dermatitis - Yes - No | 328 | 163 (49.7) | 36 (16–54) 34 (13–56) | 0.68 | |
| Paternal asthma - Yes - No | 318 | 51 (16.0) | 42 (17–61) 32 (14–55) | 0.11 | |
| Paternal rhinitis - Yes - No | 316 | 107 (33.9) | 37 (19–53) 32 (12–60) | 0.18 | |
| Paternal dermatitis - Yes - No | 316 | 43 (13.6) | 45 (22–67) 35 (14–55) | 0.044 3 | |
| Social status | |||||
| Family income, 1000 EUR/year - <53.6 - 53.6–80.4 - >80.4 | 307 | 81 (27.4) 149 (48.5) 77 (25.1) | 25 (9–49) 40 (18–55) 40 (15–61) | 0.031 4 | |
| Mother’s education - College - Elementary/Medium - University | 301 | 179 (59.5) 82 (27.2) 40 (13.3) | 35 (16–55) 37 (18–61) 37 (12–55) | 0.76 | |
| Mother’s occupation - Unemployed - Non-professional - Professional - Student | 306 | 29 (9.5) 103 (33.7) 143 (46.7) 31 (10.1) | 26 (6–45) 31 (16–52) 38 (18–61) 41 (14–58) | 0.14 | |
| Pregnancy | |||||
| Previous deliveries - None - 1 - 2 or more | 300 | 178 (59.3) 91 (30.3) 31 (10.3) | 36 (16–55) 41 (15–61) 40 (16–54) | 0.91 | |
| Antibiotic use in 3rd trimester - Yes - No | 328 | 44 (13.4) | 26 (12–51) 36 (15–55) | 0.28 | |
| Paracetamol use in 3rd trimester - Yes - No | 328 | 39 (11.9) | 40 (14–67) 33 (14–54) | 0.18 | |
| Smoking in 3rd trimester - Yes - No | 328 | 48 (14.6) | 27 (17–50) 35 (14–55) | 0.59 | |
| Alcohol in 3rd trimester - Yes - No | 328 | 51 (15.5) | 25 (14–61) 35 (15–54) | 0.86 | |
| Birth | |||||
| Gender - Female - Male | 328 | 159 (48.5) 169 (51.5) | 29 (15–53) 38 (14–60) | 0.24 | |
| Mother’s age at birth, years - <25 - 25–35 - ≥35 | 328 | 38 (11.6) 238 (72.6) 52 (15.9) | 27 (8–47) 36 (15–58) 36 (15–53) | 0.14 | |
| Gestational age, weeks - <≤38 - >38 ≤41 - >41 | 328 | 66 (20.1) 130 (39.6) 132 (40.2) | 29 (13–55) 36 (15–54) 36 (14–57) | 0.93 | |
| Mode of delivery - Natural birth - Caesarean section | 328 | 259 (79.0) 69 (21.0) | 32 (15–55) 40 (14–55) | 0.77 | |
| Apgar score ≥9 at 5min - Yes - No | 325 | 315 (96.9) | 35 (14–55) 35 (13–51) | 0.95 | |
| Season of birth - Winter - Spring - Summer - Autumn | 328 | 77 (23.5) 69 (21.0) 87 (26.5) 95 (29.0) | 25 (14–55) 26 (7–51) 46 (28–69) 31 (12–51) | 0.003 5 | |
| Postnatal exposures | |||||
| Cat at birth - Yes - No | 303 | 38 (12.5) | 23 (10–61) 37 (15–55) | 0.32 | |
| Dog at birth - Yes - No | 311 | 34 (10.9) | 34 (20–62) 37 (15–55) | 0.46 | |
| Older siblings at birth - None - 1 - 2 or more | 309 | 185 (59.9) 89 (28.8) 35 (11.3) | 32 (15–52) 40 (15–61) 40 (17–55) | 0.54 | |
| Solely breastfed > 30 days - Yes - No | 321 | 195 (60.7) | 37 (15–55) 30 (14–56) | 0.67 | |
| Specimen collection | |||||
| Child age, days - ≤30 - 31–60 -- ≥61 | 321 | 35 (10.9) 230 (71.7) 56 (17.4) | 17 (9–46) 37 (16–59) 30 (15–53) | 0.73 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schoos, A.-M.M.; Kragh, M.; Ahrens, P.; Kuhn, K.G.; Rasmussen, M.A.; Chawes, B.L.; Jensen, J.S.; Brix, S.; Bisgaard, H.; Stokholm, J. Season of Birth Impacts the Neonatal Nasopharyngeal Microbiota. Children 2020, 7, 45. https://doi.org/10.3390/children7050045
Schoos A-MM, Kragh M, Ahrens P, Kuhn KG, Rasmussen MA, Chawes BL, Jensen JS, Brix S, Bisgaard H, Stokholm J. Season of Birth Impacts the Neonatal Nasopharyngeal Microbiota. Children. 2020; 7(5):45. https://doi.org/10.3390/children7050045
Chicago/Turabian StyleSchoos, Ann-Marie Malby, Marie Kragh, Peter Ahrens, Katrin Gaardbo Kuhn, Morten Arendt Rasmussen, Bo Lund Chawes, Jørgen Skov Jensen, Susanne Brix, Hans Bisgaard, and Jakob Stokholm. 2020. "Season of Birth Impacts the Neonatal Nasopharyngeal Microbiota" Children 7, no. 5: 45. https://doi.org/10.3390/children7050045
APA StyleSchoos, A.-M. M., Kragh, M., Ahrens, P., Kuhn, K. G., Rasmussen, M. A., Chawes, B. L., Jensen, J. S., Brix, S., Bisgaard, H., & Stokholm, J. (2020). Season of Birth Impacts the Neonatal Nasopharyngeal Microbiota. Children, 7(5), 45. https://doi.org/10.3390/children7050045