
Identification of Phlogacantholide C as a Novel ADAM10 Enhancer from Traditional Chinese Medicinal Plants




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Cell Culture
2.3. Cytotoxicity Test
2.4. Transfection and Promoter Assays
2.5. Western Blotting
3. Results and Discussions
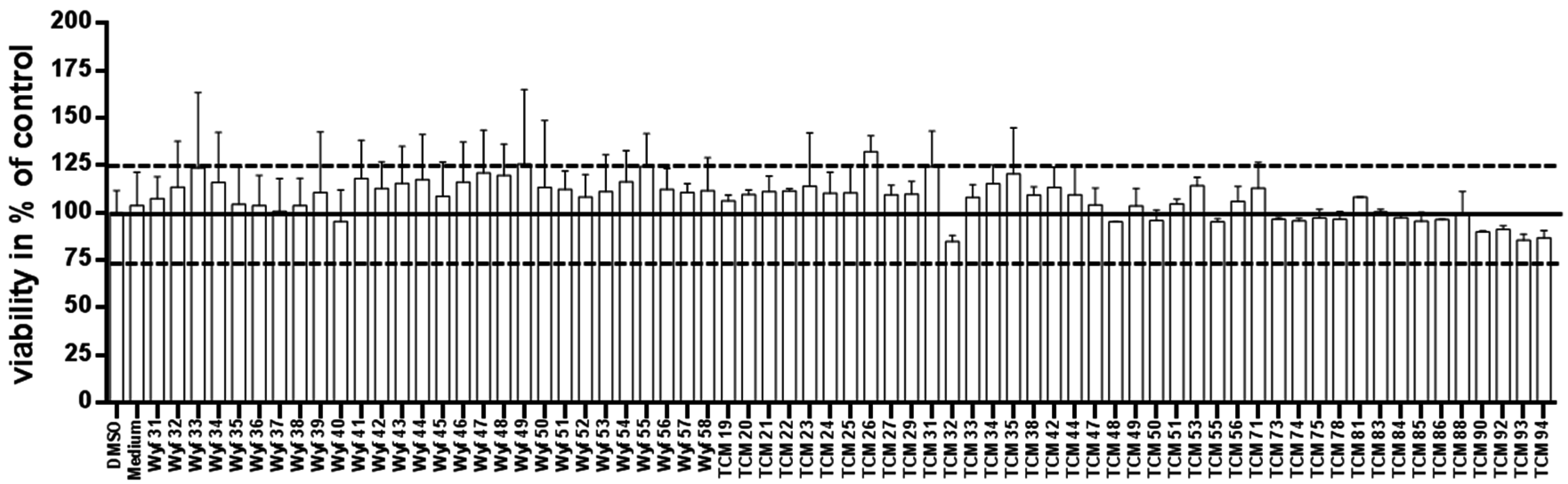
3.1. Results for Toxicity Assay
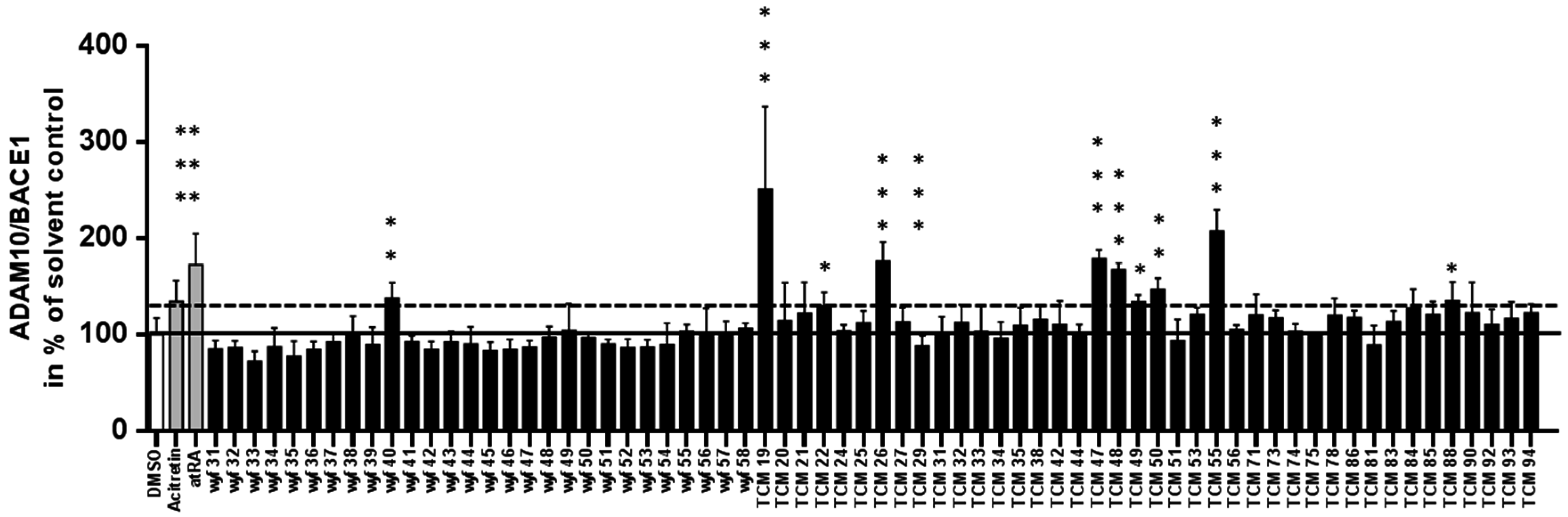
3.2. Results for Dual Promoter Assay
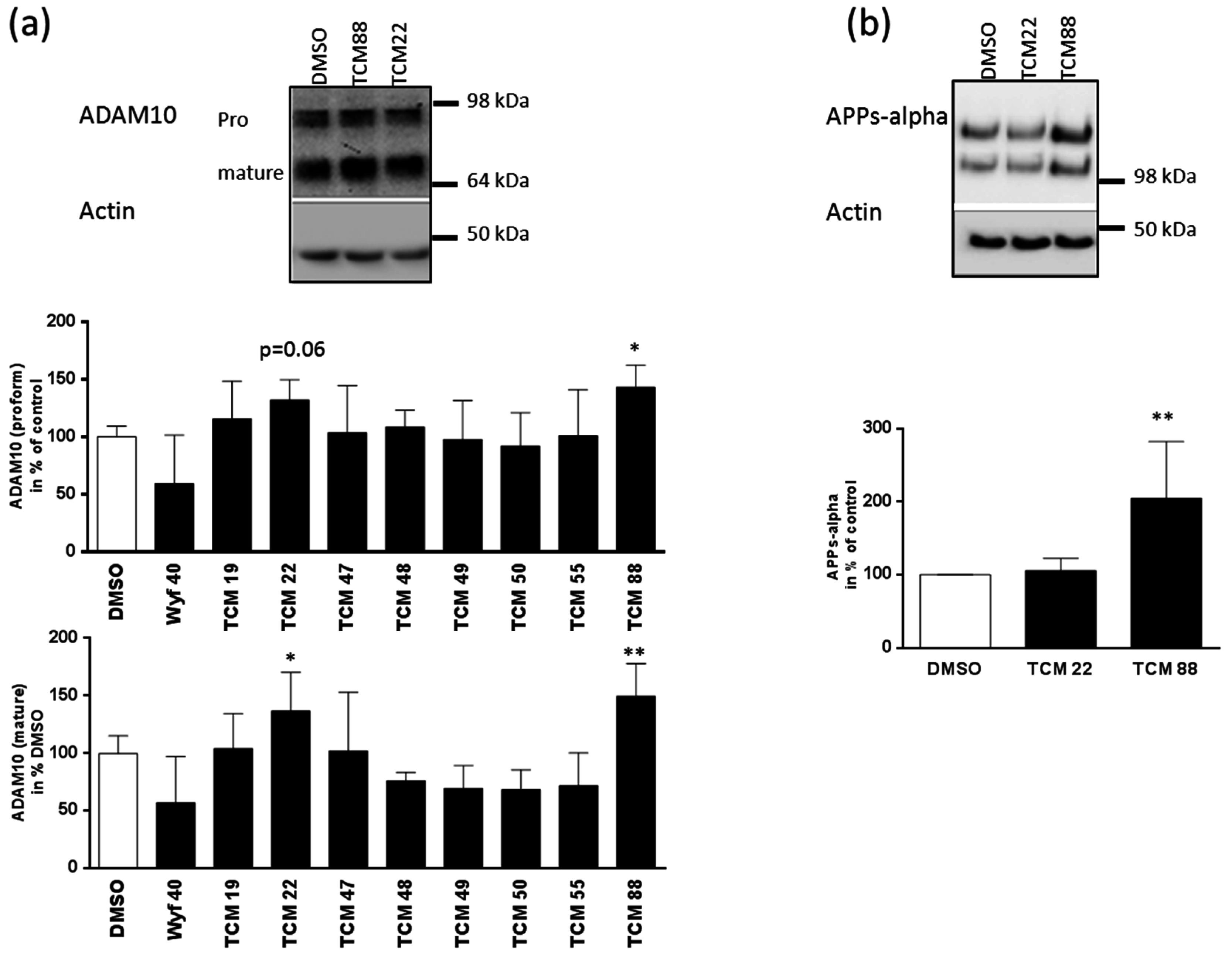
3.3. Results for ADAM10 Expression and Enzymatic Activity
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chung, J.A.; Cummings, J.L. Neurobehavioral and neuropsychiatric symptoms in Alzheimer’s disease: Characteristics and treatment. Neurol. Clin. 2000, 18, 829–846. [Google Scholar] [CrossRef]
- Wimo, A.; Jonsson, L.; Bond, J.; Prince, M.; Winblad, B. Alzheimer Disease International. The worldwide economic impact of dementia 2010. Alzheimers Dement. 2013, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tanzi, R.E. The genetics of Alzheimer disease. Cold Spring Harb. Perspect. Med. 2012, 2, a006296. [Google Scholar] [CrossRef] [PubMed]
- Citron, M.; Diehl, T.S.; Gordon, G.; Biere, A.L.; Seubert, P.; Selkoe, D.J. Evidence that the 42- and 40-amino acid forms of amyloid beta protein are generated from the beta-amyloid precursor protein by different protease activities. Proc. Natl. Acad. Sci. USA 1996, 93, 13170–13175. [Google Scholar] [CrossRef] [PubMed]
- Vassar, R.; Bennett, B.D.; Babu-Khan, S.; Kahn, S.; Mendiaz, E.A.; Denis, P.; Teplow, D.B.; Ross, S.; Amarante, P.; Loeloff, R.; et al. Beta-secretase cleavage of Alzheimer’s amyloid precursor protein by the transmembrane aspartic protease BACE. Science 1999, 286, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Zhang, J.; Ostaszewski, B.L.; Kimberly, W.T.; Seubert, P.; Koo, E.H.; Shen, J.; Selkoe, D.J. Presenilin 1 regulates the processing of beta-amyloid precursor protein C-terminal fragments and the generation of amyloid beta-protein in endoplasmic reticulum and Golgi. Biochemistry 1998, 37, 16465–16471. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Liu, F.; Gong, C.X. Alzheimer disease therapeutics: Focus on the disease and not just plaques and tangles. Biochem. Pharmacol. 2014, 88, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Lammich, S.; Kojro, E.; Postina, R.; Gilbert, S.; Pfeiffer, R.; Jasionowski, M.; Haass, C.; Fahrenholz, F. Constitutive and regulated alpha-secretase cleavage of Alzheimer’s amyloid precursor protein by a disintegrin metalloprotease. Proc. Natl. Acad. Sci. USA 1999, 96, 3922–3927. [Google Scholar] [CrossRef] [PubMed]
- Fol, R.; Braudeau, J.; Ludewig, S.; Abel, T.; Weyer, S.W.; Roederer, J.P.; Brod, F.; Audrain, M.; Bemelmans, A.-P.; Buchholz, C.J.; et al. Viral gene transfer of APPsalpha rescues synaptic failure in an Alzheimer’s disease mouse model. Acta Neuropathol. 2016, 131, 247–266. [Google Scholar] [CrossRef] [PubMed]
- Gakhar-Koppole, N.; Hundeshagen, P.; Mandl, C.; Weyer, S.W.; Allinquant, B.; Muller, U.; Ciccolini, F. Activity requires soluble amyloid precursor protein alpha to promote neurite outgrowth in neural stem cell-derived neurons via activation of the MAPK pathway. Eur. J. Neurosci. 2008, 28, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Hasebe, N.; Fujita, Y.; Ueno, M.; Yoshimura, K.; Fujino, Y.; Yamashita, T. Soluble beta-amyloid Precursor Protein Alpha binds to p75 neurotrophin receptor to promote neurite outgrowth. PLoS ONE 2013, 8, e82321. [Google Scholar] [CrossRef] [PubMed]
- Hick, M.; Herrmann, U.; Weyer, S.W.; Mallm, J.P.; Tschape, J.A.; Borgers, M.; Mercken, M.; Roth, F.C.; Draguhn, A.; Slomianka, L.; et al. Acute function of secreted amyloid precursor protein fragment APPsalpha in synaptic plasticity. Acta Neuropathol. 2015, 129, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Howes, M.J.; Houghton, P.J. Ethnobotanical treatment strategies against Alzheimer’s disease. Curr. Alzheimer Res. 2012, 9, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. WHO Traditional Medicine Strategy 2002–2005; WHO: Geneva, Switzerland, 2002. [Google Scholar]
- Park, H.L.; Lee, H.S.; Shin, B.C.; Liu, J.P.; Shang, Q.; Yamashita, H.; Lim, B. Traditional medicine in China, Korea, and Japan: A brief introduction and comparison. Evid. Based Complement. Alternat. Med. 2012, 2012, 429103. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; More, S.V.; Han, S.D.; Choi, J.Y.; Choi, D.K. Promising therapeutics with natural bioactive compounds for improving learning and memory—A review of randomized trials. Molecules 2012, 17, 10503–10539. [Google Scholar] [CrossRef] [PubMed]
- Gray, N.E.; Morre, J.; Kelley, J.; Maier, C.S.; Stevens, J.F.; Quinn, J.F.; Soumyanath, A. Caffeoylquinic acids in Centella asiatica protect against amyloid-beta toxicity. J. Alzheimers Dis. 2014, 40, 359–373. [Google Scholar] [PubMed]
- Miyamae, Y.; Kurisu, M.; Murakami, K.; Han, J.; Isoda, H.; Irie, K.; Shigemori, H. Protective effects of caffeoylquinic acids on the aggregation and neurotoxicity of the 42-residue amyloid beta-protein. Bioorg. Med. Chem. 2012, 20, 5844–5849. [Google Scholar] [CrossRef] [PubMed]
- Schuck, F.; Schmitt, U.; Reinhardt, S.; Freese, C.; Lee, I.S.; Thines, E.; Efferth, T.; Endres, K. Extract of Caragana sinica as a potential therapeutic option for increasing alpha-secretase gene expression. Phytomedicine 2015, 22, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.H.; Li, B.G.; Zhang, X.Y.; Qi, H.Y.; Zhou, M.; Zhang, G.L. Two diterpenes and three diterpene glucosides from Phlogacanthus curviflorus. J. Nat. Prod. 2005, 68, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Chui, H.B. Flora Yunnanica; Science Press: Beijing, China, 2006; Volume 1. [Google Scholar]
- Phurailatpam, A.K.; Singh, S.R.; Chanu, T.M.; Ngangbam, P. Phlogacanthus—An important medicinal plant of North East India: A review. Afr. J. Agric. Res. 2014, 9, 2068–2072. [Google Scholar]
- Panyaphu, K.; On, T.V.; Sirisa-ard, P.; Srisa-nga, P.; Chansa Kaow, S.; Nathakarnkitkul, S. Medicinal plants of the Mien (Yao) in Northern Thailand and their potential value in the primary healthcare of postpartum women. J. Ethnopharmacol. 2011, 135, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Kahl, S.; Paulus, K.; Adams, M.; Rauh, R.; Boechzelt, H.; Hao, X.; Kaina, B.; Bauer, R. Phytochemistry and pharmacogenomics of natural products derived from traditional Chinese medicine and Chinese materia medica with activity against tumor cells. Mol. Cancer Ther. 2008, 7, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Mahringer, A.; Karamustafa, S.; Klotz, D.; Kahl, S.; Konkimalla, V.B.; Wang, Y.; Wang, J.; Liu, H.Y.; Boechzelt, H.; Hao, X.; et al. Inhibition of P-glycoprotein at the blood-brain barrier by phytochemicals derived from traditional Chinese medicine. Cancer Genom. Proteom. 2010, 7, 191–205. [Google Scholar]
- Reinhardt, S.; Schuck, F.; Grosgen, S.; Riemenschneider, M.; Hartmann, T.; Postina, R.; Grimm, M.; Endres, K. Unfolded protein response signaling by transcription factor XBP-1 regulates ADAM10 and is affected in Alzheimer’s disease. FASEB J. 2014, 28, 978–997. [Google Scholar] [CrossRef] [PubMed]
- Endres, K.; Postina, R.; Schroeder, A.; Mueller, U.; Fahrenholz, F. Shedding of the amyloid precursor protein-like protein APLP2 by disintegrin-metalloproteinases. FEBS J. 2005, 272, 5808–5820. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Di, Y.; Yang, X.; Li, S.; Wang, Y.; Hao, X. Hydroquinone diglycoside acyl esters from the stems of Glycosmis pentaphylla. Phytochemistry 2006, 67, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.F.; Lai, G.F.; Efferth, T.; Cao, J.X.; Luo, S.D. New glycosides from Tetracentron sinense and their cytotoxic activity. Chem. Biodivers. 2006, 3, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Chen, J.; Lin, H. Metabolism of flavonoids via enteric recycling: Mechanistic studies of disposition of apigenin in the Caco-2 cell culture model. J. Pharmacol. Exp. Ther. 2003, 307, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, H.; Iinuma, M. Reduction of membrane fluidity by antibacterial sophoraflavanone G isolated from Sophora exigua. Phytomedicine 2000, 7, 161–165. [Google Scholar] [CrossRef]
- Son, J.K.; Park, J.S.; Kim, J.A.; Kim, Y.; Chung, S.R.; Lee, S.H. Prenylated flavonoids from the roots of Sophora flavescens with tyrosinase inhibitory activity. Planta Med. 2003, 69, 559–561. [Google Scholar] [PubMed]
- Prinzen, C.; Muller, U.; Endres, K.; Fahrenholz, F.; Postina, R. Genomic structure and functional characterization of the human ADAM10 promoter. FASEB J. 2005, 19, 1522–1524. [Google Scholar] [CrossRef] [PubMed]
- Tippmann, F.; Hundt, J.; Schneider, A.; Endres, K.; Fahrenholz, F. Up-regulation of the alpha-secretase ADAM10 by retinoic acid receptors and acitretin. FASEB J. 2009, 23, 1643–1654. [Google Scholar] [CrossRef] [PubMed]
- Head, E.; Lott, I.T.; Wilcock, D.M.; Lemere, C.A. Aging in Down Syndrome and the Development of Alzheimer’s Disease Neuropathology. Curr. Alzheimer Res. 2016, 13, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Lai, G.-F.; Wang, X.-Y.; Wang, Y.-F.; Cao, J.-X.; Luo, S.-D.; Peng, J. Diterpenes and Diterpene Glucosides from Phlogacanthus curviflorus. Helvetica Chim. Acta 2009, 92, 470–480. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Substance Code | Substance | Formula | Plant | Concentration (mg/mL) | Effect on APP Promoter (MW ± SD) |
|---|---|---|---|---|---|
| Wyf40 | Cynaropicrin | C19H22O6 | Saussurea deltoidea | 2 | 93.13 ± 24.96 |
| TCM19 | Sophoraflavon G | C25H28O6 | Sophora flavescens, Sophora pachycarpa, and Sophora exigua | 2 | 115.3 ± 58.45 |
| TCM22 | Norkurarinol | C25H30O7 | Sophora flavescens | 2 | 84.50 ± 47.86 |
| TCM47 | 5-Methoxy-3-methyl-9H-carbazol-2-ol | C14H13NO2 | Glycosmis pentaphylla | 12 | 86.88 ± 36.46 |
| TCM48 | 7-methoxyglycomaurin | C18H17NO2 | Glycosmis rupestris | 12 | 61.38 ± 25.81 |
| TCM49 | glybomine B | C19H21NO2 | Glycosmis arborea | 12 | 68.57 ± 34.91 |
| TCM50 | (2E)-2-Methyl-4-[7-(3-methyl-2-buten-1-yl)-1H-indol-3-yl]-2-buten-1-ol | C18H23NO | 12 | 117.8 ± 53.42 | |
| TCM55 | (4R,4aS,8aR,10R,10aR,12S,13S,14bS)-4-methyl-12-((methylthio)methyl)decahydro-1H,8aH,10H,11H-4,14b,10-(epiethane[1,1,2]triyl)-10a,13-ethanoisochromeno[4,3-g]oxazolo[3,2-a]azocin-11-one | C23H33NO3S | Spiraea japonica | 4 | 96.75 ± 53.34 |
| TCM88 | Phlogacantholide C | C20H28O4 | Pholgacanthus curviflorus | 2 | 80.25 ± 31.15 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meineck, M.; Schuck, F.; Abdelfatah, S.; Efferth, T.; Endres, K. Identification of Phlogacantholide C as a Novel ADAM10 Enhancer from Traditional Chinese Medicinal Plants. Medicines 2016, 3, 30. https://doi.org/10.3390/medicines3040030
Meineck M, Schuck F, Abdelfatah S, Efferth T, Endres K. Identification of Phlogacantholide C as a Novel ADAM10 Enhancer from Traditional Chinese Medicinal Plants. Medicines. 2016; 3(4):30. https://doi.org/10.3390/medicines3040030
Chicago/Turabian StyleMeineck, Myriam, Florian Schuck, Sara Abdelfatah, Thomas Efferth, and Kristina Endres. 2016. "Identification of Phlogacantholide C as a Novel ADAM10 Enhancer from Traditional Chinese Medicinal Plants" Medicines 3, no. 4: 30. https://doi.org/10.3390/medicines3040030
APA StyleMeineck, M., Schuck, F., Abdelfatah, S., Efferth, T., & Endres, K. (2016). Identification of Phlogacantholide C as a Novel ADAM10 Enhancer from Traditional Chinese Medicinal Plants. Medicines, 3(4), 30. https://doi.org/10.3390/medicines3040030