
Intestinal Glucuronidation, Prior to Hepatic Glucuronidation, Plays an Important Role in the Low Circulating Levels of Calycosin

Abstract
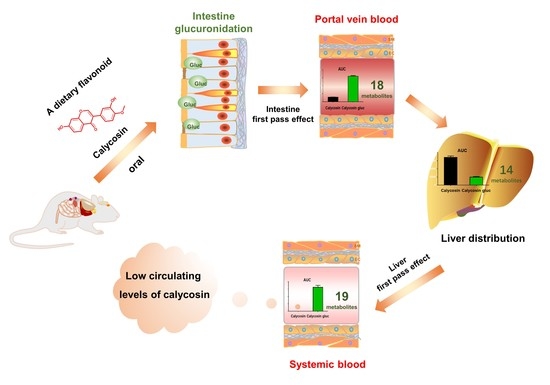
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Instrumentation
2.3. Animal Experiment
2.4. Sample Preparation
2.4.1. Sample Preparation for HPLC-Q-TOF Analysis
2.4.2. Sample Preparation for HPLC-MS/MS Analysis
2.5. Data Analysis
3. Results
3.1. Qualitative Analysis of Calycosin and Its Metabolites In Vivo
3.1.1. M0 and Its Characteristic Fragmentation Patterns
3.1.2. M12 and Its Characteristic Fragmentation Patterns
3.1.3. M15 and Its Characteristic Fragmentation Patterns
3.1.4. M21 and Its Characteristic Fragmentation Patterns
3.2. Quantitative Analysis of Calycosin and Its Metabolites In Vivo
3.2.1. Semi-Quantitative Analysis of Calycosin and Its Metabolites In Vivo
3.2.2. Quantitative Analysis of Calycosin and Calycosin Glucuronides In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Zhang, L.J.; Liu, H.K.; Hsiao, P.C.; Kuo, L.M.; Lee, I.J.; Wu, T.S.; Chiou, W.F.; Kuo, Y.H. New isoflavonoid glycosides and related constituents from astragali radix (Astragalus membranaceus) and their inhibitory activity on nitric oxide production. J. Agric. Food Chem. 2011, 59, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Available online: www.nhc.gov.cn/cms-search/xxgk/getManuscriptXxgk.htm?id=3bcf8b4d12e34b11bfc6e5404b6e74a (accessed on 7 August 2021).
- Yu, Q.T.; Qi, L.W.; Li, P.; Yi, L.; Zhao, J.; Bi, Z. Determination of seventeen main flavonoids and saponins in the medicinal plant Huang-qi (Radix astragali) by HPLC-DAD-ELSD. J. Sep. Sci. 2007, 30, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Q.; Zhuang, Y.; Pan, J.C.; Wang, H.B.; Li, H.; Yu, Y.; Wang, D.Q. Investigation of effects and mechanisms of total flavonoids of Astragalus and calycosin on human erythroleukemia cells. Oxid. Med. Cell Longev. 2012, 2012, 209843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.P.; Wei, G.M.; Li, Z.; Liu, Y.H.; Wang, Z.D. Flavonoids from the Roots of Astragalus membranaceus (Fisch.) Bge. Prevent Development of Diabetic Nephropathy In Vitro. Lat. Am. J. Pharm. 2014, 33, 339–343. [Google Scholar]
- Chen, L.Y.; Li, Z.X.; Tang, Y.H.; Cui, X.L.; Luo, R.H.; Guo, S.S.; Zheng, Y.T.; Huang, C.G. Isolation, identification and antiviral activities of metabolites of calycosin-7-O-beta-D-glucopyranoside. J. Pharm. Biomed. Anal. 2011, 56, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Meng, Q.; Wang, C.Y.; Liu, Q.; Sun, H.J.; Huo, X.K.; Sun, P.Y.; Yang, X.B.; Peng, J.Y.; Liu, K. Protective effects of calycosin against CCl4-induced liver injury with activation of FXR and STAT3 in mice. Pharm. Res. 2015, 32, 538–548. [Google Scholar] [CrossRef]
- Deng, T.; Liu, J.; Zhang, M.M.; Wang, Y.X.; Zhu, G.N.; Wang, J.J. Inhibition effect of phytoestrogen calycosin on TGF-beta1-induced hepatic stellate cell activation, proliferation, and migration via estrogen receptor beta. Can. J. Physiol. Pharmacol. 2018, 96, 1268–1275. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, J.Z.; Liu, W.H.; Liu, N.N.; Fu, X.Q.; Kwan, H.Y.; Liu, S.J.; Liu, B.R.; Zhang, S.W.; Yu, Z.L.; et al. Calycosin inhibits oxidative stress-induced cardiomyocyte apoptosis via activating estrogen receptor-alpha/beta. Bioorg. Med. Chem. Lett. 2016, 26, 181–185. [Google Scholar] [CrossRef]
- Su, X.H.; Huang, Q.C.; Chen, J.Y.; Wang, M.J.; Pan, H.D.; Wang, R.; Zhou, H.; Zhou, Z.Q.; Liu, J.; Yang, F.; et al. Calycosin suppresses expression of pro-inflammatory cytokines via the activation of p62/Nrf2-linked heme oxygenase 1 in rheumatoid arthritis synovial fibroblasts. Pharmacol. Res. 2016, 113, 695–704. [Google Scholar] [CrossRef]
- Duan, X.P.; Meng, Q.; Wang, C.Y.; Liu, Z.H.; Sun, H.J.; Huo, X.K.; Sun, P.Y.; Ma, X.D.; Peng, J.Y.; Liu, K.X. Effects of calycosin against high-fat diet-induced nonalcoholic fatty liver disease in mice. J. Gastroenterol. Hepatol. 2018, 33, 533–542. [Google Scholar] [CrossRef]
- Yu, J.; Zhu, L.J.; Zheng, H.H.; Gong, X.; Jiang, H.Y.; Chen, J.M.; Li, Y.H.; Zheng, H.M.; Qi, X.X.; Wang, Y.; et al. Sulfotransferases and Breast Cancer Resistance Protein Determine the Disposition of Calycosin in Vitro and in Vivo. Mol. Pharm. 2017, 14, 2917–2929. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Z.; Xu, F.; Dong, J.; Liang, J.; Hashi, Y.; Shang, M.Y.; Yang, D.H.; Wang, X.; Cai, S.Q. Profiling and identification of the metabolites of calycosin in rat hepatic 9000xg supernatant incubation system and the metabolites of calycosin-7-O-beta-D-glucoside in rat urine by HPLC-DAD-ESI-IT-TOF-MS(n) technique. J. Pharm. Biomed. Anal. 2012, 70, 425–439. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.Q.; Yan, R. Regioselective glucuronidation of the isoflavone calycosin by human liver microsomes and recombinant human UDP-glucuronosyltransferases. Chem. Biol. Interact. 2014, 220, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zheng, H.H.; Yu, J.; Zhu, L.J.; Yan, T.M.; Wu, P.; Lu, L.L.; Wang, Y.; Hu, M.; Liu, Z.Q. SGLT-1 Transport and Deglycosylation inside Intestinal Cells Are Key Steps in the Absorption and Disposition of Calycosin-7-O-beta-d-Glucoside in Rats. Drug Metab. Dispos. 2016, 44, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.T.; Chen, S.J.; Zhang, Y.Y.; Chen, L.Y.; Guo, X.Z.; Xu, Z.; Liu, H.; Hu, P.; Chen, Z.Y.; Li, Z.X.; et al. Erratum to "Absorption, liver first-pass effect, pharmacokinetics and tissue distribution of calycosin-7-O-ß-d-glucopyranoside (C7G) and its major active metabolite, calycosin, following oral administration of C7G in rats by LC-MS/MS". J.Pharm. Biomed. Anal. 2018, 148, 350–354. [Google Scholar] [CrossRef]
- Hou, Y.C.; Lin, S.P.; Tsai, S.Y.; Ko, M.H.; Chang, Y.C.; Chao, P.D. Flavonoid pharmacokinetics and tissue distribution after repeated dosing of the roots of Scutellaria baicalensis in rats. Planta. Med. 2011, 77, 455–460. [Google Scholar] [CrossRef]
- Tian, X.T.; Xu, Z.; Chen, M.C.; Hu, P.; Liu, F.; Sun, Z.L.; Liu, H.; Guo, X.Z.; Li, Z.X.; Huang, C.G. Simultaneous determination of eight bioactive compounds by LC-MS/MS and its application to the pharmacokinetics, liver first-pass effect, liver and brain distribution of orally administrated Gouteng-Baitouweng (G.B.) in rats. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2018, 1084, 122–131. [Google Scholar] [CrossRef]
- Liu, H.; Chen, M.C.; Yin, H.; Hu, P.; Wang, Y.Y.; Liu, F.; Tian, X.T.; Huang, C.G. Exploration of the hepatoprotective chemical base of an orally administered herbal formulation (YCHT) in normal and CCl4-intoxicated liver injury rats. Part 1: Metabolic profiles from the liver-centric perspective. J. Ethnopharmacol. 2019, 237, 81–91. [Google Scholar] [CrossRef]
- Tian, X.T.; Xu, Z.; Hu, P.; Yu, Y.Y.; Li, Z.X.; Ma, Y.; Chen, M.C.; Sun, Z.L.; Liu, F.; Li, J.Y.; et al. Determination of the antidiabetic chemical basis of Phellodendri Chinensis Cortex by integrating hepatic disposition in vivo and hepatic gluconeogenese inhibition in vitro. J. Ethnopharmacol. 2020, 263, 113215. [Google Scholar] [CrossRef]
- Wang, Q.; Zou, Z.Y.; Zhang, Y.Z.; Lin, P.; Lan, T.H.; Qin, Z.F.; Xu, D.P.; Wu, H.L.; Yao, Z.H. Characterization of chemical profile and quantification of major representative components of Wendan decoction, a classical traditional Chinese medicine formula. J. Sep. Sci. 2021, 44, 1036–1061. [Google Scholar] [CrossRef]
- Zhang, L.; Zuo, Z.; Lin, G. Intestinal and hepatic glucuronidation of flavonoids. Mol. Pharm. 2007, 4, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lin, G.; Zuo, Z. Position preference on glucuronidation of mono-hydroxylflavones in human intestine. Life Sci. 2006, 78, 2772–2780. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Bennett, G.; Jones, J.P. Mechanistic studies of uridine diphosphate glucuronosyltransferase. Chem. Biol. Interact. 1994, 90, 47–58. [Google Scholar] [CrossRef]
- Natsume, M.; Osakabe, N.; Oyama, M.; Sasaki, M.; Baba, S.; Nakamura, Y.; Osawa, T.; Terao, J. Structures of (-)-epicatechin glucuronide identified from plasma and urine after oral ingestion of (-)-epicatechin: Differences between human and rat. Free Radic. Biol. Med. 2003, 34, 840–849. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.L.; Gu, L.W. Flavonoid metabolism and challenges to understanding mechanisms of health effects. J. Sci. Food Agr. 2006, 86, 2487–2491. [Google Scholar] [CrossRef]
- Ruan, J.Q.; Li, S.; Li, Y.P.; Wu, W.J.; Lee, S.M.; Yan, R. The Presystemic Interplay between Gut Microbiota and Orally Administered Calycosin-7-O-beta-D-Glucoside. Drug Metab. Dispos. 2015, 43, 1601–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzales, G.B.; Smagghe, G.; Grootaert, C.; Zotti, M.; Raes, K.; Van Camp, J. Flavonoid interactions during digestion, absorption, distribution and metabolism: A sequential structure-activity/property relationship-based approach in the study of bioavailability and bioactivity. Drug Metab. Rev. 2015, 47, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Ikoma, Y.; Sugiura, M.; Yano, M.; Hasegawa, Y. Identification and quantification of the conjugated metabolites derived from orally administered hesperidin in rat plasma. J. Agric. Food Chem. 2004, 52, 6653–6659. [Google Scholar] [CrossRef]
- Rong, Y.; Tu, Y.F.; Yin, T.J.; Meng, Z.Y.; Dou, G.F.; Hu, M. Rapid intestinal glucuronidation and hepatic glucuronide recycling contributes significantly to the enterohepatic circulation of icaritin and its glucuronides in vivo. Arch. Toxicol. 2020, 94, 3737–3749. [Google Scholar] [CrossRef]
- Terao, J.; Murota, K.; Kawai, Y. Conjugated quercetin glucuronides as bioactive metabolites and precursors of aglycone in vivo. Food Funct. 2011, 2, 11–17. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO | tR (min) | Name | Formula (Neutral) | aP | L | S | Observed m/z | Calculated m/z | Fragment Ions | Diff (ppm) |
|---|---|---|---|---|---|---|---|---|---|---|
| M0 | 24.93 | Calycosin | C16H12O5 | + | + | + | 285.0754 | 285.0757 | 270.0519 (100%), 253.0493 (42%), 225.0543 (50%), 197.0591 (18%), 137.0231 (56%) | −1.05 |
| M1 | 8.55 | Calycosin-7-sulfate-3′-glucuronide | C22H2 O14S | + | − | + | 541.0643 | 541.0647 | 365.0324 (14%), 285.0752 (100%), 270.0522 (2%), 225.0539 (0.7%) | −0.74 |
| M2 * | 8.83 | 7,3′,4′-Trihydroxyisoflavone-disulfate | C15H10O11S2 | + | − | + | 430.9738 | 430.9737 | 351.0175 (13%), 271.0601 (100%), 137.0238 (5%) | 0.23 |
| M3 * | 9.30 | Daidzein-7-glucuronide | C21H18O10 | + | − | + | 431.0972 | 431.0973 | 255.0650 (100%), 199.0745 (13%), 137.0226 (22%) | −0.23 |
| M4 * | 10.28 | Calycosin-7,3′-disulfate | C16H12O11S2 | + | − | + | 444.9891 | 444.9894 | 365.0322 (9%), 285.0756 (100%), 270.0524 (4%), 253.0499 (2%), 137.0233 (1%) | −0.67 |
| M5 | 10.77 | Calycosin-7-glucuronide | C22H20O11 | + | − | + | 461.1077 | 461.1078 | 285.0752 (100%), 270.0516 (6%), 253.0491 (2%), 137.0228 (1%) | −0.22 |
| M6 * | 11.31 | 7,3′,4′-Trihydroxyisoflavone-glucuronide | C21H18O11 | + | − | + | 447.0920 | 447.0922 | 271.0594 (100%), 253.0492 (7%), 225.0538 (10%), 137.0236 (11%) | −0.45 |
| M7 * | 12.04 | 7,3′,4′-trihydroxyisoflavanone-glucuronide | C21H20O11 | + | + | + | 449.1076 | 449.1078 | 273.0752 (100%), 163.0388 (29%), 123.0448 (5%) | −0.45 |
| M8 | 13.79 | Calycosin-7-glucuronide-3′-sulfate | C22H20O14S | + | + | + | 541.0644 | 541.0647 | 365.0324 (46%), 285.0756 (100%), 270.0515 (2%) | −0.55 |
| M9 * | 14.38 | 7,3′,4′-Trihydroxyisoflavone-glucuronide | C21H18O11 | + | + | + | 447.0921 | 447.0922 | 271.0593 (100%), 253.0494 (4%), 225.0540 (4%), 137.0223 (4%) | −0.22 |
| M10 * | 14.63 | 7,3′,4′-Trihydroxyisoflavone-sulfate | C15H10O8S | + | + | + | 351.0161 | 351.0169 | 271.0590 (100%), 225.0542 (26%), 137.0237 (15%) | −2.28 |
| M11 * | 14.67 | 2,3-Dihydrocalycosin-glucuronide | C22H22O11 | + | − | + | 463.1224 | 463.1235 | 287.0914 (100%), 163.0386 (57%), 137.0588 (12%) | −2.38 |
| M12 | 16.07 | 7,3′,4′-Trihydroxyisoflavone | C15H10O5 | − | + | − | 271.0597 | 271.0601 | 253.0495 (34%), 225.0542 (100%), 197.0592 (64%), 137.0231 (93%) | −1.48 |
| M13 | 16.14 | Calycosin-3′-sulfate | C16H12O8S | + | + | + | 365.0323 | 365.0326 | 285.0755 (100%), 270.0518 (43%), 253.0488 (16%), 225.0542 (23%), 137.0228 (11%) | −0.82 |
| M14 | 16.76 | Calycosin-3′-glucuronide | C22H20O11 | + | + | + | 461.1069 | 461.1078 | 285.0753 (100%), 270.0515 (6%), 253.0489 (2%), 137.0226 (1%) | −1.95 |
| M15* | 21.91 | Daidzein | C15H10O4 | + | + | + | 255.0646 | 255.0652 | 227.0696 (47%), 199.0748 (100%), 181.0641 (64%), 137.0230 (94%) | −2.35 |
| M16* | 22.19 | 7,3′-dihydroxy-4′-methoxyisoflavanone | C16H14O5 | + | + | + | 287.0905 | 287.0914 | 163.0386 (100%), 137.0589 (79%) | −3.13 |
| M17* | 22.59 | Calycosin-7-sulfate-3′-methyl ether | C17H14O8S | − | + | + | 379.0481 | 379.0482 | 299.0907 (100%), 271.0606 (14%), 253.0495 (3%), 225.0553 (4%) | −0.26 |
| M18* | 23.08 | Calycosin-7-glucuronide-3′-methyl ether | C23H22O11 | + | + | + | 475.1230 | 475.1235 | 299.0914 (100%), 271.0592 (7%), 253.0488 (3%), 225.0548 (2%) | −1.05 |
| M19* | 23.71 | 7,3′,4′-Trihydroxyisoflavone-3′-methyl ether | C16H12O5 | + | + | + | 285.0755 | 285.0757 | 270.0511 (100%), 253.0487 (45%), 225.0539 (54%), 137.0229 (62%) | −0.70 |
| M20* | 27.20 | Calycosin-3′-methyl ether | C17H14O5 | + | + | − | 299.0914 | 299.0914 | 271.0604 (100%), 253.0488 (38%), 137.0236 (89%) | 0.00 |
| M21 | 27.74 | Formononetin | C16H12O4 | + | − | − | 269.0805 | 269.0808 | 253.0487 (41%), 225.0541 (40%), 197.0589 (100%), 137.0229 (19%), 118.0409 (35%) | −1.11 |
| PK Parameters | T1/2 (h) | Tmax (h) | Cmax ± SEM (μmol/L) | AUC0-t ± SEM (h μmol/L) | AUC (Calycosin Glucuronides) / AUC (Calycosin) | |
|---|---|---|---|---|---|---|
| Portal vein plasma | Calycosin | 1.4 | 1.0 | 12.6 ± 0.8 | 40.0 ± 1.9 | 3.5 |
| Calycosin glucuronides | 13.0 | 1.0 | 33.9±1.1 | 138.3 ± 3.4 | ||
| Systemic plasma | Calycosin | 1.9 | 1.0 | 0.06 ± 0.01 | 0.15 ± 0.02 | 934.7 |
| Calycosin glucuronides | 14.2 | 2.0 | 44.2 ± 6.3 | 140.2 ± 11.3 | ||
| Liver | Calycosin | 0.8 | 1.0 | 31.1 ± 1.3 | 119.6 ± 6.2 | 0.2 |
| Calycosin glucuronides | 0.7 | 2.0 | 8.5 ± 1.0 | 21.2 ± 1.7 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, H.; Liu, H.; Hu, P.; Chen, S.; Ye, Y.; Huang, C.; Tian, X. Intestinal Glucuronidation, Prior to Hepatic Glucuronidation, Plays an Important Role in the Low Circulating Levels of Calycosin. Separations 2022, 9, 115. https://doi.org/10.3390/separations9050115
Jiang H, Liu H, Hu P, Chen S, Ye Y, Huang C, Tian X. Intestinal Glucuronidation, Prior to Hepatic Glucuronidation, Plays an Important Role in the Low Circulating Levels of Calycosin. Separations. 2022; 9(5):115. https://doi.org/10.3390/separations9050115
Chicago/Turabian StyleJiang, Haodong, Huan Liu, Pei Hu, Shuoji Chen, Yaqing Ye, Chenggang Huang, and Xiaoting Tian. 2022. "Intestinal Glucuronidation, Prior to Hepatic Glucuronidation, Plays an Important Role in the Low Circulating Levels of Calycosin" Separations 9, no. 5: 115. https://doi.org/10.3390/separations9050115
APA StyleJiang, H., Liu, H., Hu, P., Chen, S., Ye, Y., Huang, C., & Tian, X. (2022). Intestinal Glucuronidation, Prior to Hepatic Glucuronidation, Plays an Important Role in the Low Circulating Levels of Calycosin. Separations, 9(5), 115. https://doi.org/10.3390/separations9050115