
Prenatal Maternal Lipopolysaccharide and Mild Newborn Hyperoxia Increase Intrapulmonary Airway but Not Vessel Reactivity in a Mouse Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Mice and LPS Injections
2.3. Hyperoxia Exposure
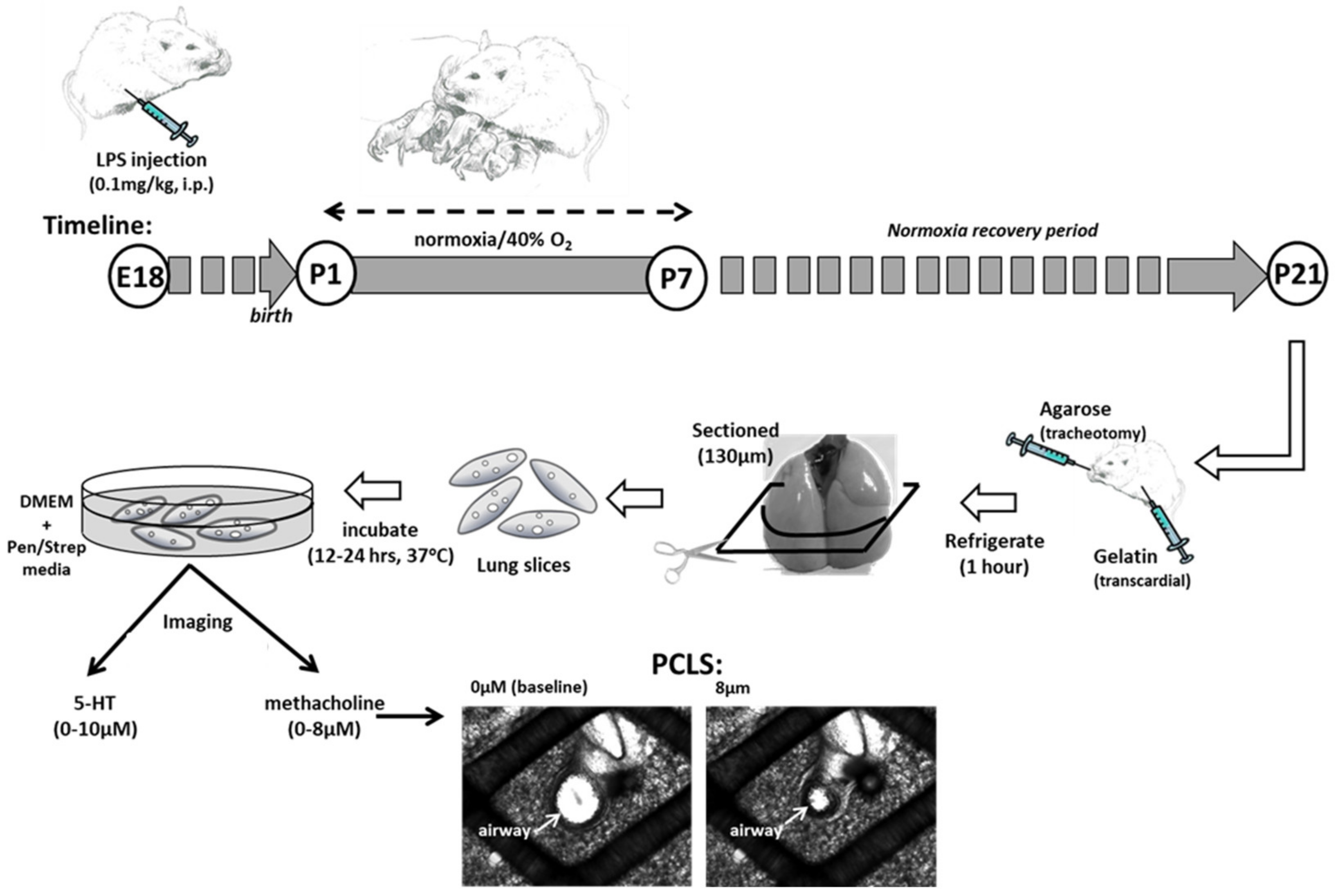
2.4. Lung Slice Preparation and Reactivity Measurements
2.5. qRT-PCR
2.6. Materials
2.7. Incusion and Exclusion Criteria
2.8. Statistical Analysis
3. Results
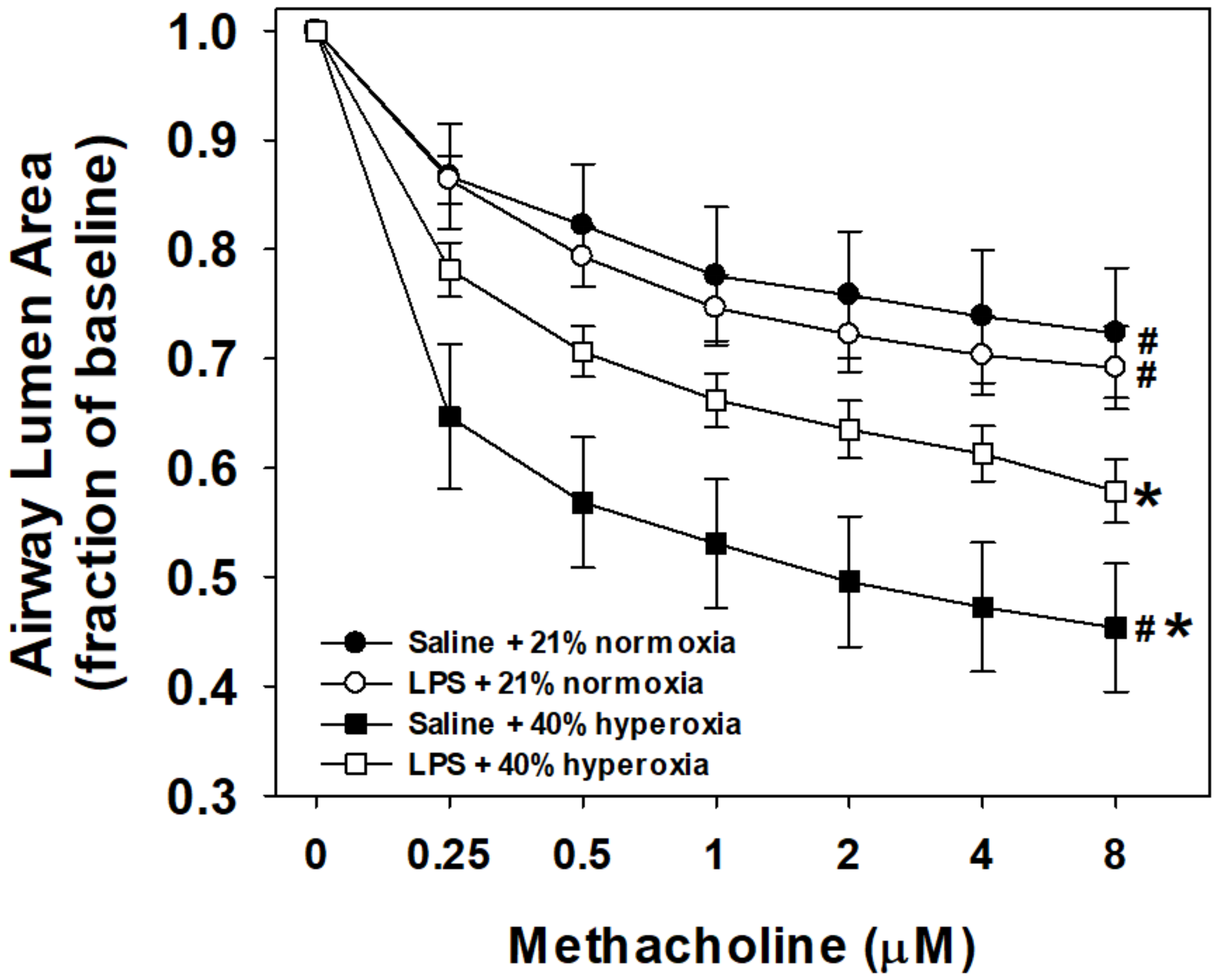
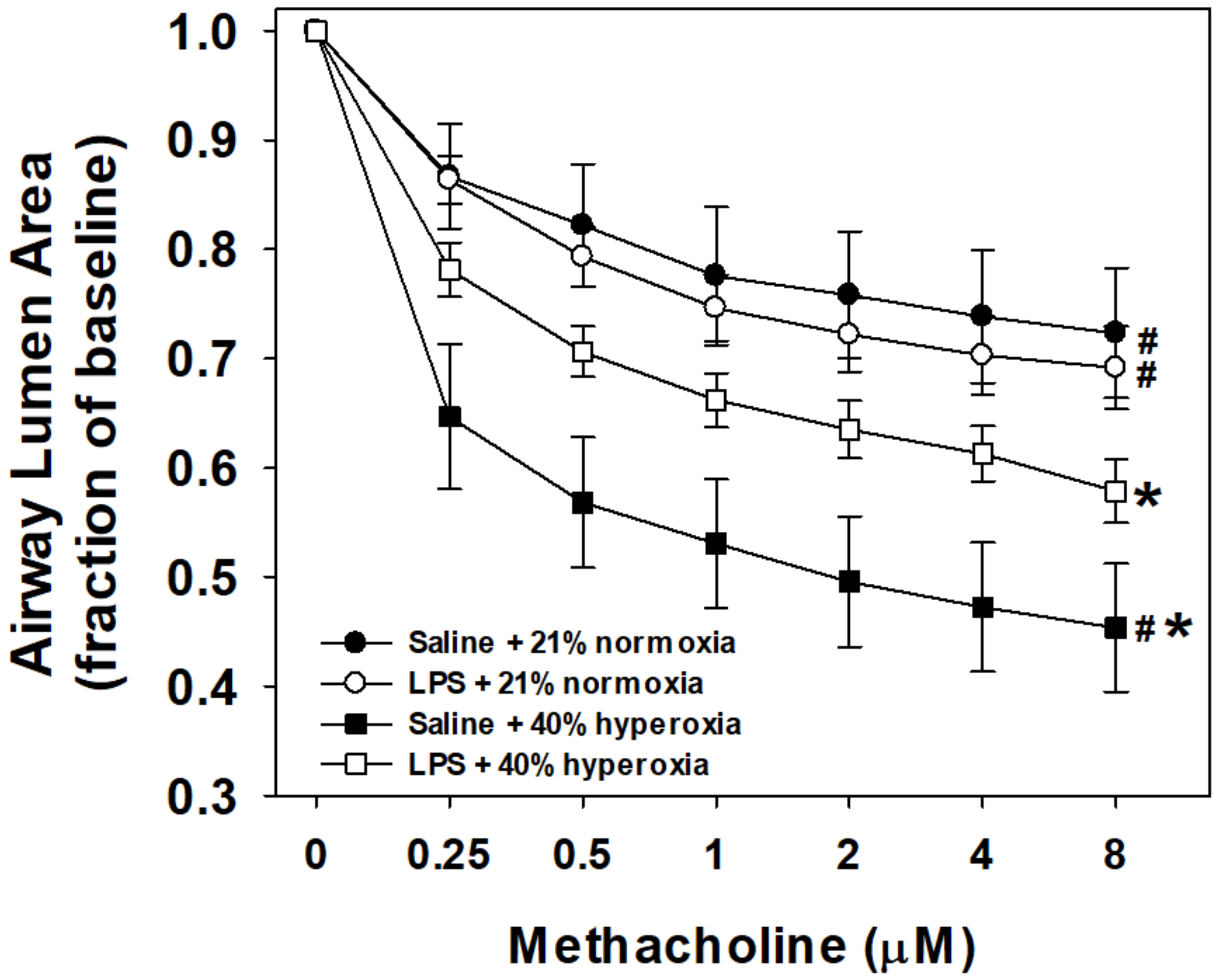
3.1. Airway Reactivity to Methacholine
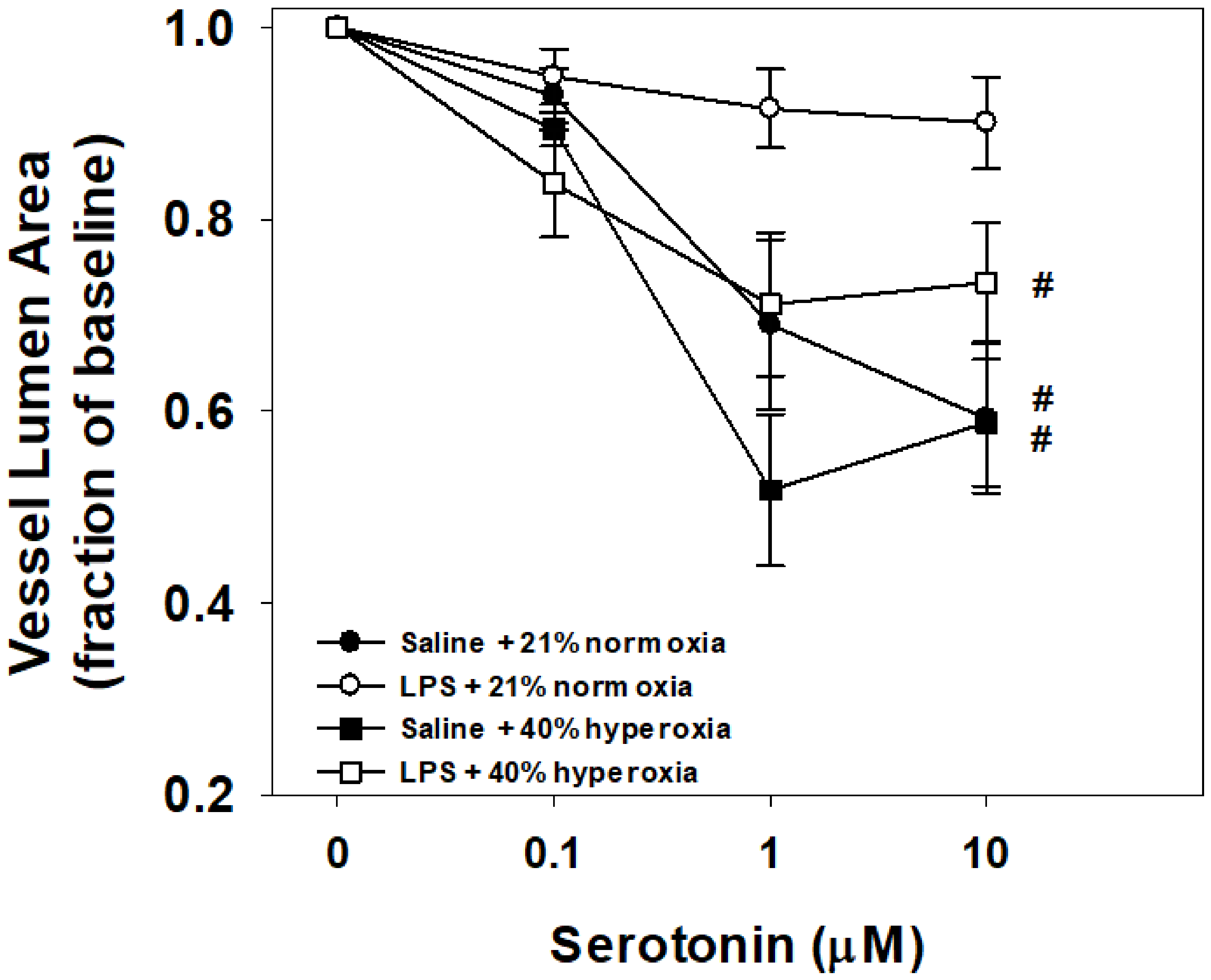
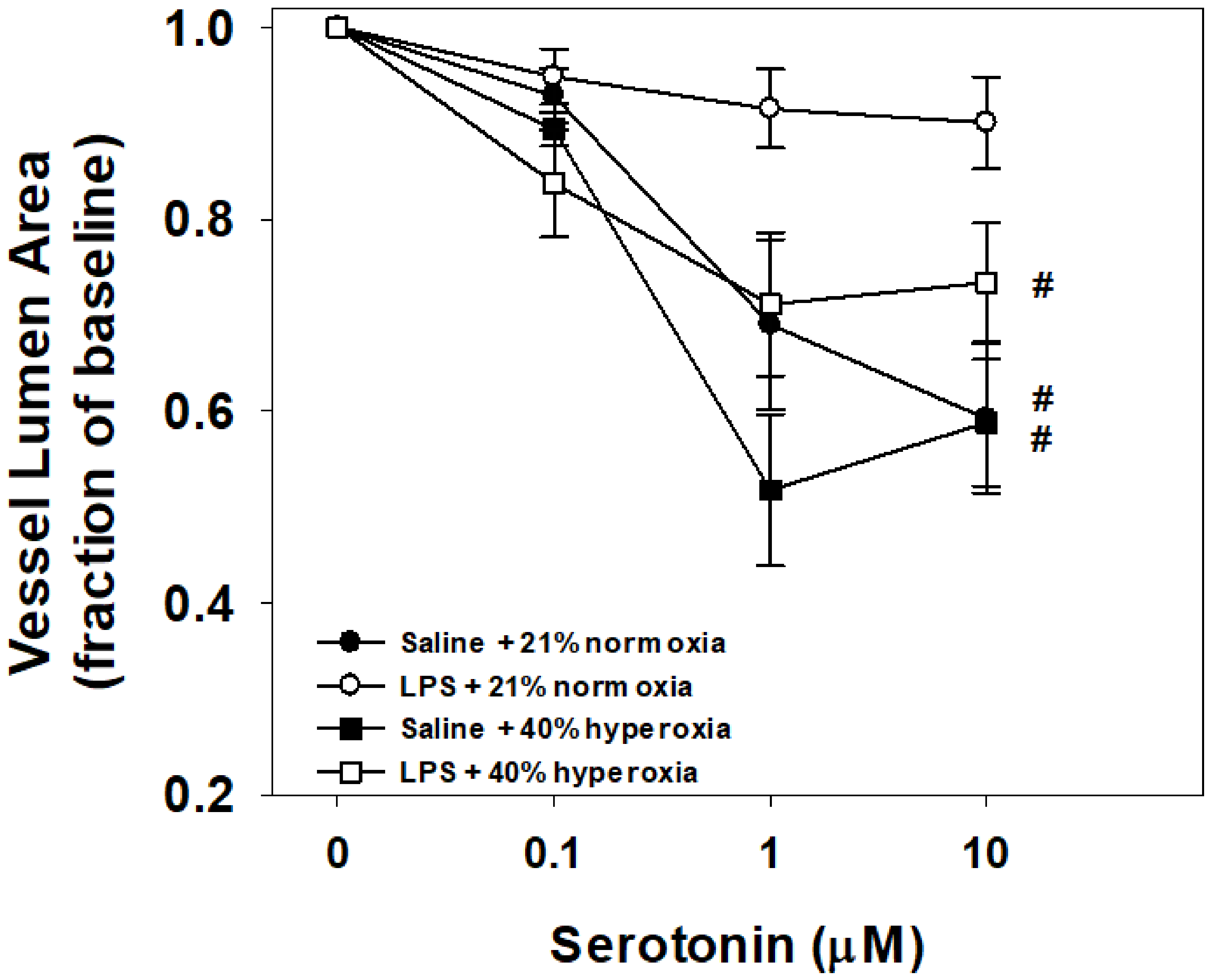
3.2. Vessel Reactivity to Serotonin
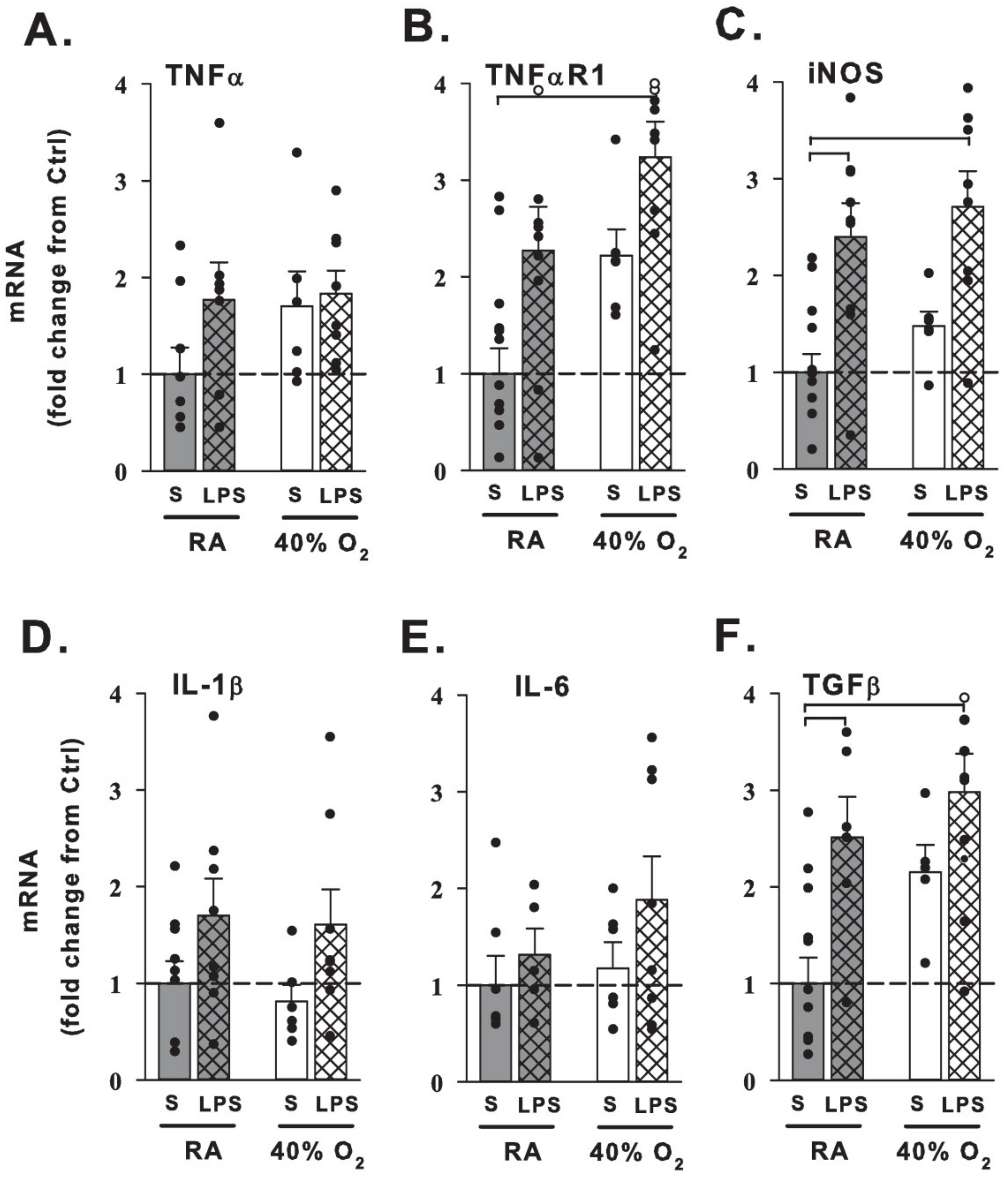
3.3. Lung mRNA Expression
4. Discussion
4.1. Maternal LPS and Hyperoxia Effects on Airway Reactivity to Methacholine
4.2. Maternal LPS and Hyperoxia Effects on Vessel Reactivity to Serotonin
4.3. Maternal LPS and Hyperoxia Effects on Lung Gene Expression
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stoll, B.J.; Hansen, N.I.; Bell, E.F.; Walsh, M.C.; Carlo, W.A.; Shankaran, S.; Laptook, A.R.; Sanchez, P.J.; Van Meurs, K.P.; Wyckoff, M.; et al. Trends in Care Practices, Morbidity, and Mortality of Extremely Preterm Neonates, 1993–2012. JAMA 2015, 314, 1039–1051. [Google Scholar] [CrossRef] [Green Version]
- Jaakkola, J.J.; Ahmed, P.; Ieromnimon, A.; Goepfert, P.; Laiou, E.; Quansah, R.; Jaakkola, M.S. Preterm delivery and asthma: A systematic review and meta-analysis. J. Allergy Clin. Immunol. 2006, 118, 823–830. [Google Scholar] [CrossRef]
- Joshi, S.; Powell, T.; Watkins, W.J.; Drayton, M.; Williams, E.M.; Kotecha, S. Exercise-induced bronchoconstriction in school-aged children who had chronic lung disease in infancy. J. Pediatrics 2013, 162, 813–818.e811. [Google Scholar] [CrossRef]
- Been, J.V.; Lugtenberg, M.J.; Smets, E.; van Schayck, C.P.; Kramer, B.W.; Mommers, M.; Sheikh, A. Preterm birth and childhood wheezing disorders: A systematic review and meta-analysis. PLoS Med. 2014, 11, e1001596. [Google Scholar] [CrossRef] [PubMed]
- Reyburn, B.; Martin, R.J.; Prakash, Y.S.; MacFarlane, P.M. Mechanisms of injury to the preterm lung and airway: Implications for long-term pulmonary outcome. Neonatology 2012, 101, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Di Fiore, J.M.; Dylag, A.M.; Honomichl, R.D.; Hibbs, A.M.; Martin, R.J.; Tatsuoka, C.; Raffay, T.M. Early inspired oxygen and intermittent hypoxemic events in extremely premature infants are associated with asthma medication use at 2 years of age. J. Perinatol. 2019, 39, 203–211. [Google Scholar] [CrossRef]
- Dylag, A.M.; Kopin, H.G.; O’Reilly, M.A.; Wang, H.; Davis, S.D.; Ren, C.L.; Pryhuber, G.S. Early Neonatal Oxygen Exposure Predicts Pulmonary Morbidity and Functional Deficits at 1 Year. J. Pediatrics 2020, 223, 20–28.e22. [Google Scholar] [CrossRef] [PubMed]
- Stevens, T.P.; Dylag, A.; Panthagani, I.; Pryhuber, G.; Halterman, J. Effect of cumulative oxygen exposure on respiratory symptoms during infancy among VLBW infants without bronchopulmonary dysplasia. Pediatric Pulmonol. 2010, 45, 371–379. [Google Scholar] [CrossRef]
- Onugha, H.; MacFarlane, P.M.; Mayer, C.A.; Abrah, A.; Jafri, A.; Martin, R.J. Airway Hyperreactivity Is Delayed after Mild Neonatal Hyperoxic Exposure. Neonatology 2015, 108, 65–72. [Google Scholar] [CrossRef]
- Wang, H.; Jafri, A.; Martin, R.J.; Nnanabu, J.; Farver, C.; Prakash, Y.S.; MacFarlane, P.M. Severity of neonatal hyperoxia determines structural and functional changes in developing mouse airway. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 307, L295–L301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dylag, A.M.; Haak, J.; Yee, M.; O’Reilly, M.A. Pulmonary mechanics and structural lung development after neonatal hyperoxia in mice. Pediatric Res. 2020, 87, 1201–1210. [Google Scholar] [CrossRef]
- Kumar, V.H.S. Diagnostic Approach to Pulmonary Hypertension in Premature Neonates. Children 2017, 4, 75. [Google Scholar] [CrossRef] [Green Version]
- Bhat, R.; Salas, A.A.; Foster, C.; Carlo, W.A.; Ambalavanan, N. Prospective analysis of pulmonary hypertension in extremely low birth weight infants. Pediatrics 2012, 129, e682–e689. [Google Scholar] [CrossRef] [Green Version]
- Dumas de la Roque, E.; Smeralda, G.; Quignard, J.F.; Freund-Michel, V.; Courtois, A.; Marthan, R.; Muller, B.; Guibert, C.; Dubois, M. Altered vasoreactivity in neonatal rats with pulmonary hypertension associated with bronchopulmonary dysplasia: Implication of both eNOS phosphorylation and calcium signaling. PLoS ONE 2017, 12, e0173044. [Google Scholar] [CrossRef] [Green Version]
- Bui, C.B.; Kolodziej, M.; Lamanna, E.; Elgass, K.; Sehgal, A.; Rudloff, I.; Schwenke, D.O.; Tsuchimochi, H.; Kroon, M.; Cho, S.X.; et al. Interleukin-1 Receptor Antagonist Protects Newborn Mice Against Pulmonary Hypertension. Front. Immunol. 2019, 10, 1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belik, J.; Jankov, R.P.; Pan, J.; Tanswell, A.K. Chronic O2 exposure enhances vascular and airway smooth muscle contraction in the newborn but not adult rat. J. Appl. Physiol. 2003, 94, 2303–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belik, J.; Jankov, R.P.; Pan, J.; Yi, M.; Chaudhry, I.; Tanswell, A.K. Chronic O2 exposure in the newborn rat results in decreased pulmonary arterial nitric oxide release and altered smooth muscle response to isoprostane. J. Appl. Physiol. 2004, 96, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Bastek, J.A.; Gomez, L.M.; Elovitz, M.A. The role of inflammation and infection in preterm birth. Clin. Perinatol. 2011, 38, 385–406. [Google Scholar] [CrossRef] [PubMed]
- Getahun, D.; Strickland, D.; Zeiger, R.S.; Fassett, M.J.; Chen, W.; Rhoads, G.G.; Jacobsen, S.J. Effect of chorioamnionitis on early childhood asthma. Arch. Pediatrics Adolesc. Med. 2010, 164, 187–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Yu, Y.; Story, R.E.; Pongracic, J.A.; Gupta, R.; Pearson, C.; Ortiz, K.; Bauchner, H.C.; Wang, X. Prematurity, chorioamnionitis, and the development of recurrent wheezing: A prospective birth cohort study. J. Allergy Clin. Immunol. 2008, 121, 878–884.e876. [Google Scholar] [CrossRef]
- McDowell, K.M.; Jobe, A.H.; Fenchel, M.; Hardie, W.D.; Gisslen, T.; Young, L.R.; Chougnet, C.A.; Davis, S.D.; Kallapur, S.G. Pulmonary Morbidity in Infancy after Exposure to Chorioamnionitis in Late Preterm Infants. Ann. Am. Thorac. Soc. 2016, 13, 867–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fricke, E.M.; Elgin, T.G.; Gong, H.; Reese, J.; Gibson-Corley, K.N.; Weiss, R.M.; Zimmerman, K.; Bowdler, N.C.; Kalantera, K.M.; Mills, D.A.; et al. Lipopolysaccharide-induced maternal inflammation induces direct placental injury without alteration in placental blood flow and induces a secondary fetal intestinal injury that persists into adulthood. Am. J. Reprod. Immunol. 2018, 79, e12816. [Google Scholar] [CrossRef] [Green Version]
- Gayle, D.A.; Beloosesky, R.; Desai, M.; Amidi, F.; Nunez, S.E.; Ross, M.G. Maternal LPS induces cytokines in the amniotic fluid and corticotropin releasing hormone in the fetal rat brain. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R1024–R1029. [Google Scholar] [CrossRef] [PubMed]
- Velten, M.; Gorr, M.W.; Youtz, D.J.; Velten, C.; Rogers, L.K.; Wold, L.E. Adverse perinatal environment contributes to altered cardiac development and function. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H1334–H1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faksh, A.; Britt, R.D., Jr.; Vogel, E.R.; Kuipers, I.; Thompson, M.A.; Sieck, G.C.; Pabelick, C.M.; Martin, R.J.; Prakash, Y.S. Effects of antenatal lipopolysaccharide and postnatal hyperoxia on airway reactivity and remodeling in a neonatal mouse model. Pediatric Res. 2016, 79, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Royce, S.G.; Nold, M.F.; Bui, C.; Donovan, C.; Lam, M.; Lamanna, E.; Rudloff, I.; Bourke, J.E.; Nold-Petry, C.A. Airway Remodeling and Hyperreactivity in a Model of Bronchopulmonary Dysplasia and Their Modulation by IL-1Ra. Am. J. Respir. Cell Mol. Biol. 2016, 55, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Velten, M.; Heyob, K.M.; Rogers, L.K.; Welty, S.E. Deficits in lung alveolarization and function after systemic maternal inflammation and neonatal hyperoxia exposure. J. Appl. Physiol. 2010, 108, 1347–1356. [Google Scholar] [CrossRef]
- Amy, R.W.; Bowes, D.; Burri, P.H.; Haines, J.; Thurlbeck, W.M. Postnatal growth of the mouse lung. J. Anat. 1977, 124 Pt 1, 131–151. [Google Scholar]
- Berger, J.; Bhandari, V. Animal models of bronchopulmonary dysplasia. The term mouse models. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 307, L936–L947. [Google Scholar] [CrossRef] [Green Version]
- Dylag, A.M.; Raffay, T.M. Rodent models of respiratory control and respiratory system development-Clinical significance. Respir. Physiol. Neurobiol. 2019, 268, 103249. [Google Scholar] [CrossRef]
- Perez, J.F.; Sanderson, M.J. The contraction of smooth muscle cells of intrapulmonary arterioles is determined by the frequency of Ca2+ oscillations induced by 5-HT and KCl. J. Gen. Physiol. 2005, 125, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Sanderson, M.J. Exploring lung physiology in health and disease with lung slices. Pulm Pharm. Ther. 2011, 24, 452–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, C.A.; Martin, R.J.; MacFarlane, P.M. Increased airway reactivity in a neonatal mouse model of continuous positive airway pressure. Pediatric Res. 2015. [Google Scholar] [CrossRef] [Green Version]
- Velten, M.; Britt, R.D., Jr.; Heyob, K.M.; Tipple, T.E.; Rogers, L.K. Maternal dietary docosahexaenoic acid supplementation attenuates fetal growth restriction and enhances pulmonary function in a newborn mouse model of perinatal inflammation. J. Nutr. 2014, 144, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Moss, T.J.; Davey, M.G.; Harding, R.; Newnham, J.P. Effects of intra-amniotic endotoxin on lung structure and function two months after term birth in sheep. J. Soc. Gynecol. Investig. 2002, 9, 220–225. [Google Scholar] [CrossRef]
- Normann, E.; Lacaze-Masmonteil, T.; Eaton, F.; Schwendimann, L.; Gressens, P.; Thebaud, B. A novel mouse model of Ureaplasma-induced perinatal inflammation: Effects on lung and brain injury. Pediatric Res. 2009, 65, 430–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.R.; Seedorf, G.J.; Muehlethaler, V.; Walker, D.L.; Markham, N.E.; Balasubramaniam, V.; Abman, S.H. Moderate postnatal hyperoxia accelerates lung growth and attenuates pulmonary hypertension in infant rats after exposure to intra-amniotic endotoxin. Am. J. Physiol. Lung Cell Mol. Physiol. 2010, 299, L735–L748. [Google Scholar] [CrossRef]
- Choi, C.W.; Kim, B.I.; Hong, J.S.; Kim, E.K.; Kim, H.S.; Choi, J.H. Bronchopulmonary dysplasia in a rat model induced by intra-amniotic inflammation and postnatal hyperoxia: Morphometric aspects. Pediatric Res. 2009, 65, 323–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiktinsky, M.H.; Morin, F.C., 3rd. Increasing oxygen tension dilates fetal pulmonary circulation via endothelium-derived relaxing factor. Am. J. Physiol. 1993, 265 Pt 2, H376–H380. [Google Scholar] [CrossRef]
- Iluchev, D.; Kostianev, S.; Atanassov, A.; Lazarov, S. Blood gases, electrolytes and metabolic monitoring in children with acute failure of vital functions. Acta Anaesthesiol. Scand. Suppl. 1995, 107, 249–255. [Google Scholar] [CrossRef]
- Askie, L.M.; Darlow, B.A.; Finer, N.; Schmidt, B.; Stenson, B.; Tarnow-Mordi, W.; Davis, P.G.; Carlo, W.A.; Brocklehurst, P.; Davies, L.C.; et al. Association Between Oxygen Saturation Targeting and Death or Disability in Extremely Preterm Infants in the Neonatal Oxygenation Prospective Meta-analysis Collaboration. JAMA 2018, 319, 2190–2201. [Google Scholar] [CrossRef]
- Mourani, P.M.; Sontag, M.K.; Younoszai, A.; Miller, J.I.; Kinsella, J.P.; Baker, C.D.; Poindexter, B.B.; Ingram, D.A.; Abman, S.H. Early pulmonary vascular disease in preterm infants at risk for bronchopulmonary dysplasia. Am. J. Respir Crit. Care Med. 2015, 191, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Thebaud, B.; Abman, S.H. Bronchopulmonary dysplasia: Where have all the vessels gone? Roles of angiogenic growth factors in chronic lung disease. Am. J. Respir Crit. Care Med. 2007, 175, 978–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourani, P.M.; Ivy, D.D.; Gao, D.; Abman, S.H. Pulmonary vascular effects of inhaled nitric oxide and oxygen tension in bronchopulmonary dysplasia. Am. J. Respir Crit. Care Med. 2004, 170, 1006–1013. [Google Scholar] [CrossRef]
- Frank, B.S.; Schafer, M.; Grenolds, A.; Ivy, D.D.; Abman, S.H.; Darst, J.R. Acute Vasoreactivity Testing during Cardiac Catheterization of Neonates with Bronchopulmonary Dysplasia-Associated Pulmonary Hypertension. J. Pediatrics 2019, 208, 127–133. [Google Scholar] [CrossRef]
- Jobe, A.H. Mechanisms of Lung Injury and Bronchopulmonary Dysplasia. Am. J. Perinatol. 2016, 33, 1076–1078. [Google Scholar] [CrossRef]
- Iliodromiti, Z.; Zygouris, D.; Sifakis, S.; Pappa, K.I.; Tsikouras, P.; Salakos, N.; Daniilidis, A.; Siristatidis, C.; Vrachnis, N. Acute lung injury in preterm fetuses and neonates: Mechanisms and molecular pathways. J. Matern. Fetal Neonatal Med. Off. J. Eur. Assoc. Perinat. Med. Fed. Asia Ocean. Perinat. Soc. Int. Soc. Perinat. Obs. 2013, 26, 1696–1704. [Google Scholar] [CrossRef] [PubMed]
- Baymakova, M.; Plochev, K.; Dikov, I.; Popov, G.T.; Mihaylova-Garnizova, R.; Kovaleva, V.; Kundurdjiev, T. Fever of Unknown Origin in a Bulgarian Hospital: Evaluation of 54 Cases for a Four Year-Period. J. Clin. Anal. Med. 2016, 7, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Papagianis, P.C.; Pillow, J.J.; Moss, T.J. Bronchopulmonary dysplasia: Pathophysiology and potential anti-inflammatory therapies. Paediatric Respir Rev. 2019, 30, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Maturu, P.; Wei-Liang, Y.; Androutsopoulos, V.P.; Jiang, W.; Wang, L.; Tsatsakis, A.M.; Couroucli, X.I. Quercetin attenuates the hyperoxic lung injury in neonatal mice: Implications for Bronchopulmonary dysplasia (BPD). Food Chem. Toxicol. 2018, 114, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Nold, M.F.; Mangan, N.E.; Rudloff, I.; Cho, S.X.; Shariatian, N.; Samarasinghe, T.D.; Skuza, E.M.; Pedersen, J.; Veldman, A.; Berger, P.J.; et al. Interleukin-1 receptor antagonist prevents murine bronchopulmonary dysplasia induced by perinatal inflammation and hyperoxia. Proc. Natl. Acad Sci. USA 2013, 110, 14384–14389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosenko, I.R.; Jobe, A.H. Intraamniotic endotoxin increases lung antioxidant enzyme activity in preterm lambs. Pediatric Res. 2003, 53, 679–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuper-Sassé, M.E.; MacFarlane, P.M.; Mayer, C.A.; Martin, R.J.; Prakash, Y.S.; Pabelick, C.M.; Raffay, T.M. Prenatal Maternal Lipopolysaccharide and Mild Newborn Hyperoxia Increase Intrapulmonary Airway but Not Vessel Reactivity in a Mouse Model. Children 2021, 8, 195. https://doi.org/10.3390/children8030195
Kuper-Sassé ME, MacFarlane PM, Mayer CA, Martin RJ, Prakash YS, Pabelick CM, Raffay TM. Prenatal Maternal Lipopolysaccharide and Mild Newborn Hyperoxia Increase Intrapulmonary Airway but Not Vessel Reactivity in a Mouse Model. Children. 2021; 8(3):195. https://doi.org/10.3390/children8030195
Chicago/Turabian StyleKuper-Sassé, Margaret E., Peter M. MacFarlane, Catherine A. Mayer, Richard J. Martin, Y. S. Prakash, Christina M. Pabelick, and Thomas M. Raffay. 2021. "Prenatal Maternal Lipopolysaccharide and Mild Newborn Hyperoxia Increase Intrapulmonary Airway but Not Vessel Reactivity in a Mouse Model" Children 8, no. 3: 195. https://doi.org/10.3390/children8030195
APA StyleKuper-Sassé, M. E., MacFarlane, P. M., Mayer, C. A., Martin, R. J., Prakash, Y. S., Pabelick, C. M., & Raffay, T. M. (2021). Prenatal Maternal Lipopolysaccharide and Mild Newborn Hyperoxia Increase Intrapulmonary Airway but Not Vessel Reactivity in a Mouse Model. Children, 8(3), 195. https://doi.org/10.3390/children8030195