1. Introduction
The development and accessibility of next-generation sequencing (NGS) technologies has made the generation of large amounts of genomic and transcriptomic data relatively straightforward. On the other hand, fast and reliable screening methods to characterize enzymes identified from these data are generally lacking, resulting in an ever-increasing number of enzymes awaiting biochemical characterization. Several methods have been developed, but the use of hyphenated analytical chemistry technologies especially has been shown to have the potential for fast biochemical screenings. This includes techniques such as gas chromatography-mass spectroscopy (GC-MS) and liquid chromatography-mass spectroscopy (LC-MS) [
1].
Sesquiterpenoids are a subgroup of terpenoids that contain a core structure of 15 carbon atoms. Sesquiterpene hydrocarbons and sesquiterpenoids with a single hydroxyl group are often very volatile, and these are best analyzed using gas chromatography (GC) combined with mass spectroscopy (MS) [
2,
3]. The first step of sesquiterpenoid biosynthesis is performed by sesquiterpene synthases (sqTPS) [
4]. SqTPSs utilize farnesyl pyrophosphate as their substrate and characterization of sqTPSs from plants can be achieved by expression of the genes in heterologous hosts like
Escherichia coli,
Saccharomyces cerevisiae (yeast), or
Nicotiana benthamiana, followed by product analysis using GC-MS [
5,
6,
7]. Expression of sqTPSs has previously been successful in all these systems. However, in yeast it has been found that the pH of the media decreases during growth, which can cause acid-induced rearrangement of unstable compounds [
7].
N. benthamiana are fast-growing plants and expression in this host system offers the advantage of a native plant cell environment along with availability of codons, which are optimal for expression of plant TPSs in general [
5].
Some biosynthesized sesquiterpenes are thermo labile and unstable in acidic conditions [
7,
8]. For example, it has been shown that germacrenes are prone to heat-induced rearrangements [
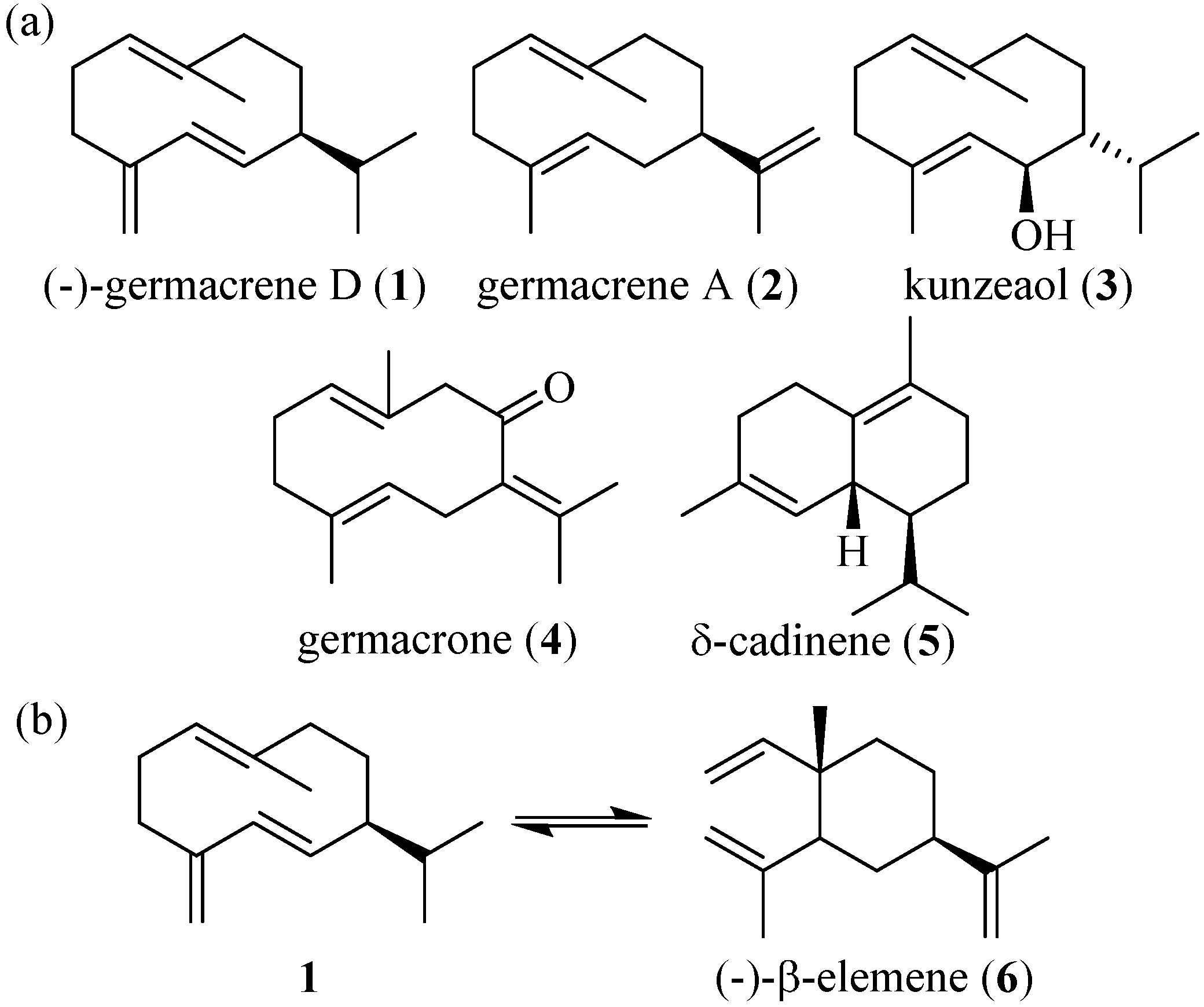
9]. Germacrenes contain an open 10-membered ring structure (
Figure 1). Due to the instability of germacrenes, it was initially suggested that they were an enzymatic intermediate in the biosynthesis of other sesquiterpenes [
10,
11]. It has since been shown that germacrenes are enzymatic end-products themselves, but that they are unstable and rearrange when exposed to high temperatures, acids, or reactive materials [
7,
9]. One such describe rearrangement is the heat-induced Cope rearrangement where the germacrene ring is transformed into the elemene structure (
Figure 1b), but acid-assisted rearrangement has also been described [
9,
12,
13]. The inherent instability of many sesquiterpenes under commonly used GC-MS conditions means their analysis must be carried out with caution [
7,
8].
Head Space-Solid Phase Micro Extraction (HS-SPME) fibers are used in many areas of volatility studies and are an obvious choice for fast screening of volatile sesquiterpenoids on GC-MS [
5,
14,
15,
16,
17]. Continued studies have led to a variety of different fiber coating materials, and combinations thereof, which are optimal for different kinds of volatiles [
18,
19]. SPME fibers often consist of a fused silica core coated with different materials that make them suitable for different analytes. Commercially available SPME fibers can be divided in two groups with regards to the mechanism involved in the interaction between the analytes and the SPME phase,
absorption and
adsorption [
20]. With
absorption, the analytes are extracted by partition in the bulk of the SPME stationary phase; with
adsorption, the analytes physically interact or chemically bind to the surface of the stationary phase.
Figure 1.
(a) (−)-germacrene D (1), germacrene A (2), kunzeaol (3), germacrone (4), and δ-cadinene (5). (b) Cope rearrangement of (−)-germacrene D into (−)-β-elemene (6).
Figure 1.
(a) (−)-germacrene D (1), germacrene A (2), kunzeaol (3), germacrone (4), and δ-cadinene (5). (b) Cope rearrangement of (−)-germacrene D into (−)-β-elemene (6).
Stationary phases that have
absorption properties include polydimethylsiloxane (PDMS) and polyacrylate. The diffusion of small molecules in PDMS is similar to that in organic solvents; this provides a fast diffusion of the analytes into PDMS and an
absorption type of extraction. Diffusion in polyacrylate is significantly lower than for PDMS, but still fast enough to allow for
absorption of the analytes to the stationary phase [
20]. The
adsorption type of fibers contains carboxen (CAR), divinylbenzene (DVB), or a mixture of these two plus PDMS. In the CAR/DVB/PDMS fiber material the primary extracting mechanism is
adsorption to the porous material [
20]. Overall, in these materials, the diffusion is poor and the analytes are retained at the surface by
adsorption to the stationary phase. The pore size of the material is then very important for the capacity of the fiber and when the maximum is reached, there is constant equilibrium with the surrounding phase [
20,
21]. CAR and DVB have a similar surface area, but while the former has an even distribution of macro, meso, and micro pores, the latter is mainly a meso porous material with a moderate amount of macro pores. Macro pores are mainly present on the surface of the material and trap analytes by hydrogen bonding or van der Waals interaction while micro- and meso pores physically trap analytes [
22].
Fibers with PDMS/DVB/CAR are a combination of CAR-PDMS coated with DVB-PDMS. Thus, the larger analytes are retained in the meso and marco pores of the outer DVB layer, while the smaller analytes migrate through this layer and are retained by the micropores in the inner layer of CAR. This enables the fiber to cover a wide range of analytes in the range of 40–275 Da [
8].
In the present study, we systematically compare the effect of different types of HS-SPME fiber coating material and injection temperatures on the rearrangements of sesquiterpenoids during HS-SPME-GC-MS analysis using H2 as the carrier gas. Germacrene D and germacrene A were used as examples of volatile unstable hydrocarbons and kunzeaol as a semi-volatile hydroxylated sesquiterpenoid. A pure standard of germacrone (4), along with the stable sesquiterpene δ-cadinene (5), was also included for comparison. Through this work, we provide examples of thermal and acid rearrangement on various SPME fibers and suggest solutions that can minimize these common problems associated with GC-MS sesquiterpenoid analysis.
2. Experimental Section
2.1. Cloning
The sesquiterpene synthase
TlTPS7414 (KR822706) was found in a trinity transcriptome dataset from
Thapsia laciniata, as previously described [
23]. The full-length sequence was obtained from a cDNA library from
T. laciniata Rouy (syn.
T. villosa [
24]) root material that was collected in early June 2008 just west of Cannes, France (GPS 43.540958, 6.816158), as previously described [
2]. Forward and reverse primers (
Table 1) were designed with USER-overhangs to enable cloning into a USER compatible version of the pEAQ-
HT vector. pEAQ-
HT was kindly provided by George Lomonosonoff (John Innes Research Centre, Norwich, UK) and was modified to harbor a USER cassette [
25]. USER cloning was performed as previously described [
26]. pEAQ-
HT harbors the viral suppressor p19 [
27]. Characterization of
TgSTS1 (JQ290344) and
TgTPS2 (JQ290345) from
Thapsia garganica is previously described [
7].
TgSTS1 and
TgTPS2 were also cloned into pEAQ-
HT.
AtHMGR1 was amplified from an
Arabidopsis thaliana cDNA library.
cfDXS from
Coleus forskohlii was kindly provided by Johan Ranberg-Andersen (University of Copenhagen, Copenhagen, Denmark) [
28].
TpGAS (JF819848) from
Tanacetum parthenium in ImpactVector1.5 was kindly provided by Harro Bouwmeester (University of Wageningen, Wageningen, The Netherlands) [
29].
Table 1.
Overview of primers used.
Table 1.
Overview of primers used.
| Forward Primer | Reverse Primer |
|---|
| TlTPS7414 | 5'-GGCTTAAUATGGCTATGTGTGTTAATTC-3' | 5'-GGTTTAAUTTATACAGGAACAGGGTCC-3' |
| TgSTS1 | 5'-GGCTTAAUATGGCTGTGTATGTTAACTCTACAACTG-3' | 5'-GGTTTAAUTTATAGTGGAATGGGATTTATGAAAG CCGTGGT-3' |
| TgSTS2 | 5'-GGCTTAAUATGGCTGTGTATGTTAACTCTACAACAG-3' | 5'-GGTTTAAUTTATGCTGGAATGGGATTTATGAGA ACCGAGGT-3' |
| AtHMGR1 | 5'-GGCTTAAUATGGATCTCCGTCGGAGGCCTCCTAAAC-3' | 5'-GGTTTAAUTTATGTTGTTGTTGTTGTCGTTGTCGTT GCTCC-3' |
2.2. Expression in Nicotiana benthamiana
Nicotiana benthamiana plants were grown from seeds at 24 °C day/19 °C night for five weeks before infiltration. Transformation of
Agrobacterium tumefaciens followed by infiltration of
N. benthamiana was performed as previously described [
5]. Plants were harvested five days after infiltration.
2.3. Pentane Extraction and HS-SPME Analysis
For pentane extraction, a leaf disc from N. benthamiana expressing TlTPS7414 was crushed in liquid nitrogen and extracted in 1200 µL pentane for 2 h at 23 °C. Samples were centrifuged at 14,000 rpm for 2 min before filtration through a 0.45-µm filter prior to GC-MS analysis.
For the HS-SPME analysis, five different HS-SPME fibers that could be used with an auto-sampler were chosen (see
Table 2). HS-SPME fibers were purchased from Supelco Sigma-Aldrich, Denmark and were selected to represent the different type of coatings available. For these rearrangement experiments, a whole leaf was placed in a 20-mL glass vial and extracted at 60 °C for 20 min with an SPME fiber exposed to the air above the leaf. Immediately hereafter, the SPME fiber was run on the GC-MS.
Extraction of leaves at either 25 °C or 60 °C did not influence the product profile, but higher amounts were obtained at 60 °C, due to the volatility of the compounds. In some tests, extraction at 25 °C did not result in any product detection. Therefore, 60 °C was used for all experiments.
Table 2.
Overview and describtion of the tested fibres, including their product number at Supelco, Sigma-Aldrich.
Table 2.
Overview and describtion of the tested fibres, including their product number at Supelco, Sigma-Aldrich.
| Thickness and Fiber coating | Fiber core | Molecular mass range (Da) | Recommended for | Absorbent/ Adsorbent | Purchasing information |
|---|
| 100 µm PDMS | Fused Silica | 60–275 | volatiles | Absorbent | 57341-U |
| 65 µm PDMS/DVB | Fused Silica | 50–300 | volatiles, amines, nitro-aromatic compounds | Adsorbent | 57345-U |
| 85 µm CAR/PDMS | StableFlex | 30–225 | gases, low molecular weight compounds | Adsorbent | 57295-U |
| 50/30 µm DVB/CAR/PDMS | StableFlex 1cm | 40–275 | volatiles, semi-volatiles, C3-20 | Adsorbent | 57298 |
| Polyacrylate | Fused Silica | 80–300 | polar semi-volatiles | Absorbent | 57294-U |
2.4. GC-MS Analysis
Initially five different injection port temperatures were tested for TlTPS7414 using DVB/CAR/PDMS fibers; 110 °C, 160 °C, 190 °C, 220 °C, and 250 °C. Of these 110 °C, 160 °C, and 250 °C were selected to be tested for all the fibers. Three biological replicates were performed for each injection port temperature combined with the selected types of fibers. This was repeated for TpGAS, TgTPS1 and TgTPS2. A purified germacrone standard, purchased from International Laboratory USA, was included as a further test.
Samples were analyzed on a Shimadzu GCMS-QP2010 plus. The column used was an Agilent HP-5MS UI, fused Silica capillary column 20 m, 0.18 mm diameter × 0.18 µm film thickness, inserted directly into the ion source of the MS. The pressure was kept at 66.7 kPa, giving a column flow of 1 mL/min. The oven temperature was set to 60 °C for 3 min, and then increased to 160 °C with a rate of 7 °C/min. The temperature was further increased to 300 °C with a rate of 50 °C/min and held for 5 min and finally increased to 320 °C with 50 °C/min and maintained for 3 min. The carrier gas was H2 and the ionization electron energy was 70 eV. The ion source temperature was 230 °C with an interface temperature of 280 °C. The total run time was 28.49 min. SPME fibers were conditioned at 240 °C for 20 min after each test.
A C7-C30 standard purchased from Sigma-Aldrich, Denmark was used to calculate retention indices (Ri). All data were analyzed using the Shimadzu software Lab Solutions, GCMS Solutions version 2.50 SU3, with the 2008 libraries provided by NIST and Wiley. TlTPS7414 chromatograms were integrated in the range 10 to 15 min, since this included all detected sesquiterpenes. TgTPS2 chromatograms were integrated in the range 10 to 17 min. TpGAS chromatograms were integrated in the range 11 to 14 min. In a few cases where the separation of peaks could not be done automatically, peak separation was performed manually.
3. Results and Discussion
In order to examine and establish the best conditions for fast and reliable screening of terpene synthase products, the products of
TlTPS7414 (germacrene D synthase),
TpGAS (germacrene A synthase),
TgTPS2 (kunzeaol synthase), and
TgTPS1 (δ-cadinene synthase) were analyzed using four different fibers (
Table 2) and at three different injection port temperatures: 250 °C, 160 °C, and 110 °C. Product release from fibers is correlated with the injection port temperature; therefore decreasing the temperature from 250 °C will also lead to lower levels of product detected by the GC-MS. In initial testing, a temperature of 110 °C was found to result in reasonable product detection for
TlTPS7414. Between 110 °C and 250 °C, three additional temperatures were tested: 160 °C, 190 °C, and 220 °C; however, there was little change observed in the product profile between these three temperatures (
Supplementary Figure 1).
A polyacrylate fiber was also included in initial tests but did not provide stable results in our assay due to oxidation of the fiber material in the injection port. At low temperatures (
i.e., 110 °C) the polyacrylate polymer is a solid crystalline coating and therefore the diffusion of the analytes in the fiber is negligible. The fiber turns into liquid at desorption temperatures (
i.e., 250 °C). It was observed that exposure to such temperatures resulted in oxidation of the fiber, suggesting the presence of oxygen in the injection port [
30]. Thus, polyacrylate was not tested further.
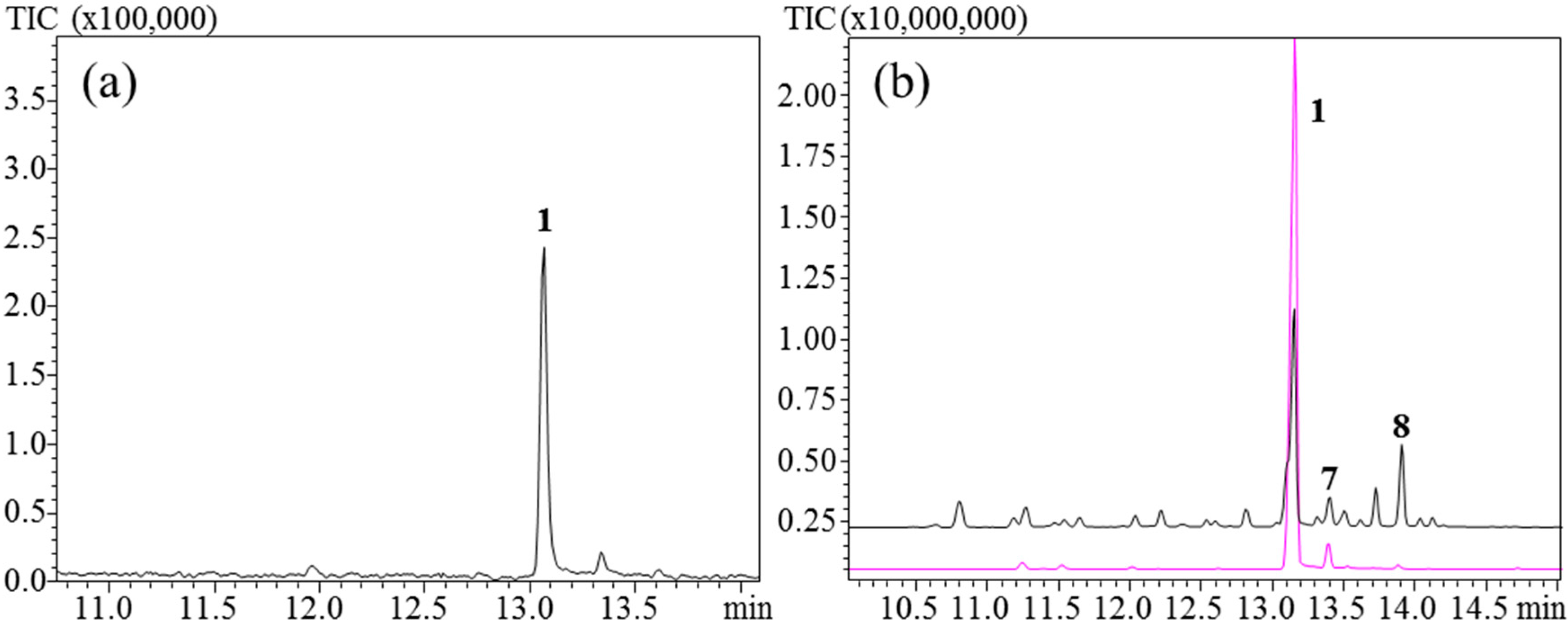
3.1. Initial Tests and Characterization of TlTPS7414
To elucidate the product profile of
TlTPS7414, this was expressed in
Nicotiana benthamiana. The resulting products were analyzed using HS-SPME and pentane extraction of the leaves. In the pentane extraction (−)-germacrene D (
1) was found to be the major product and bicyclogermacrene (
7) a minor product (
Figure 2a). Thus,
TlTPS7414 is a (−)-germacrene D synthase. This product profile was also observed with HS-SPME analysis when using a PDMS fiber and injection port temperature at 160 °C (
Figure 2b). To enhance production levels,
TlTPS7414 was co-expressed with
AtHMGR or
CfDXS, which are upstream precursor genes of the mevalonate (MVA) and the 2-
C-methyl-D-erythritol 4-phosphate (MEP) pathway, respectively. HMGR, which is the third enzyme in the MVA pathway, is tightly regulated in plants and has been shown to be a rate-limiting step [
31,
32]. Cross-talk has been shown to occur between the MVA and MEP pathways. Therefore, DXS, which is a rate-limiting enzyme in the MEP pathway, was also tested [
33,
34]. The product profile and ratios were identical whether
AtHMGR or
CfDXS were used, and for both enzymes, the product levels were higher than without the co-expression of these upstream genes (
Supplementary Figure 2). The data presented here are from co-expression of
TlTPS7414 and
CfDXS.
TpGAS,
TgTPS1, and
TgTPS2 were also co-expressed with
CfDXS.
Figure 2.
TlTPS7414 expressed in N. benthamiana (a) Pentane extraction; (b) extraction using PDMS (lower) and PDMS/DVB/CAR (upper) fibers and an injection port temperature of 160 °C. 7 was not positively identified in the pentane extract.
Figure 2.
TlTPS7414 expressed in N. benthamiana (a) Pentane extraction; (b) extraction using PDMS (lower) and PDMS/DVB/CAR (upper) fibers and an injection port temperature of 160 °C. 7 was not positively identified in the pentane extract.
3.2. Comparison of SPME Fibers Using TlTPS7414 – (-)-Germacrene D Synthase
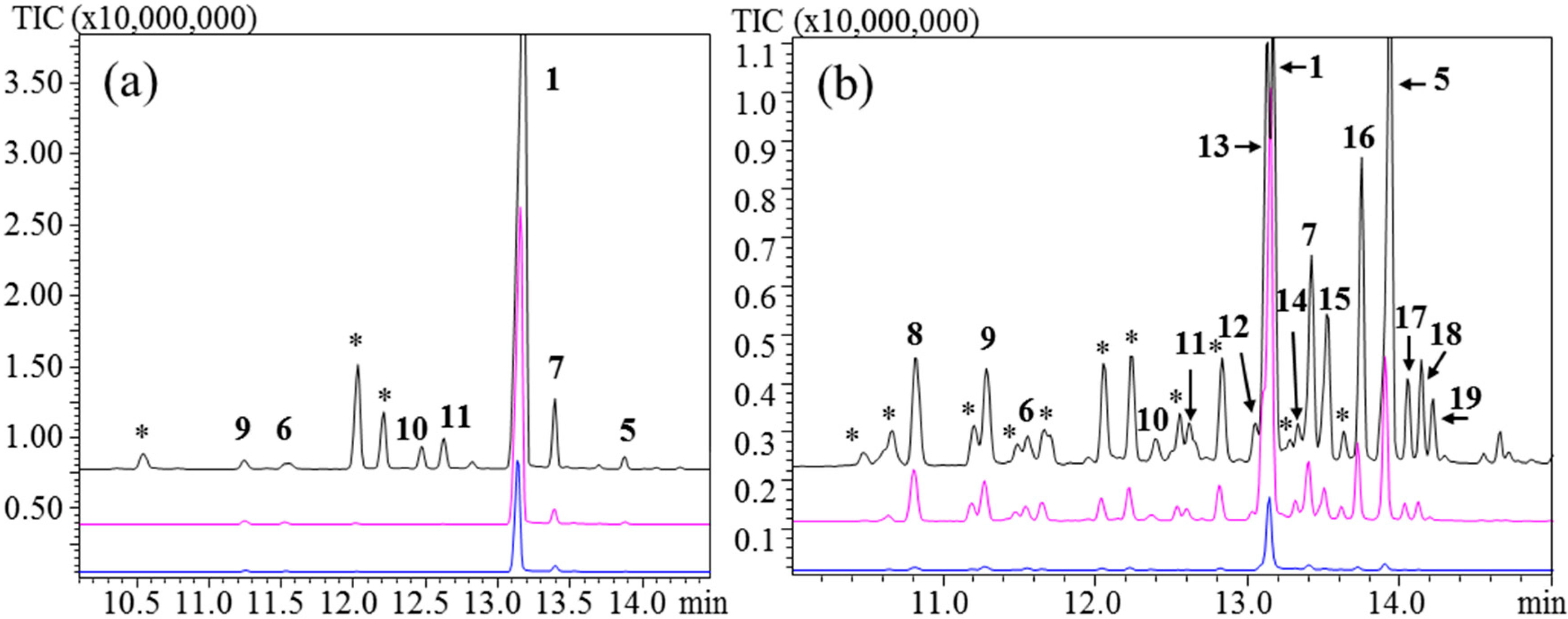
As a first assessment, a PDMS/DVB/CAR fiber was chosen together with an injection port temperature of 250 °C. The resulting chromatogram from
TlTPS7414 revealed over 30 sesquiterpene compounds where the major peak identified was δ-cadinene (
5) (
Figure 3b). Compounds with a relative area of >0.9% are listed in
Table 3. As described above, germacrenes are thermo labile and prone to acid rearrangement [
9]. The temperature of the injection port and acidic conditions can explain the many rearrangement products observed in the HS-SPME experiment at 250 °C with the PDMS/DVB/CAR fiber. This phenomenon has previously been described with similar fiber materials [
8,
35]. At 160 °C
5 was still a notable product, although
1 was the major product. The product profile had improved at 110 °C to show only
1 as the significant product along with some minor products.
Figure 3.
TlTPS7414 expressed in N. benthamiana. Injection port temperatures; 110 °C (lower), 160 °C (middle), and 250 °C (upper) (a) PDMS fiber; (b) PDMS/DVB/CAR fiber. Unknown sesquiterpene products are marked with (*).
Figure 3.
TlTPS7414 expressed in N. benthamiana. Injection port temperatures; 110 °C (lower), 160 °C (middle), and 250 °C (upper) (a) PDMS fiber; (b) PDMS/DVB/CAR fiber. Unknown sesquiterpene products are marked with (*).
Table 3.
Detected compounds of expression of
TlTPS7414 in
N. benthamiana with injection temperature at 250 °C. Area % is the given area compared to total sesquiterpenoid area. R
t: retention time, R
i: retention index. Literature values of R
i are all from Adams [
36]. NI = Non-identified, Litt = Literature.
Table 3.
Detected compounds of expression of TlTPS7414 in N. benthamiana with injection temperature at 250 °C. Area % is the given area compared to total sesquiterpenoid area. Rt: retention time, Ri: retention index. Literature values of Ri are all from Adams [36]. NI = Non-identified, Litt = Literature.
| | DVB/CAR/PDMS | PDMS/DVB | CAR/PDMS | PDMS | |
|---|
| Compounds | Area % | Rt | Ri | Area % | Rt | Ri | Area % | Rt | Ri | Area % | Rt | Ri | Litt. Ri |
|---|
| | NI | 1.0 | 10.48 | 1331 | | | | | | | | | | |
| | NI | | | | | | | | | | 1.7 | 10.54 | 1335 | |
| | NI | | | | | | | 1.2 | 10.59 | 1338 | | | | |
| | NI | 1.7 | 10.64 | 1340 | | | | | | | | | | |
| 8 | α-cubebene | 3.6 | 10.82 | 1350 | 2.9 | 10.79 | 1348 | 3.2 | 10.79 | 1348 | | | | 1345 |
| | NI | 1.6 | 11.20 | 1370 | 2.0 | 11.17 | 1369 | 1.9 | 11.18 | 1369 | | | | |
| 9 | α-Copaene | 3.9 | 11.29 | 1375 | 4.6 | 11.26 | 1373 | 5.7 | 11.26 | 1373 | 1.0 | 11.24 | 1372 | 1376 |
| | NI | 1.0 | 11.48 | 1385 | | | | | | | | | | |
| 6 | (−)-β-elemene | 1.0 | 11.56 | 1389 | | | | 1.4 | 11.53 | 1388 | 1.1 | 11.54 | 1389 | 1391 |
| | NI | 1.6 | 11.66 | 1395 | 1.2 | 11.63 | 1393 | 1.1 | 11.64 | 1393 | | | | |
| | NI | 3.2 | 12.06 | 1417 | 2.3 | 12.03 | 1415 | 2.9 | 12.03 | 1416 | 8.3 | 12.03 | 1415 | |
| | NI | 3.3 | 12.24 | 1427 | 2.8 | 12.21 | 1426 | 3.3 | 12.22 | 1426 | 4.3 | 12.21 | 1425 | |
| 10 | Aromadendrene | 1.6 | 12.40 | 1436 | 1.1 | 12.37 | 1435 | 1.6 | 12.38 | 1435 | 1.8 | 12.47 | 1441 | 1440 |
| | NI | 1.8 | 12.56 | 1445 | 1.9 | 12.53 | 1444 | 1.6 | 12.53 | 1444 | | | | |
| 11 | ε-Muurolene | 1.7 | 12.62 | 1449 | 1.7 | 12.59 | 1447 | 1.9 | 12.60 | 1447 | 2.5 | 12.63 | 1449 | 1446 |
| | NI | 3.5 | 12.84 | 1461 | 4.2 | 12.80 | 1459 | 4.3 | 12.81 | 1460 | | | | |
| 12 | β-Cadinene | | | | | | | 1.8 | 13.02 | 1472 | | | | 1473 |
| 13 | γ-Muurolene | | | | 13.5 | 13.10 | 1476 | 8.1 | 13.09 | 1476 | | | | 1477 |
| 1 | (−)-Germacrene D | 13.8 | 13.14 | 1478 | 8.3 | 13.14 | 1478 | 7.3 | 13.14 | 1478 | 74.2 | 13.18 | 1480 | 1480 |
| | NI | 8.7 | 13.18 | 1481 | | | | | | | | | | |
| | NI | 1.0 | 13.28 | 1486 | | | | | | | | | | |
| 14 | (+)-Epi-bicycloses-quiphellandrene | 1.2 | 13.33 | 1489 | | | | 1.6 | 13.31 | 1488 | | | | 1482 |
| 7 | Bicyclogermacrene | 7.1 | 13.43 | 1495 | 7.5 | 13.40 | 1493 | 5.9 | 13.40 | 1493 | 4.6 | 13.39 | 1493 | 1494 |
| 15 | α-muurolene | 5.4 | 13.54 | 1501 | 7.1 | 13.50 | 1499 | 6.7 | 13.50 | 1499 | | | | 1499 |
| | NI | 1.1 | 13.64 | 1507 | 2.4 | 13.61 | 1505 | | | | | | | |
| 16 | (+)-α-amorphene | 8.9 | 13.76 | 1514 | 9.2 | 13.73 | 1512 | 7.4 | 13.73 | 1512 | | | | 1506 |
| 5 | (+)-δ-cadinene | 18.1 | 13.95 | 1526 | 20.7 | 13.92 | 1524 | 23.9 | 13.91 | 1523 | 1.0 | 13.88 | 1521 | 1522 |
| 17 | Cadine-1,4-diene | 2.2 | 14.07 | 1533 | 3.1 | 14.03 | 1531 | 1.7 | 14.04 | 1531 | | | | 1532 |
| 18 | (−)-α-Cadinene | 2.8 | 14.16 | 1538 | 3.1 | 14.12 | 1536 | 2.6 | 14.12 | 1536 | | | | 1538 |
| 19 | α-Calacorene | 1.5 | 14.23 | 1542 | | | | 3.6 | 14.20 | 1541 | | | | 1548 |
The product profiles obtained when using CAR/PDMS and PDMS/DVB fibers were similar to that of DVB/CAR/PDMS at 250 °C. A tendency towards higher levels of rearrangement products at 160 °C and 110 °C was observed for these fiber types (
Table 4,
Supplementary Figures 3 and 4).
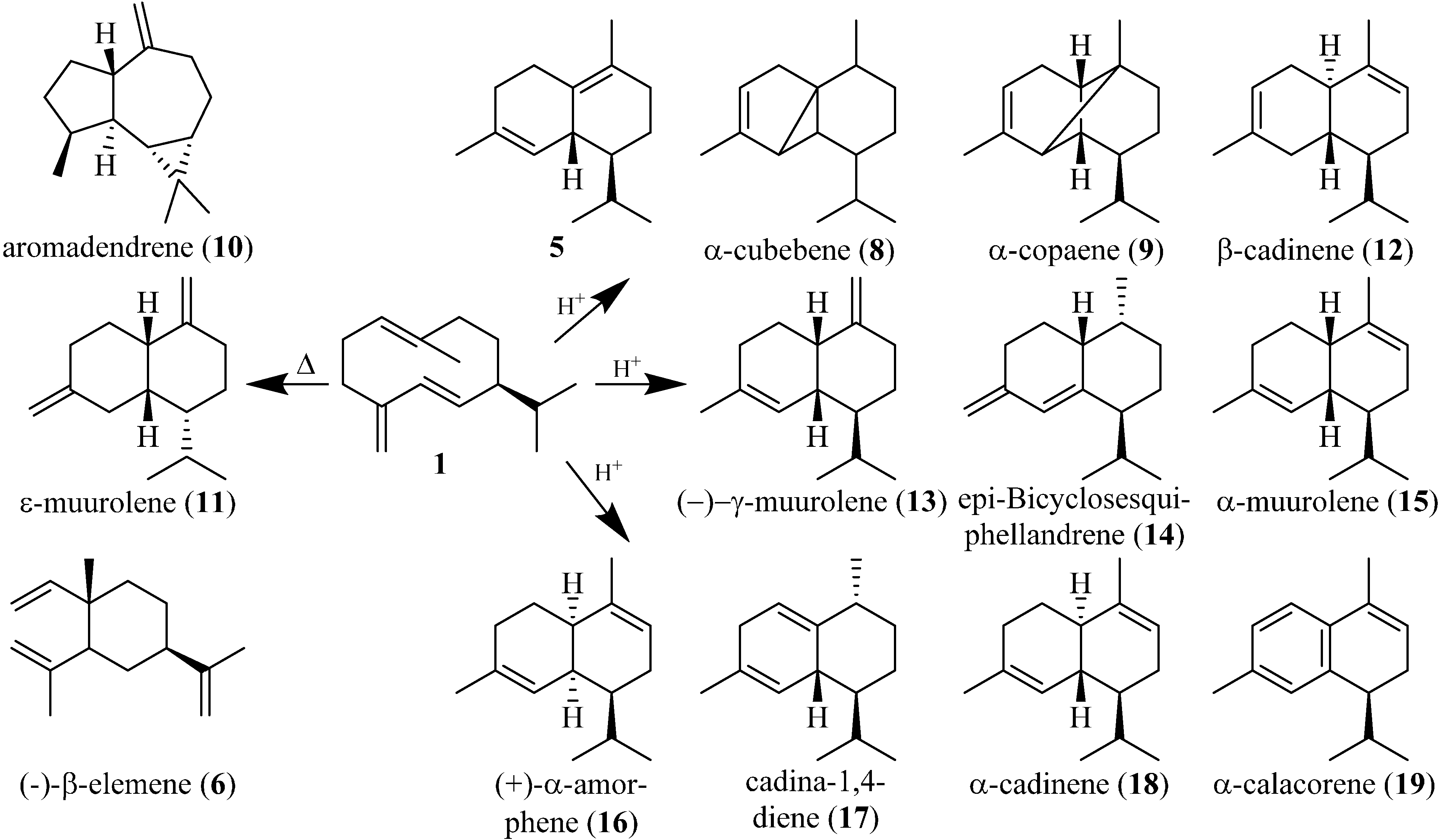
The rearrangement products of DVB/CAR/PDMS, CAR/PDMS, and PDMS/DVB at 250 °C shown in
Table 3 were identified based on retention indices and GC-MS libraries. The compounds are mainly of the cadinane type of sesquiterpenes, which includes cadinenes, muurolenes, and amorphenes. Compared with previous publications, these rearrangement products not only show the expected heat-induced rearrangement (Cope rearrangement) products, but also products that are caused by acid induced cyclizations (
Figure 4) [
13,
37]. The ability of the DVB and CAR material to
adsorb H
2O to the surface and the presence of H
2O in the carrier gas H
2 create a weak acidic environment that leads to the acid-induced rearrangements observed. (+)-δ-cadinene (
5) offers the most energy favorable composition of all the described acid-induced rearrangement products, thus
5 is chemically favored over the others and consequently is produced at the highest level. Thus, occurrence of
5 indicates acid-induced cyclizations, whereas the occurrence of β-elemene (
6) indicates heat-induced Cope rearrangements.
Using the PDMS material resulted in the lowest amount of rearrangement products at all tested temperatures (
i.e., 110 °C, 160 °C, and 250 °C) of the four fibers analyzed. PDMS is an
absorption-based coating with a fused silica core. The PDMS fiber was the only fiber where
1 was obtained as the major product at all the tested injection port temperatures (
Figure 3a). Multiple products were detected at all tested temperatures, although at 160 °C and 110 °C the relative amount of
1 was ~90%.
Figure 4.
Rearrangement products of germacrene D (1). The left side shows the identified heat-induced rearrangement products. The right side shows the identified acid-induced rearrangement products.
Figure 4.
Rearrangement products of germacrene D (1). The left side shows the identified heat-induced rearrangement products. The right side shows the identified acid-induced rearrangement products.
At an injection port temperature of 250 °C the PDMS fiber showed
1 as the major compound, despite reports that germacrene D is only stable up to 180 °C [
13]. The short time in the injection port did not allow for full rearrangement of
1; thus, when high amounts of
1 are injected, it is still found in the chromatograms, as seen in
Figure 3. The rearrangement profile detected with the PDMS fiber is mainly related to heat-induced rearrangements. Acid-induced rearrangement can possibly occur, but not to the same levels as seen for the
adsorbent materials. PDMS is very hydrophobic and H
2O does not
absorb to the material. This is one of the main arguments for the use of this fiber. At 160 °C the levels of rearrangement products are low and the detected levels of compounds are still high.
Of the four fiber materials and different temperatures tested here, the results clearly show that PDMS fiber and an injection port temperature of 160 °C provide the best conditions for analysis of germacrene D synthases (
Table 4).
Table 4.
Comparison of germacrene D (1) levels (% of total sesquiterpenoids) using different fiber materials. SD is the standard deviation of at least three biological replicates.
Table 4.
Comparison of germacrene D (1) levels (% of total sesquiterpenoids) using different fiber materials. SD is the standard deviation of at least three biological replicates.
| Temperature | 110 °C | SD | 160 °C | SD | 250 °C | SD |
|---|
| Fiber | | | | | | |
| PDMS/DVB/CAR | 55.6 | 4.7 | 30.3 | 3.5 | 8.1 | 0.6 |
| PDMS/DVB | 29.5 | 6.4 | 13.1 | 3.2 | 6.9 | 0.7 |
| PDMS/CAR | 46.1 | 9.8 | 18.7 | 5.3 | 6.3 | 0.5 |
| PDMS | 88.4 | 2.0 | 90.5 | 2.7 | 66.9 | 4.6 |
3.3. Comparison of SPME Fibers Using TpGAS -Germacrene A Synthase
Germacrene A was included as an example of a germacrene with a different positioning of double bonds compared to germacrene D (
Figure 1).
TpGAS has previously been described as a germacrene A synthase [
29]. Expression of
TpGAS in
N. benthamiana followed by extraction using HS-SPMS and analysis on GC-MS with the different fiber materials PDMS, PDMS/DVB, PDMS/CAR, and PDMS/CAR/DVB at an injection port temperature of 250 °C only resulted in rearrangement products and did not yield any germacrene A (
2) (
Figure 5,
Supplementary Figures 5 and 6).
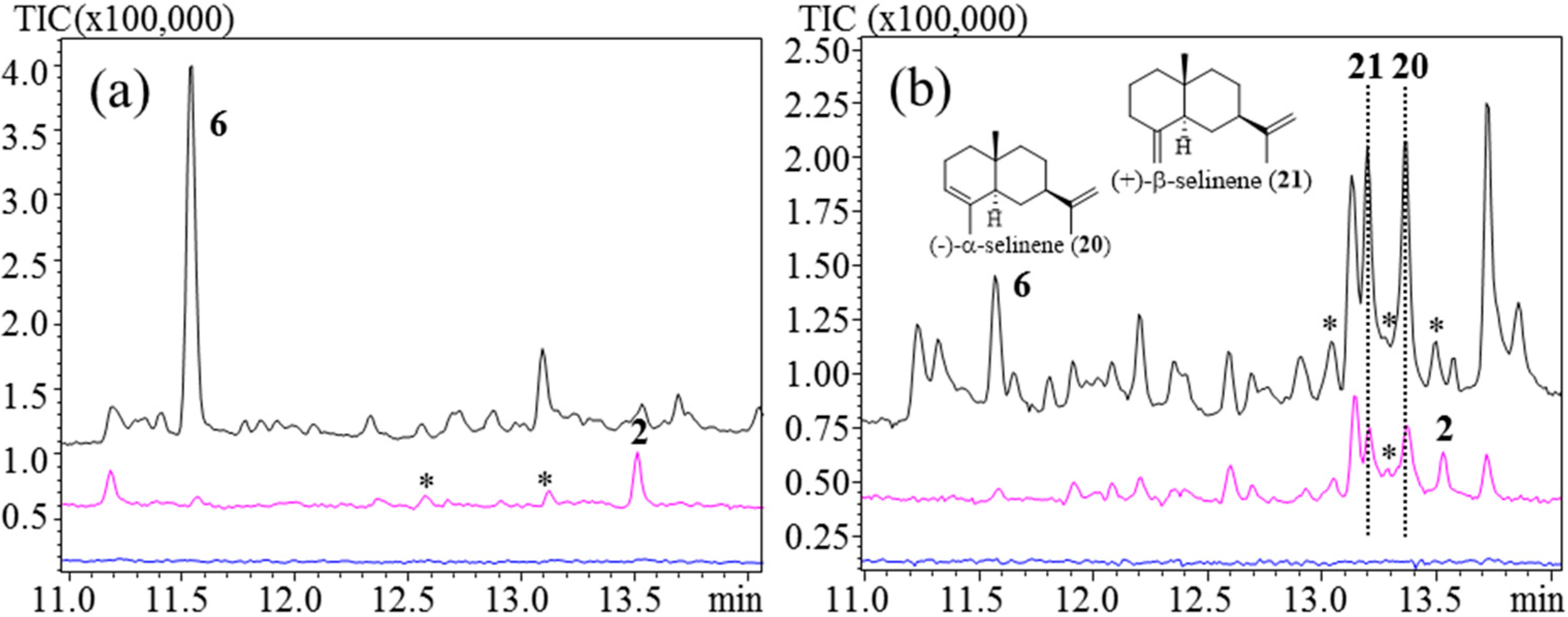
With PDMS/DVB, PDMS/CAR and PDMS/CAR/DVB at 250 °C the identified major products were (−)-α-selinene (
20) and (+)-β-selinene (
21), with
6 as a minor product and some minor unknown sesquiterpenoids (
Figure 5b,
Supplementary Figures 5 and 6).
20 and
21 are the eudesmane type of sesquiterpenes and are acid-induced rearrangement products of
2, as has also been detected previously [
13].
6 is the result of thermal Cope rearrangement of
2, as was described above [
38]. At 160 °C
2 was not detected using PDMS/DVB and PDMS/CAR (
Supplementary Figures 5 and 6). For PDMS/DVB/CAR
2 was detected along with
20,
21, and one unknown sesquiterpenoid (
Figure 5b).
For PDMS,
6 was detected as the major product at 250 °C.
2 was the major product at 160 °C followed by a few minor unknown products (
Figure 5a).
6 was not identified at 160 °C for any fibers (
Figure 5,
Supplementary Figures 5 and 6), which correlates with germacrenes in general being stable at lower temperatures. At 110 °C there were no products detected, most likely owing to the lower quantity of product produced by
TpGAS compared to
TlTPS7414. The product profile of
TpGAS was less complex than
TlTPS7414 for all four types of fibers, mainly due to the composition of the double bonds in
2.
Figure 5.
TpGAS, injection port temperatures: 110 °C (lower), 160 °C (middle), and 250 °C (upper). (a) PDMS; (b) PDMS/DVB/CAR. Unknown sesquiterpene products are marked with (*).
Figure 5.
TpGAS, injection port temperatures: 110 °C (lower), 160 °C (middle), and 250 °C (upper). (a) PDMS; (b) PDMS/DVB/CAR. Unknown sesquiterpene products are marked with (*).
3.4. Comparison of SPME Fibers Using TgTPS2–Kunzeaol Synthase
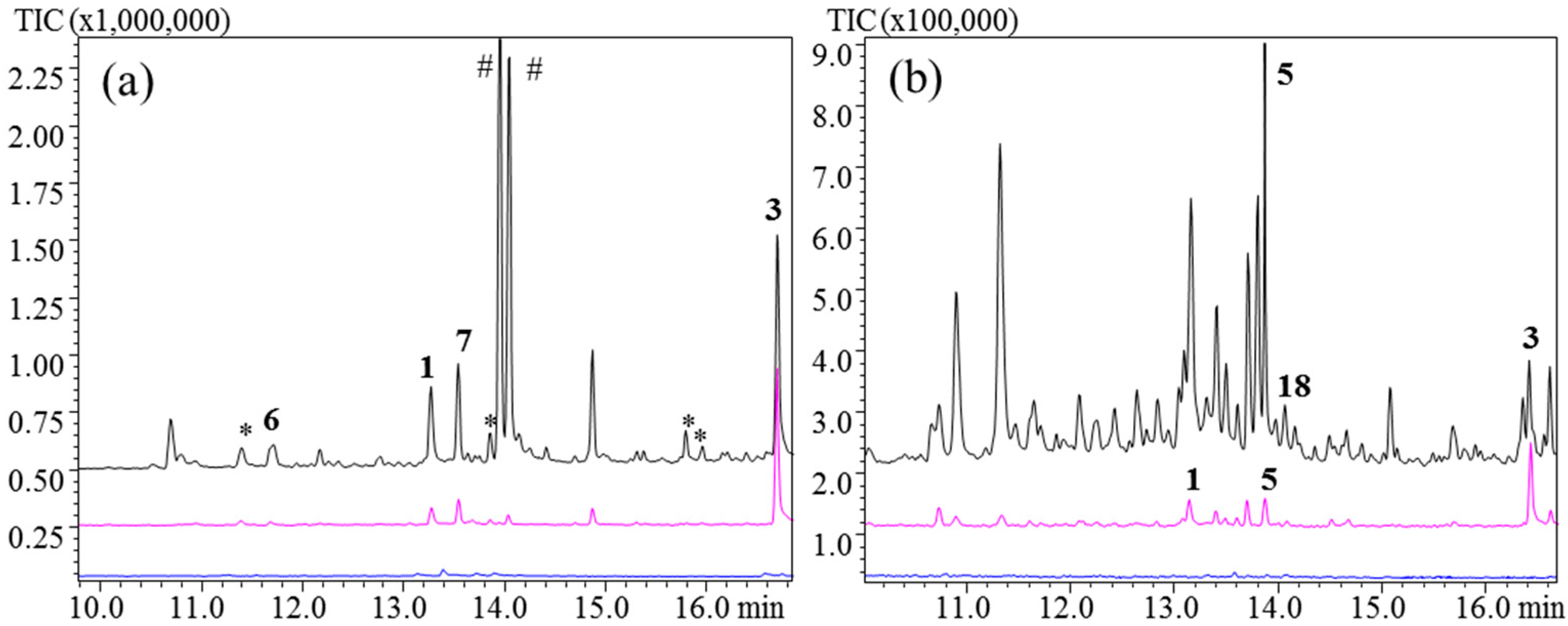
To compare hydrocarbon and hydroxylated germacrenes using HS-SPME we included kunzeaol. The main product of
TgTPS2 when expressed in
S. cerevisiae has previously been shown to be kunzeaol (
3), with the two minor products being
7 and
1 [
7]. In that study,
TgTPS2 expressing
S. cerevisiae was grown in un-buffered media, resulting in a pH drop to 3 after 84 h [
7]. Under these conditions,
3 was found to undergo acid-induced rearrangement to
5, α-cadinol and τ-muurolol, with δ-cadinene being the major product [
39].
TgTPS2 was expressed in
N. benthamiana. With an injection port temperature of 110 °C no products were observed for the PDMS/DVB, PDMS/CAR, and PDMS/CAR/DVB fibers (
Figure 6b,
Supplementary Figures 7 and 8). A few minor products could be observed with the PDMS fiber (
Figure 6a). The hydroxyl group at C6 of
3 reduces the volatility compared to hydrocarbons with the same number of carbons. This can explain why a temperature of 110 °C was not sufficient to desorb
3 from the fiber.
3 was generally not detected at 250 °C using PDMS/DVB, PDMS/CAR, and PDMS/CAR/DVB fibers; however, up to 30 other sesquiterpene rearrangement products could be detected (
Figure 6b,
Supplementary Figures 7 and 8). At 160 °C PDMS/DVB and PDMS/CAR fibers were still not suited for analysis of
3 with only a very low signal detected.
3 could be detected when PDMS/CAR/DVB fibers were used at 250 °C and 160 °C, but
5 and other sesquiterpenoids (
Figure 6) were present in high amounts. The majority of the observed rearrangement products are acid-induced and are similar to what was previously described [
7].
PDMS fibers yielded
3, at 250 °C; however, the major products were found to be two hydroxylated sesquiterpenoids (
Figure 6a). These two sesquiterpenoids might be heat-induced Cope rearrangement products of
3, but the structure could not be verified by GC-MS. At 160 °C
3 was the major product;
1 and
7 were detected as the two other biosynthetic products. At 160 °C the hydroxylated products had decreased to become very minor products.
These findings suggest that heat-induced Cope rearrangements of
3 might be the predominant rearrangement products using the PDMS fibers, whereas the acid-induced rearrangements that lead to
5 are predominant on the other three fibers. This result is consistent with the finding that
3 is not detected in hydro-distillated oil samples from
Kunzea leaves, but can be detected when gentle solvent extraction is used [
39].
Figure 6.
TgTPS2. Injection port temperatures: 110 °C, 160 °C, and 250 °C. (a) PDMS; (b) PDMS/DVB/CAR. Unknown sesquiterpene products are marked with *. Two hydroxylated unknown sesquiterpenoids are marked with #.
Figure 6.
TgTPS2. Injection port temperatures: 110 °C, 160 °C, and 250 °C. (a) PDMS; (b) PDMS/DVB/CAR. Unknown sesquiterpene products are marked with *. Two hydroxylated unknown sesquiterpenoids are marked with #.
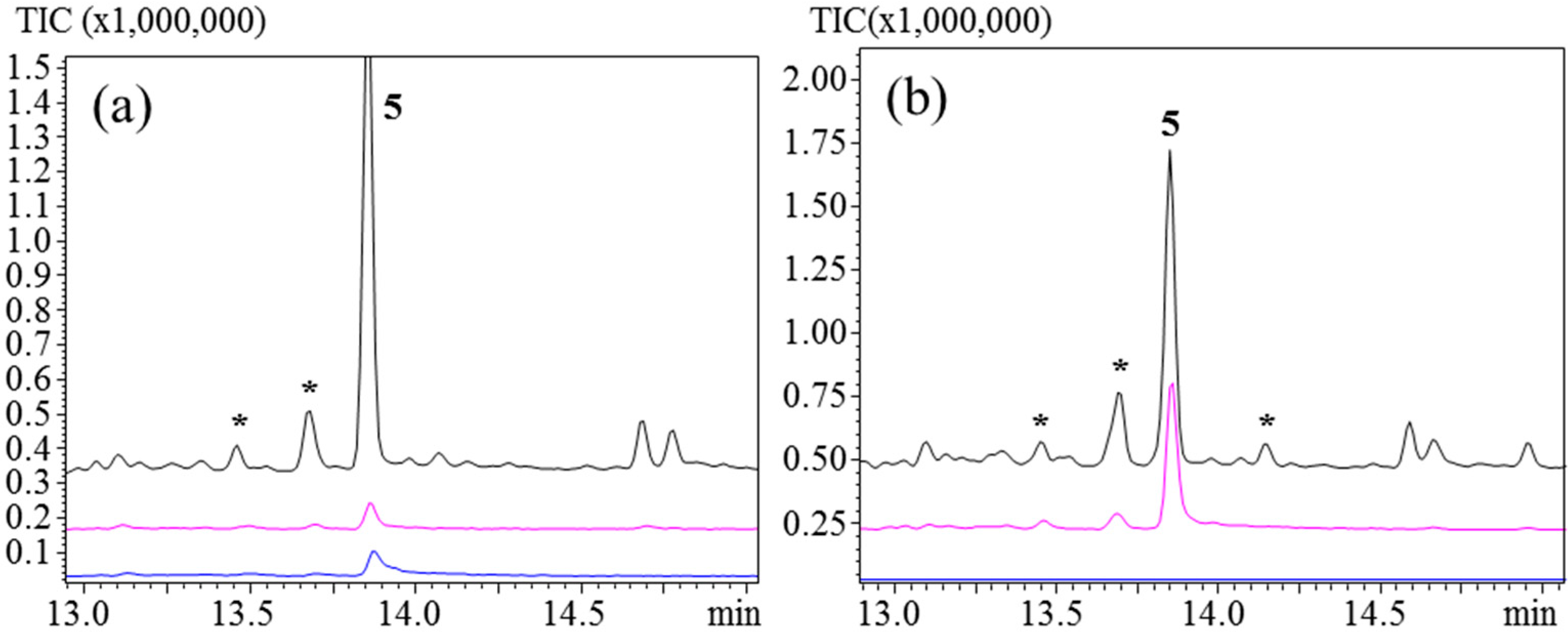
3.5. Comparison of SPME Fibers Using TgTPS1–δ-Cadinene Synthase
As an example of an sqTPS that produces a stable sesquiterpene we included
TgTPS1.
TgTPS1 was previously expressed and characterized in
S. cerevisiae [
7] and found to be a δ-cadinene (
5) synthase. The expression of
TgTPS1 in
N.
benthamiana using PDMS and PDMS/CAR/DVB also resulted in the production of
5, which was the major product detected, regardless of fiber used or injection port temperature (
Figure 7). Minor products seen in the chromatograms could be due to rearrangements of uncharacterized minor biosynthetic products. Therefore, detection of
5 does not always point towards acid rearrangement but is also an enzymatic product itself.
Figure 7.
TgTPS1, injection port temperatures: 110 °C, 160 °C, and 250 °C. (a) PDMS; (b) PDMS/DVB/CAR. Unknown sesquiterpene products are marked with (*).
Figure 7.
TgTPS1, injection port temperatures: 110 °C, 160 °C, and 250 °C. (a) PDMS; (b) PDMS/DVB/CAR. Unknown sesquiterpene products are marked with (*).
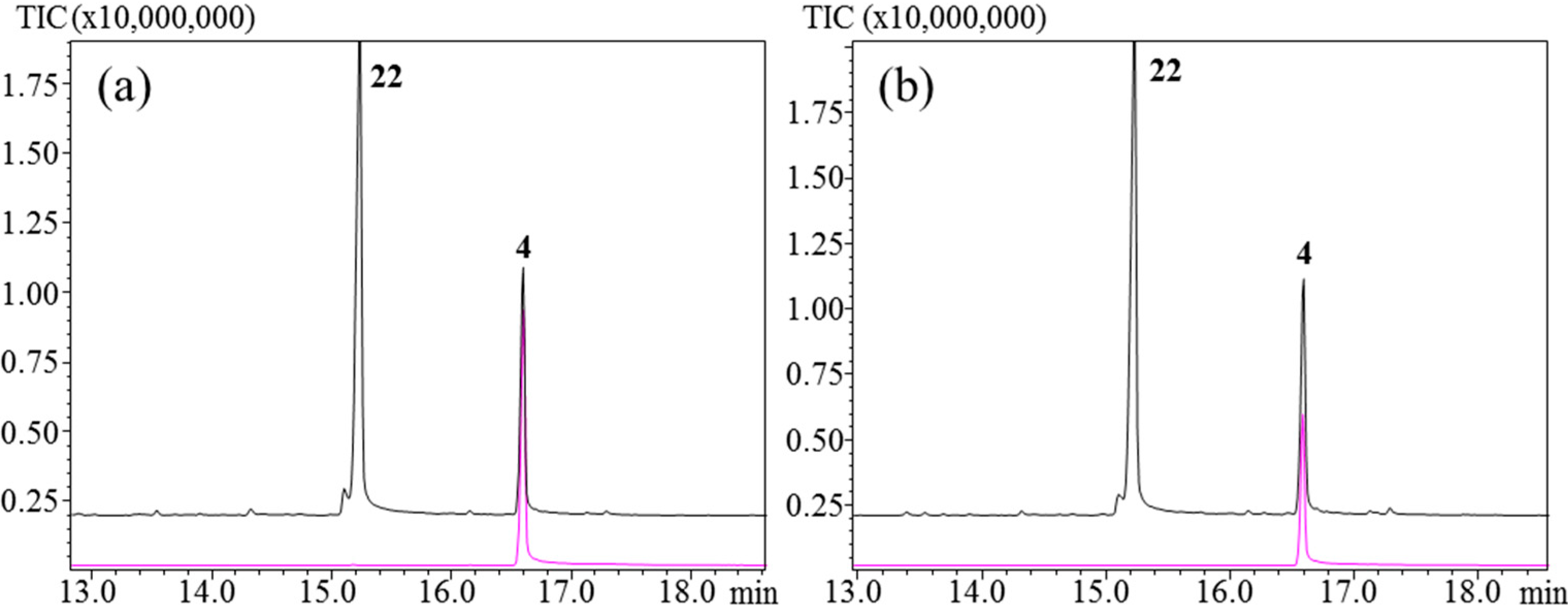
3.6. Comparison of SPME Fibers Using Germacrone (4)
Germacrone (
4) is a germacrene backbone with a ketone group attached.
4 was analyzed with the four fibers at 160 °C and 250 °C (
Figure 8).
4 has previously been described as being stable at acidic pH [
40]. For the analysis of four fibers performed at 250 °C, only two major products,
4 and β-elemenone (
22), and one minor product were detected. This profile arises from a heat-induced Cope rearrangement of
4 to
22, as previously described [
40]. At 160 °C
4 was the sole product.
4 was a pure standard and the high amount of the compound injected into the GC-MS could account for the presence of
4 and not only β-elemenone at 250 °C.
Figure 8.
Germacrone. Injection port temperatures: 160°C and 250°C. (a) PDMS; (b) PDMS/DVB/CAR.
Figure 8.
Germacrone. Injection port temperatures: 160°C and 250°C. (a) PDMS; (b) PDMS/DVB/CAR.
4. Conclusions
Choice of fiber and injection port temperature is important to understand when analyzing unstable compounds like germacrenes. Using H2 as a carrier gas is a cheap alternative to He. However, the influence of carrier gas on the rearrangements needs further studies, as He should be a more inert carrier gas then H2.
The results presented here show that the absorbent material PDMS is much less likely to support acid-induced rearrangement compared to the tested adsorbent material PDMS/DVB, PDMS/CAR, and PDMS/CAR/DVB. This especially becomes apparent with an injection temperature 160 °C, where for the PDMS the germacrenes were the major product, and for the adsorbent materials, several other products were found.
This suggests that unless H2O can be eliminated during the analysis process, one should utilize the absorbent and hydrophobic material PDSM for analysis of sesquiterpenes. Thus, for fast screening of sesquiterpene synthases using expression systems such as N. benthamiana, one should utilize PDMS fibers and an injection temperature of maximum 160 °C for the HS-SPME-GC-MS analysis of the enzymatic products. The use of injection ports with the possibility of ramping the temperature should also be considered, as with this kind of injector ports the sample can be injected at low temperatures to avoid thermal degradations of labile compounds. However, the strength of this technology requires further studies.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






