
Stable Schooling Formations Emerge from the Combined Effect of the Active Control and Passive Self-Organization



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
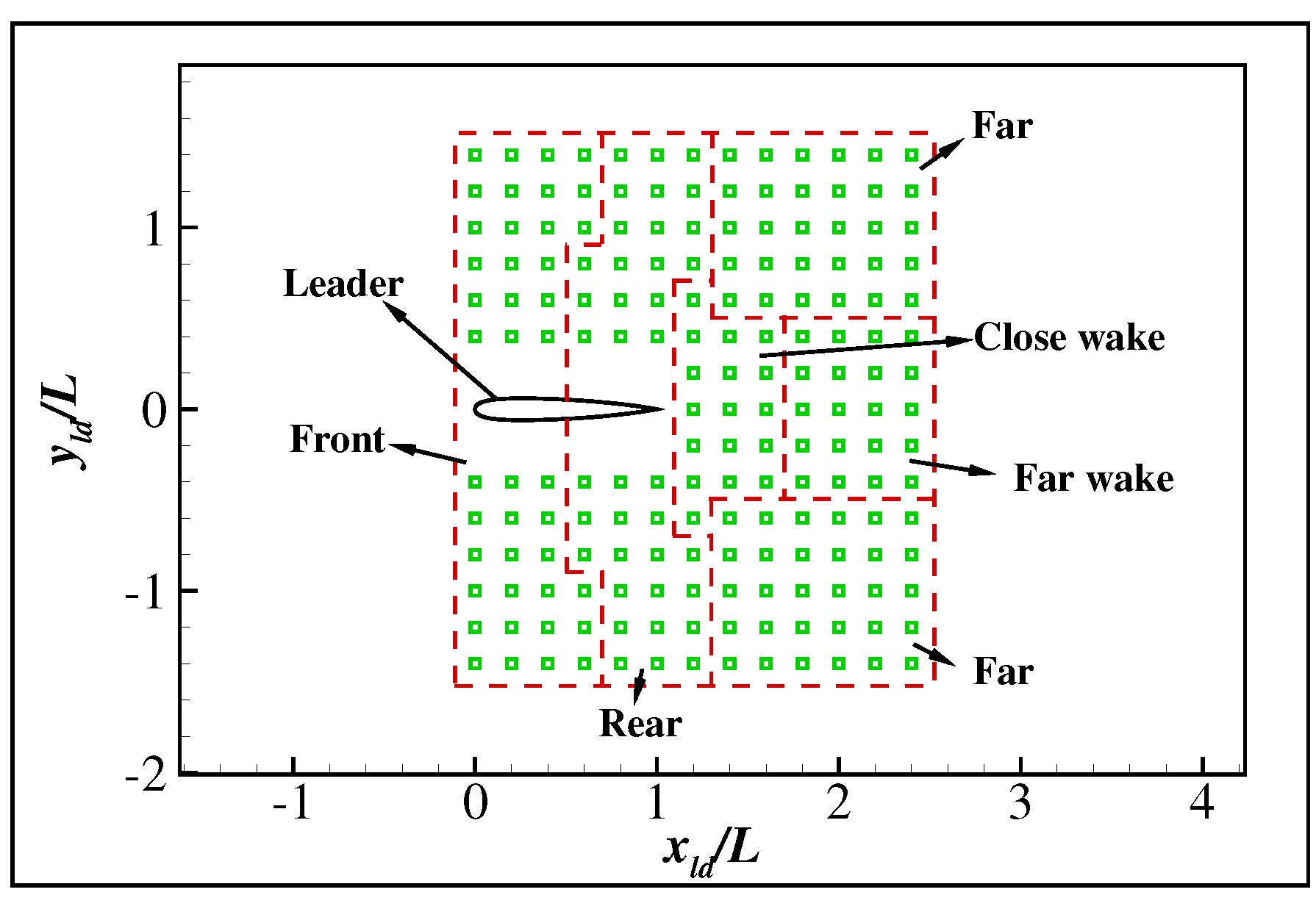{kind=link}
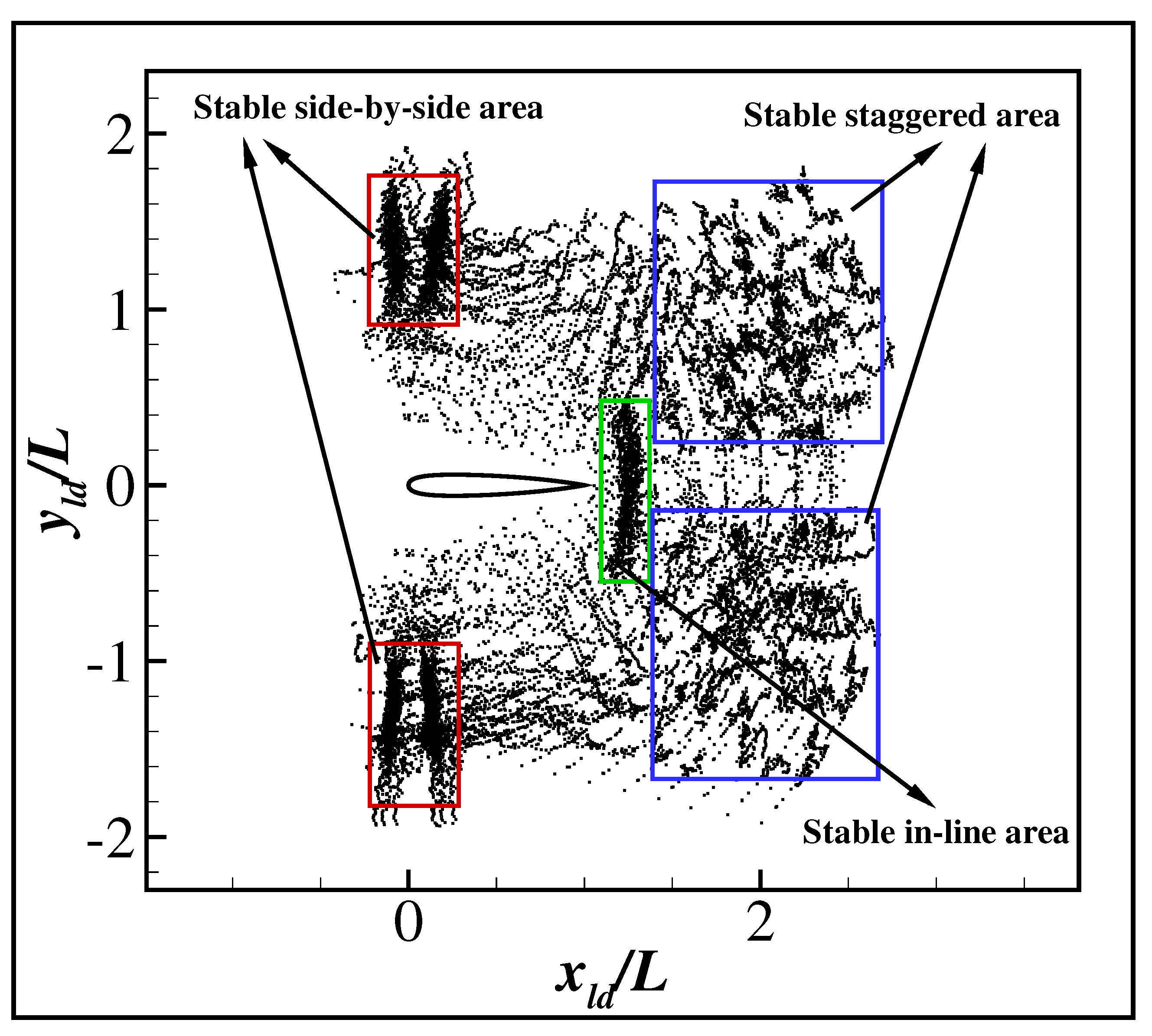{kind=link}
{kind=link}
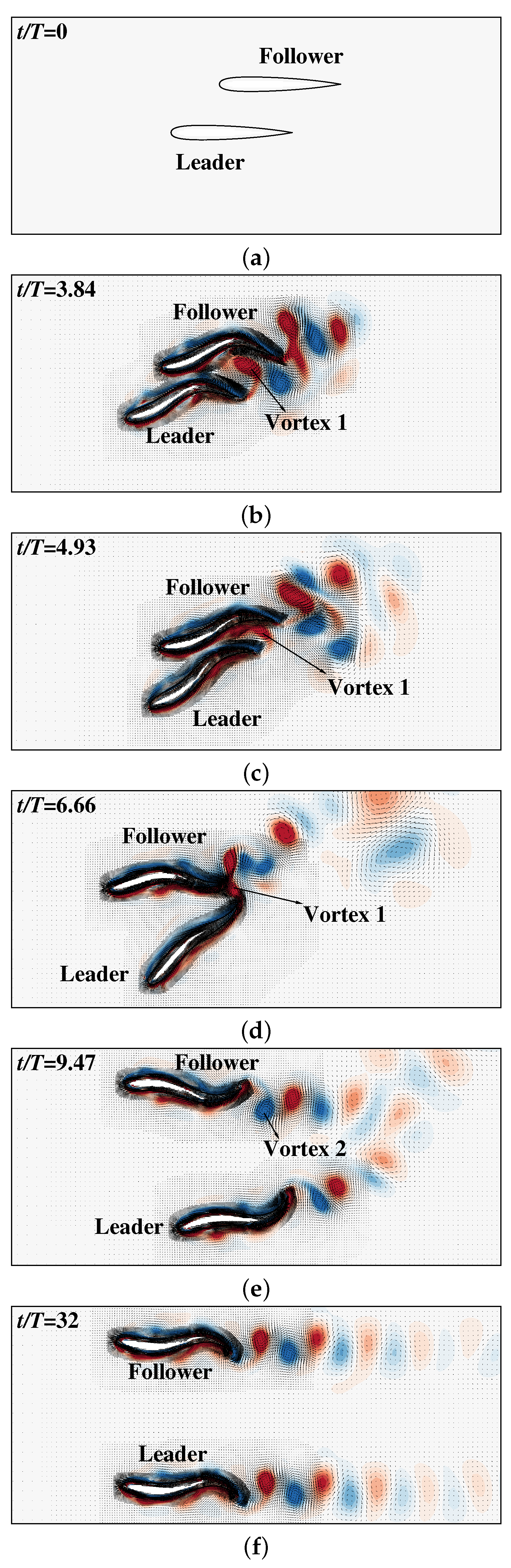{kind=link}
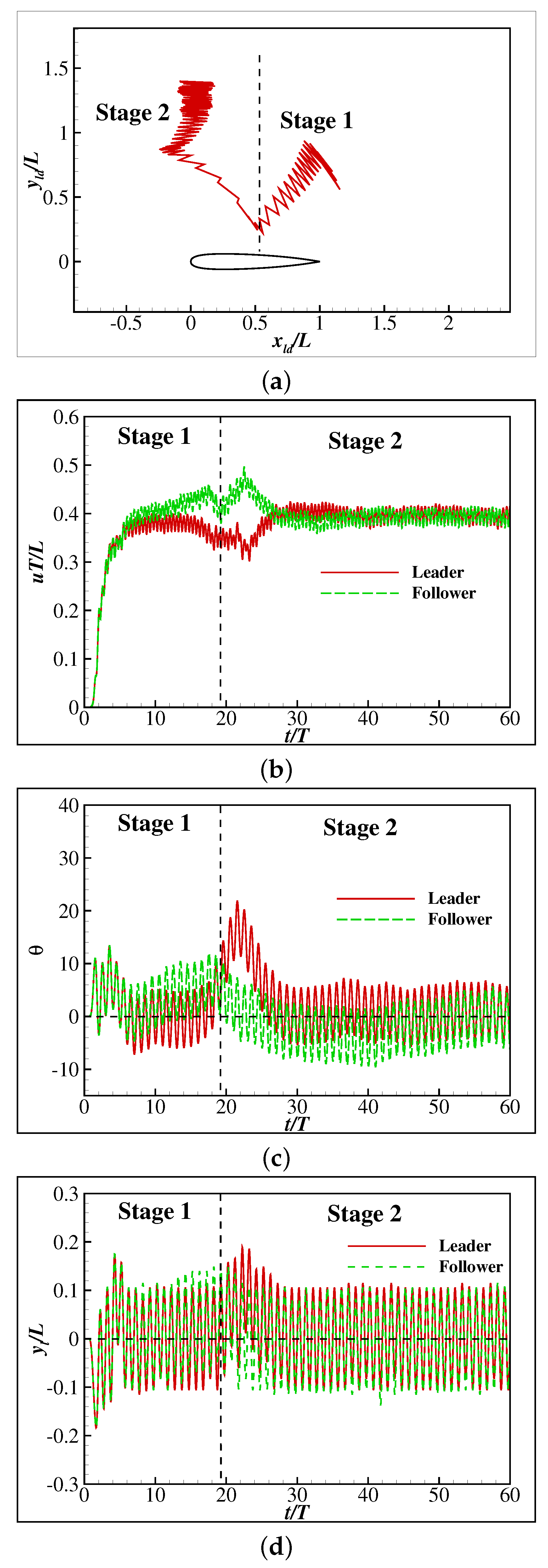{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methodology
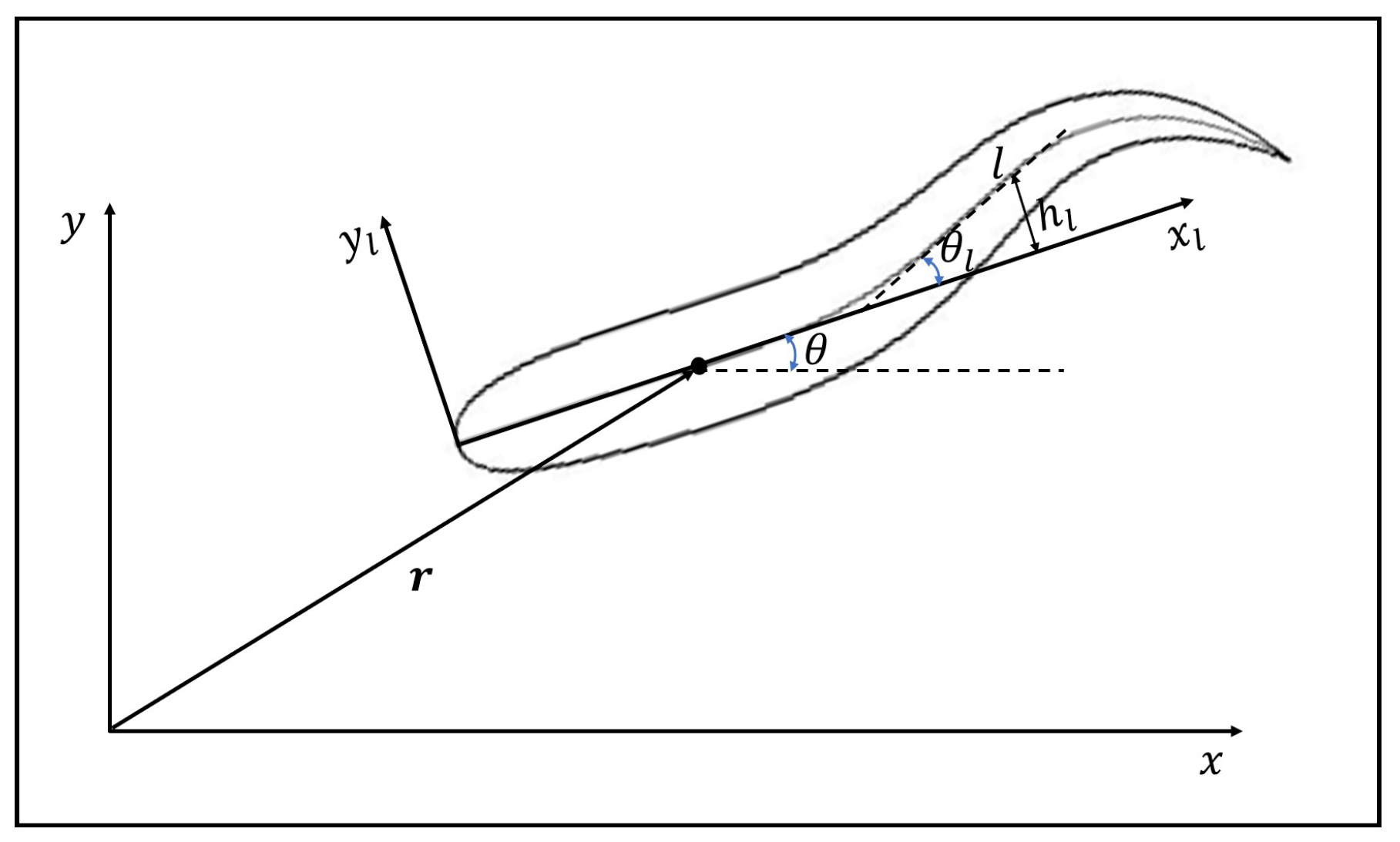
2.1. Kinematic Model of the Fish
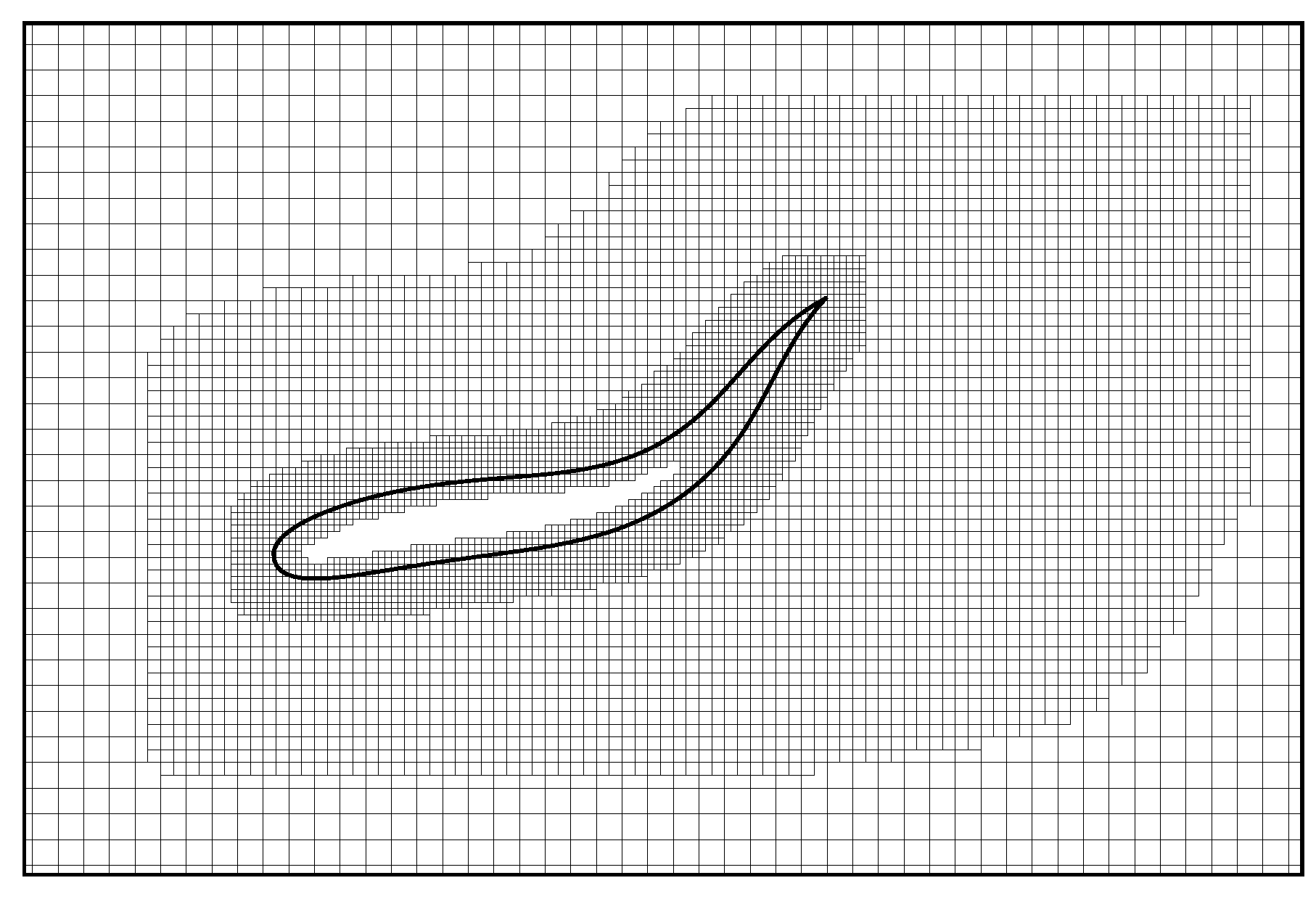
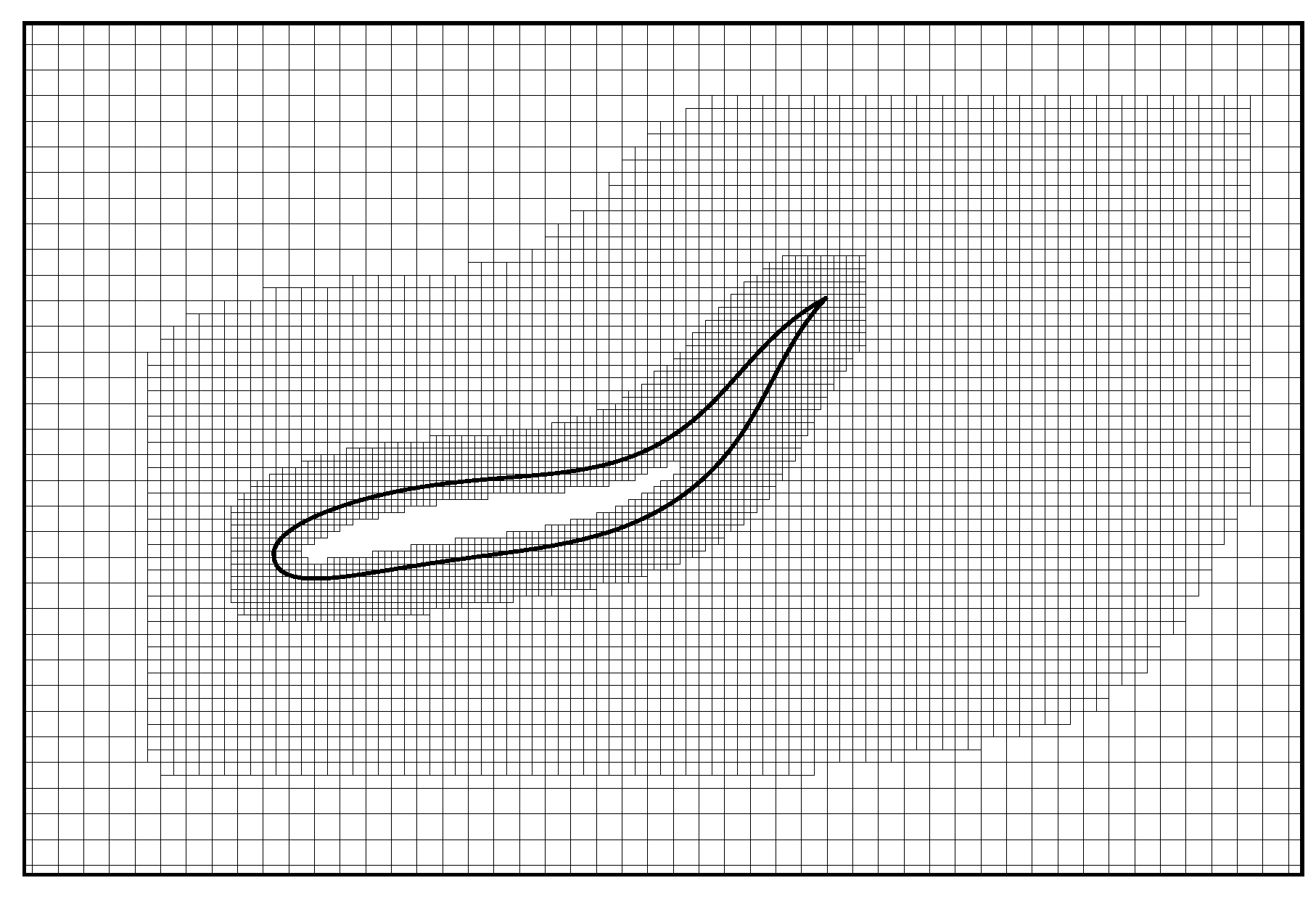
2.2. Immersed Boundary-Lattice Boltzmann Method
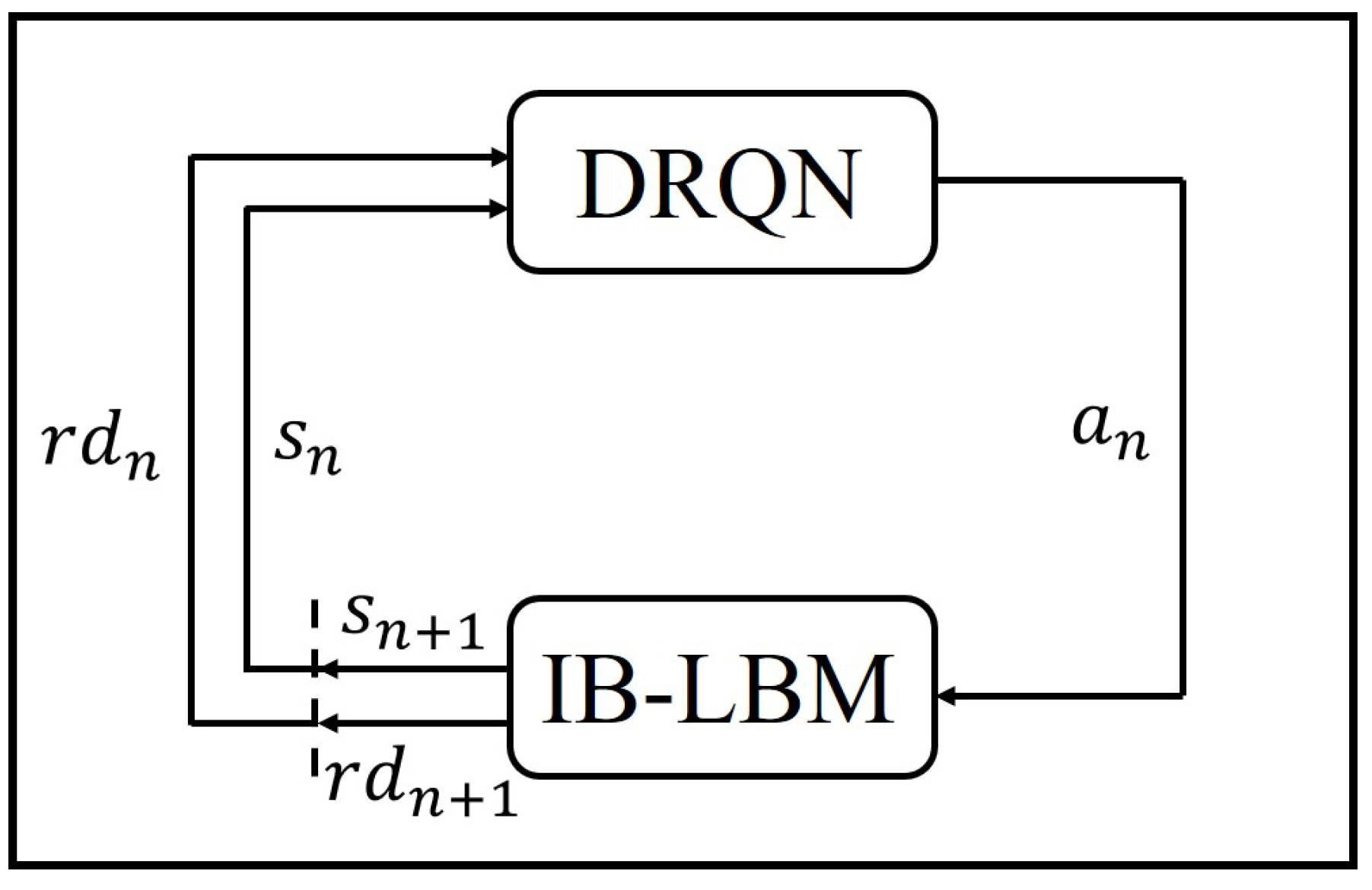
2.3. Deep Reinforcement Learning
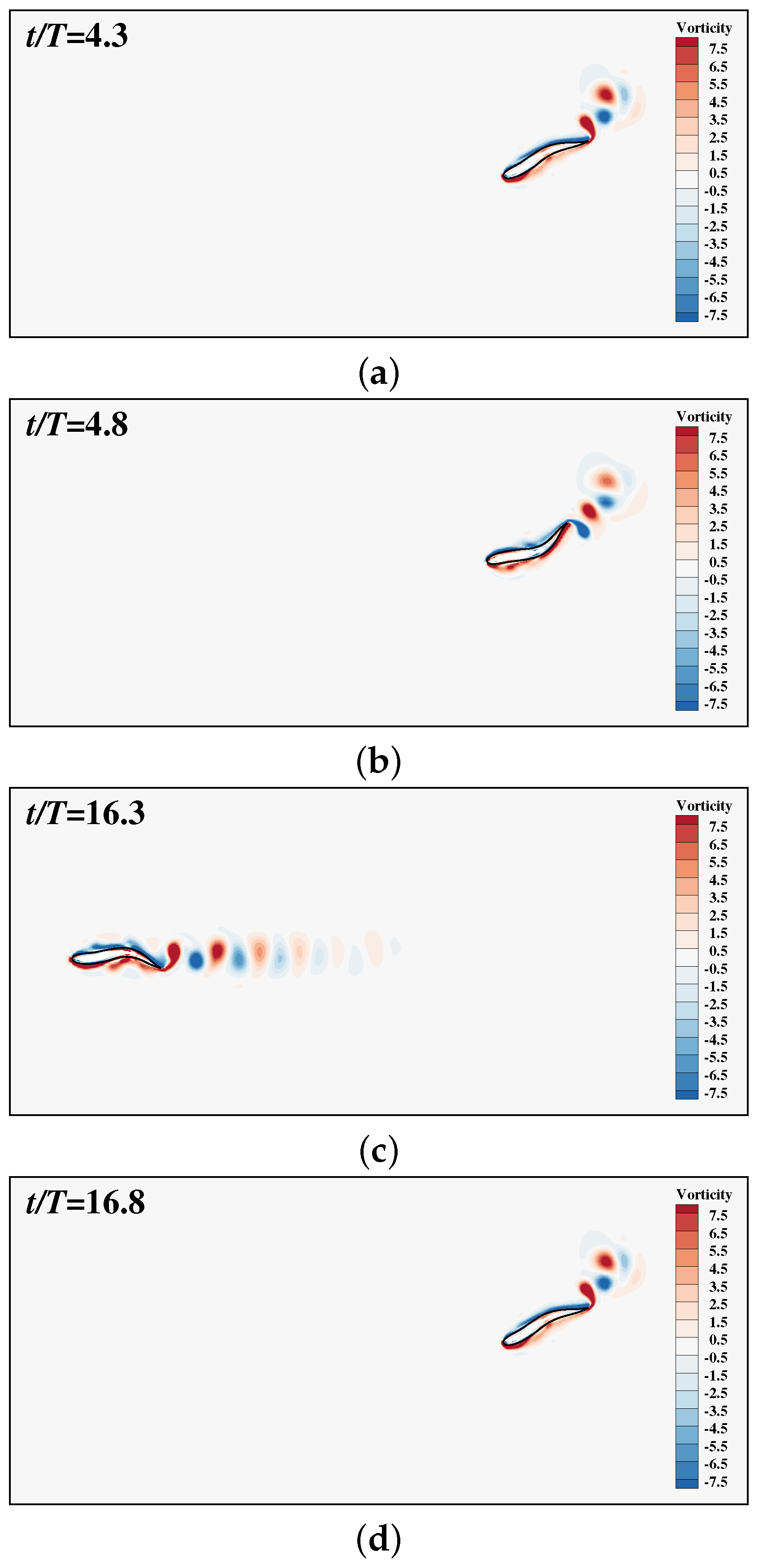
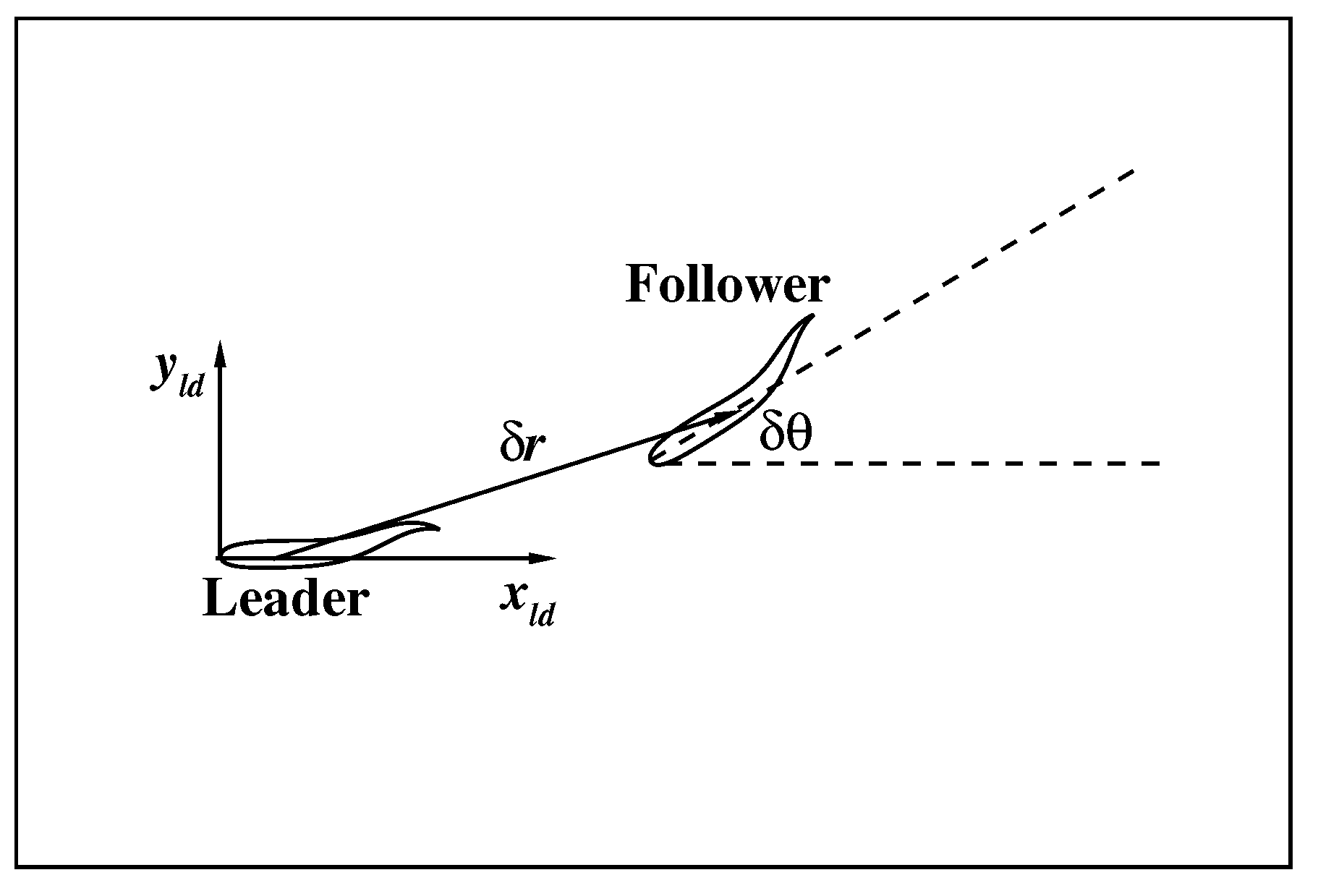
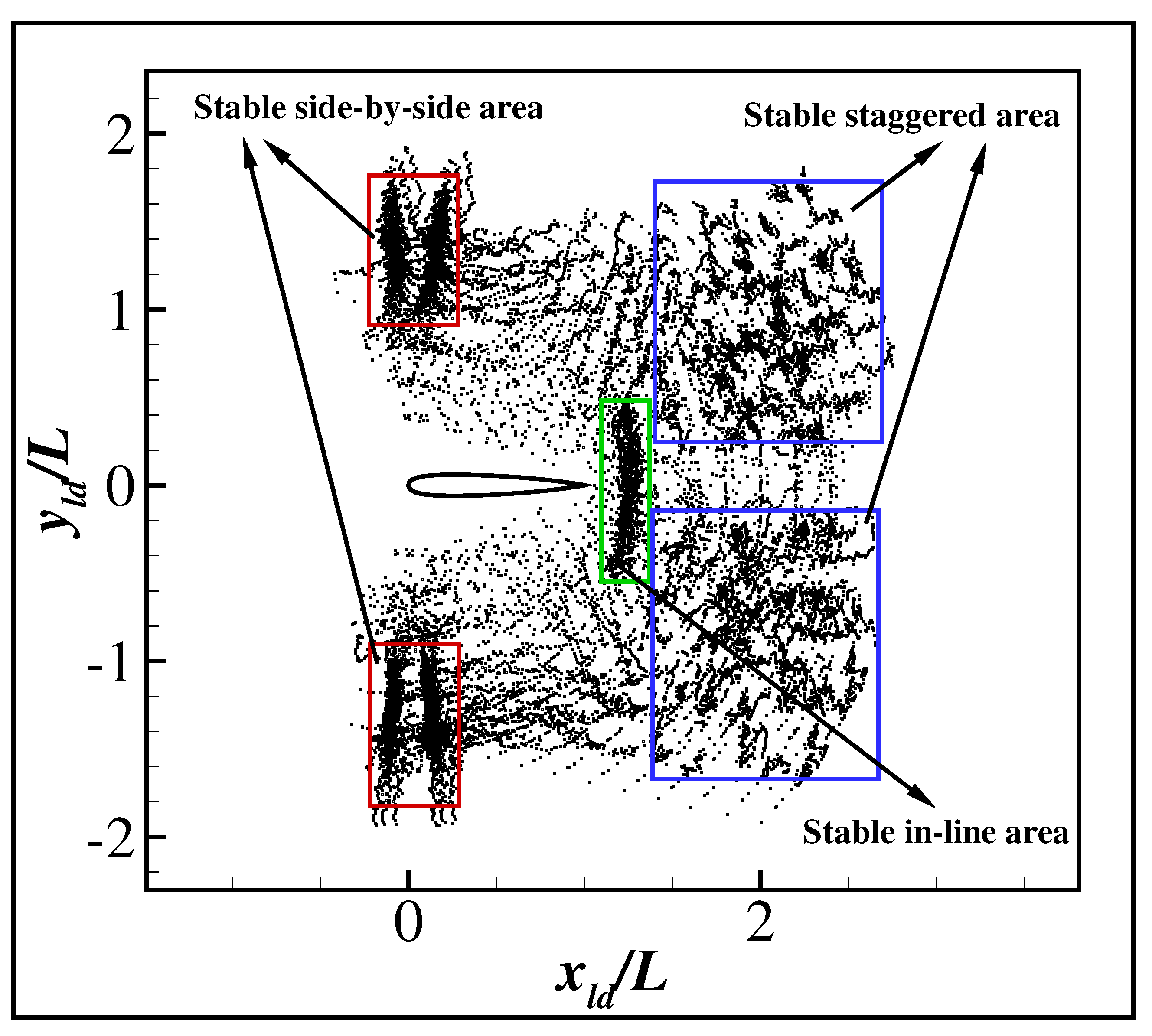
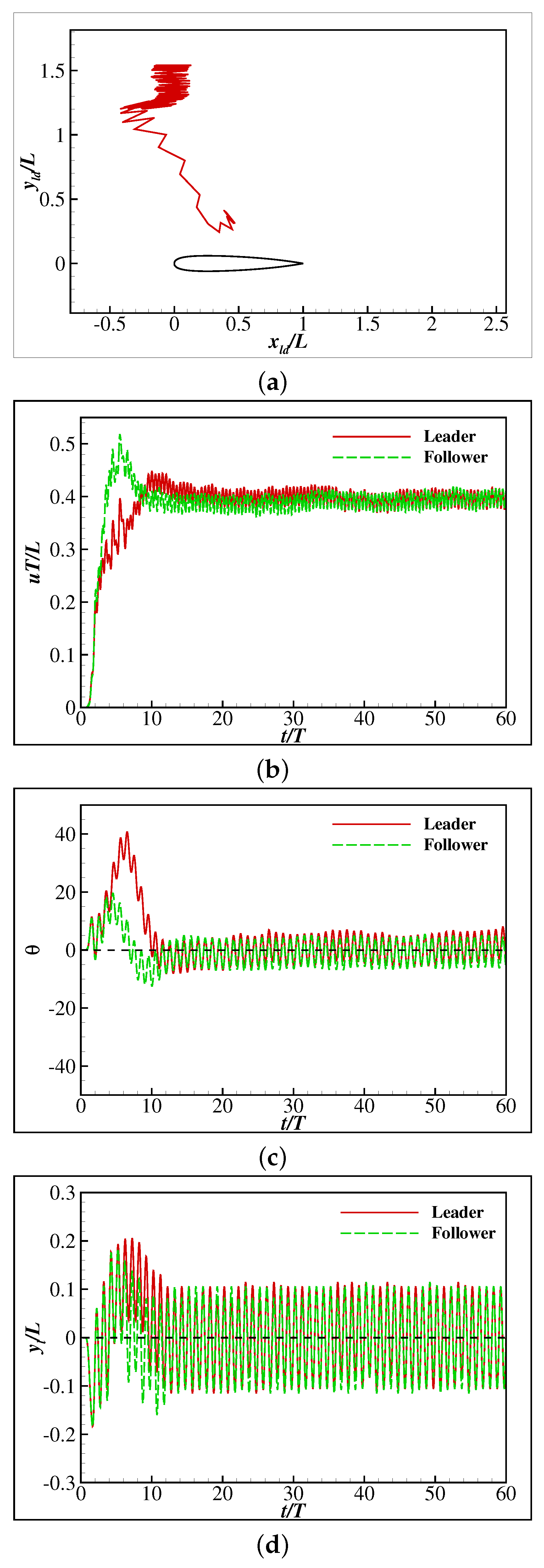
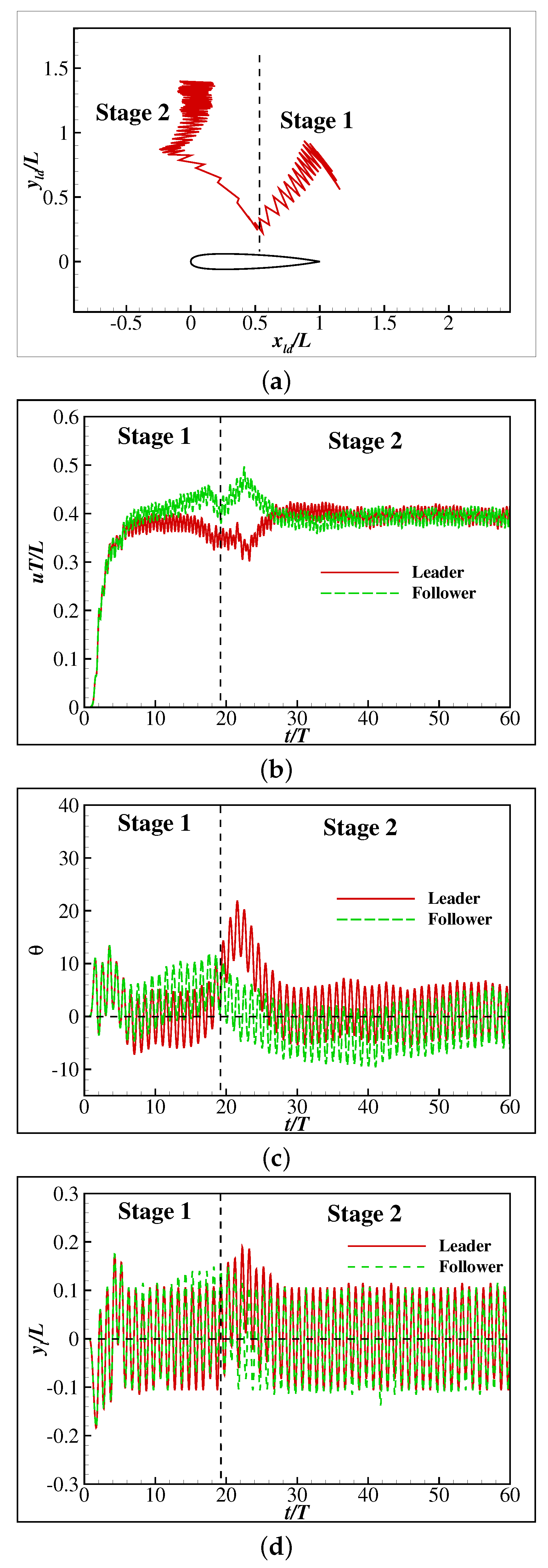
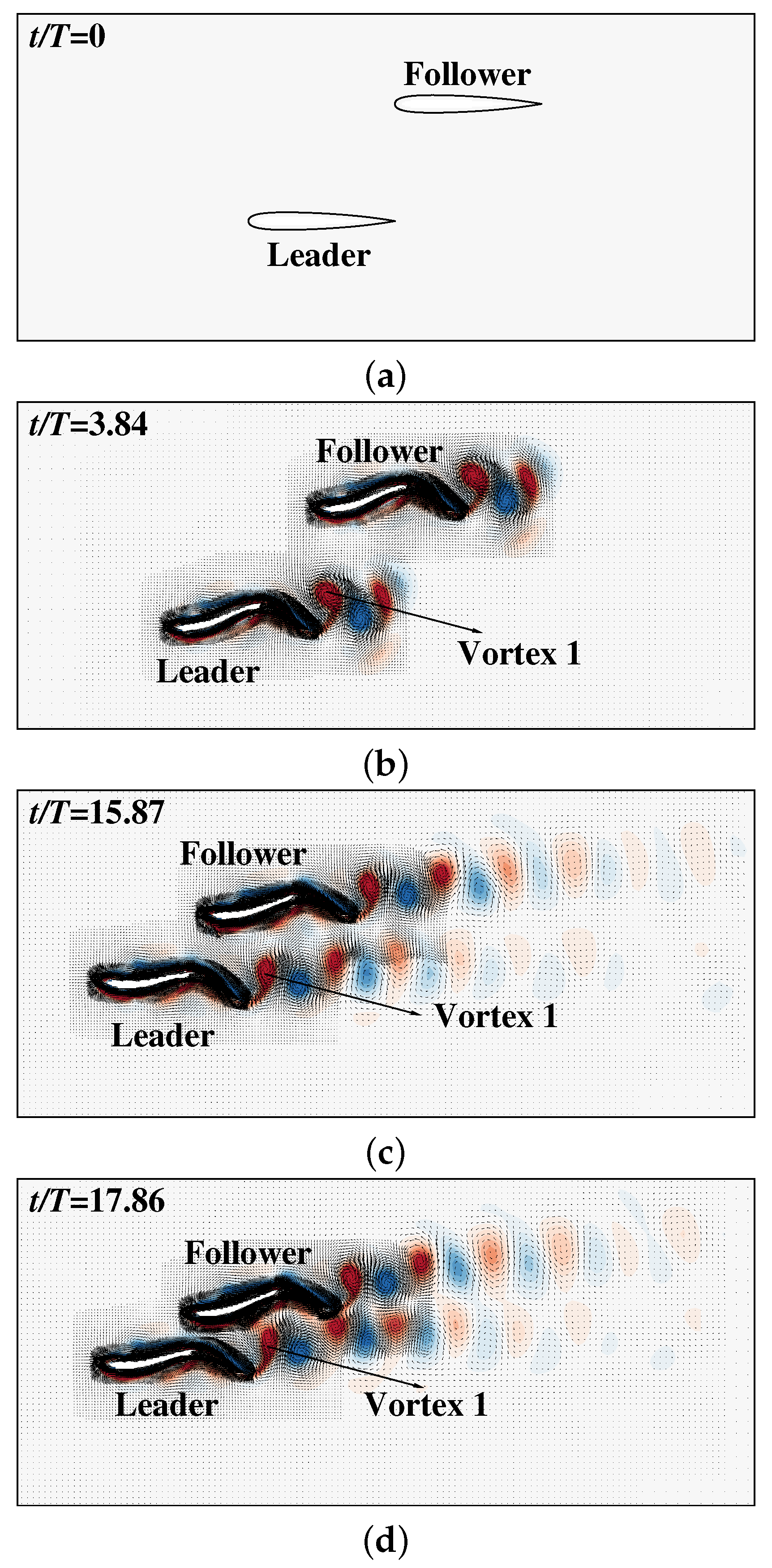
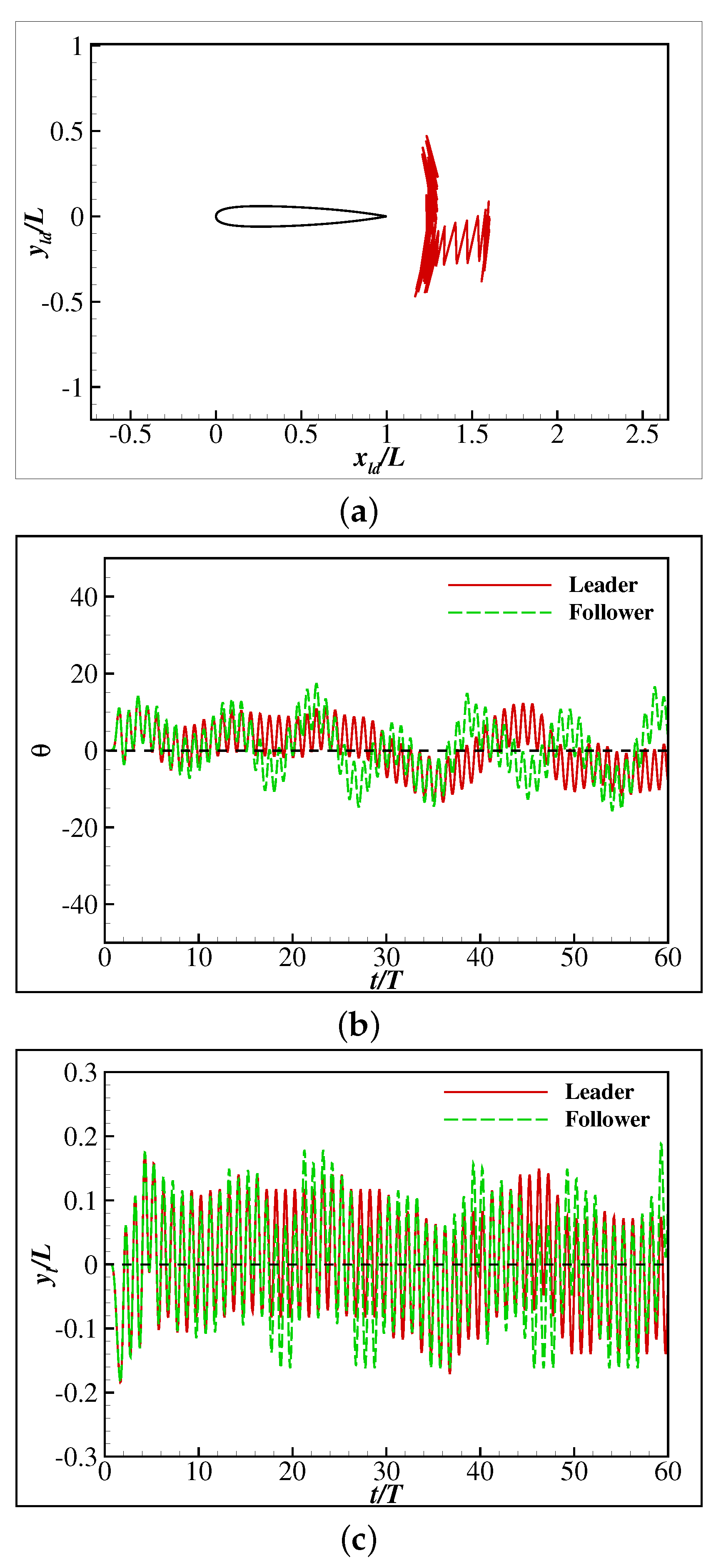
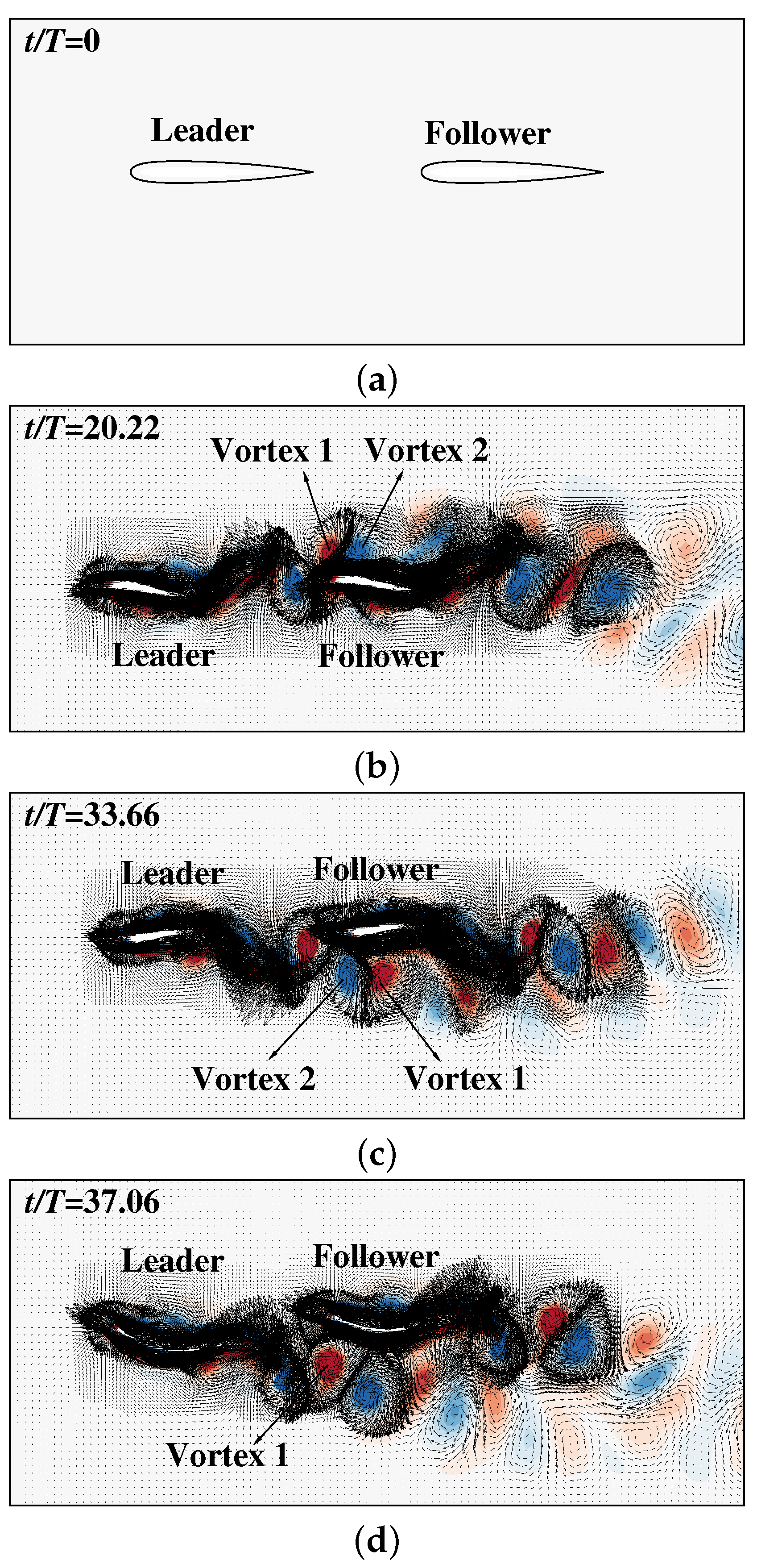
3. Results and Discussion
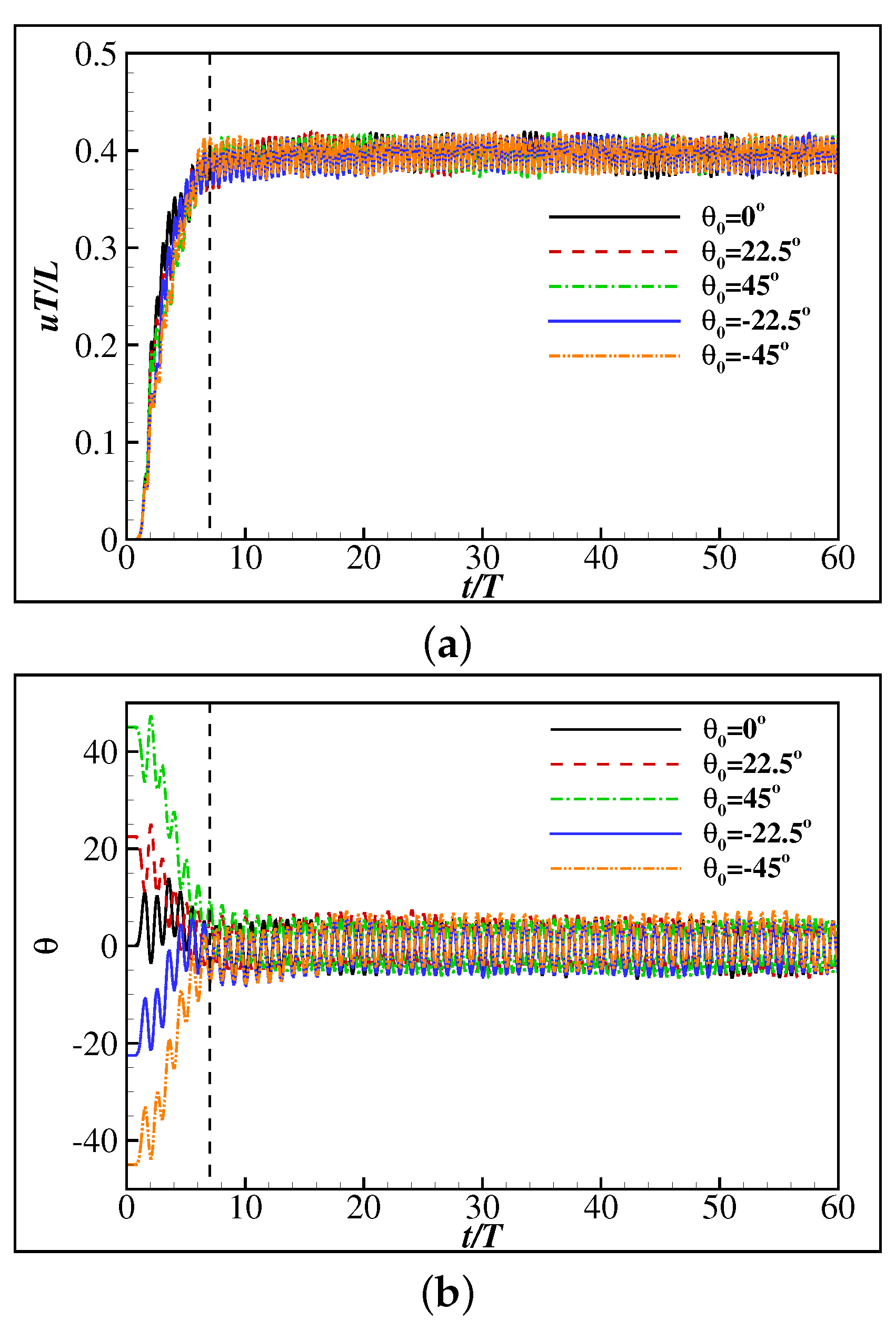
3.1. Learning to Maintain a Given Speed and Orientation
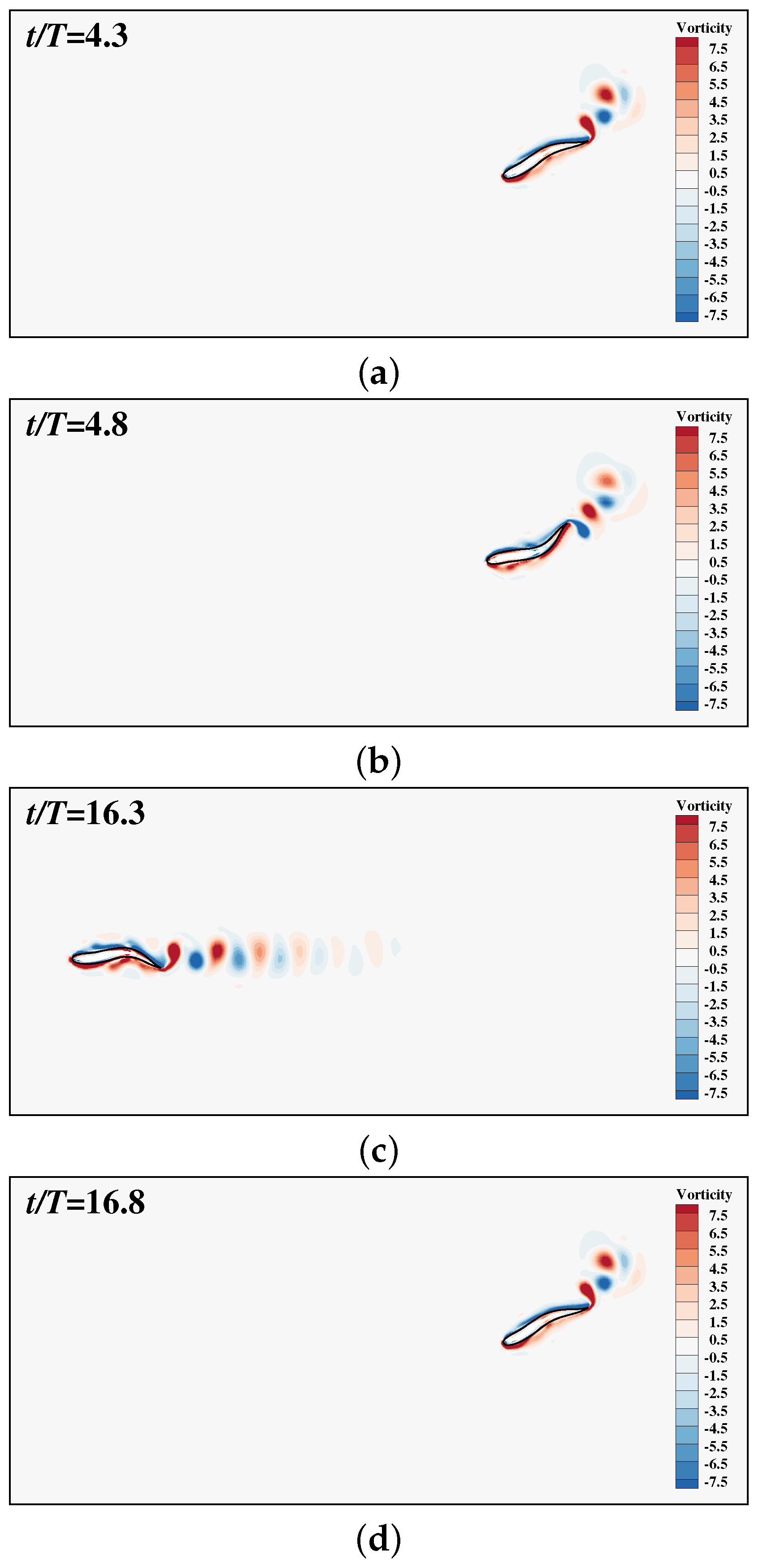
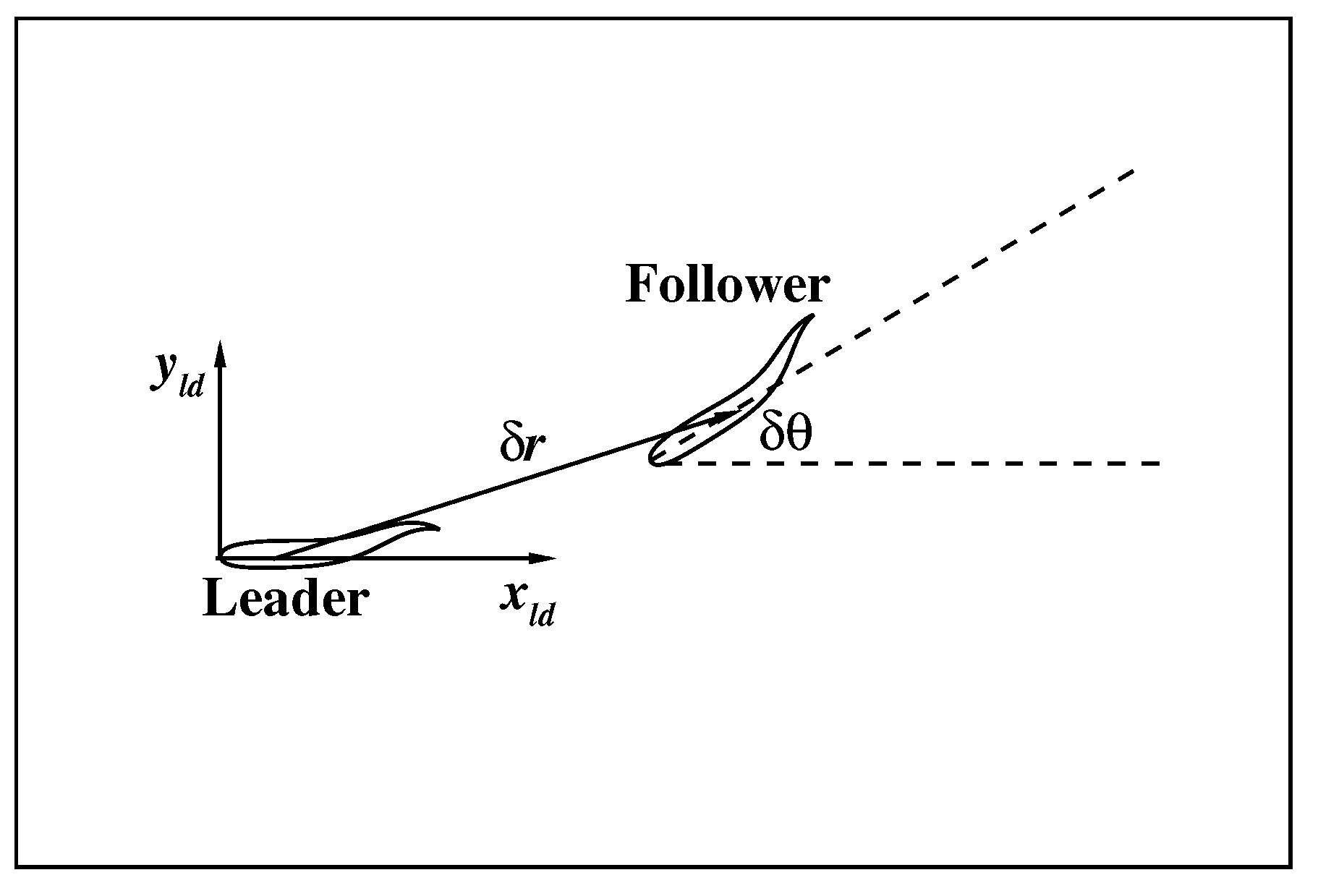
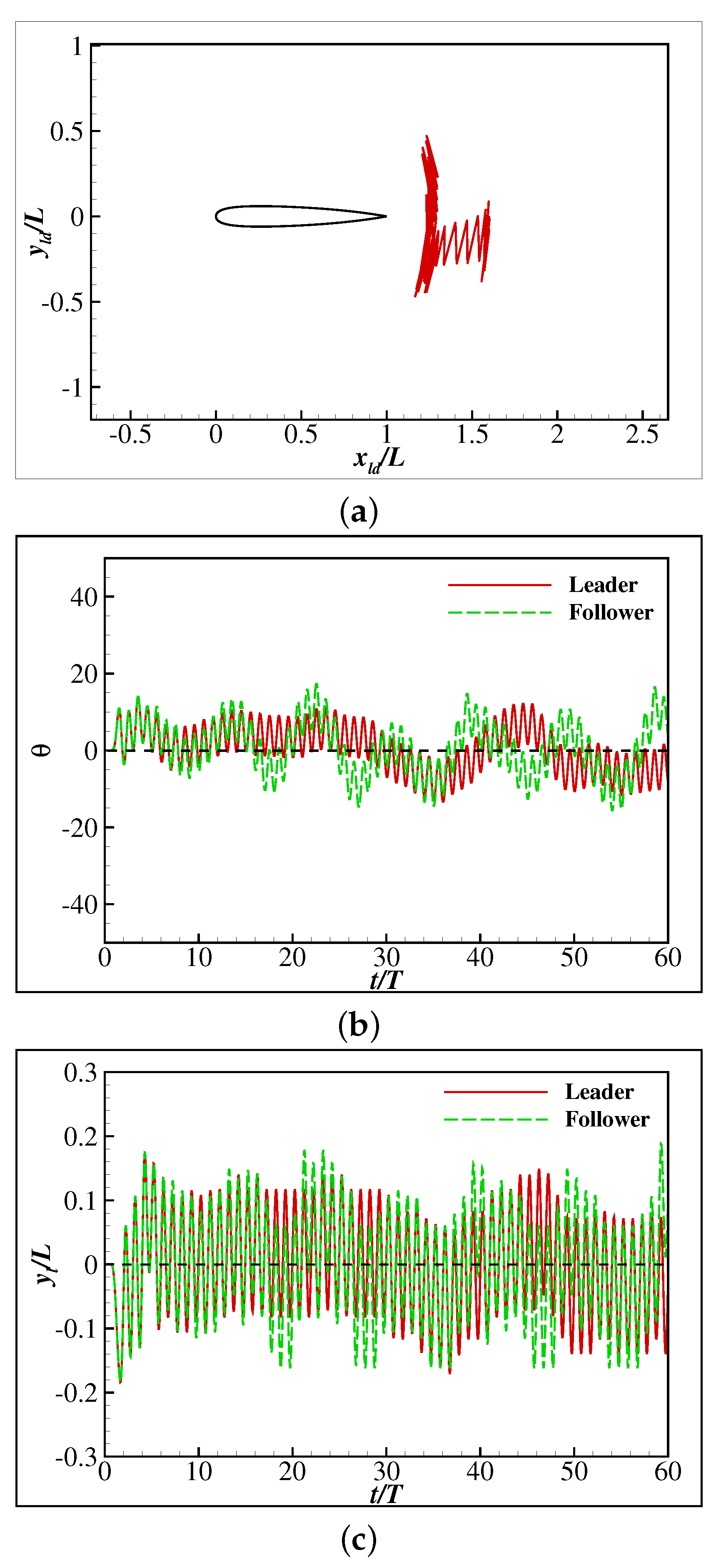
3.2. The Collective Motion of Two Smart Swimmers
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DoF | degree of freedom |
| DRL | deep reinforcement learning |
| IB-LBM | immersed boundary-lattice Boltzmann method |
| FSI | fluid–structure interaction |
| IBM | immersed boundary method |
| LBM | lattice Boltzmann method |
| DRQN | deep recurrent Q-network |
| LSTM-RNN | long-short-term-memory recurrent neural network |
References
- Larsson, M. Why do fish school? Curr. Zool. 2012, 58, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.E.; Godin, J.G.J. Anti-predator responses to conspecific and heterospecific skin extracts by threespine sticklebacks: Alarm pheromones revisited. Behaviour 1997, 134, 1123–1134. [Google Scholar] [CrossRef]
- Pitcher, T.; Magurran, A.; Winfield, I. Fish in larger shoals find food faster. Behav. Ecol. Sociobiol. 1982, 10, 149–151. [Google Scholar] [CrossRef]
- Pitcher, T.J. Functions of shoaling behaviour in teleosts. In The Behaviour of Teleost Fishes; Springer: Berlin/Heidelberg, Germany, 1986; pp. 294–337. [Google Scholar]
- Weihs, D. Hydromechanics of fish schooling. Nature 1973, 241, 290–291. [Google Scholar] [CrossRef]
- Lighthill, S.J. Mathematical Biofluiddynamics; Society for Industrial and Applied Mathematics SIAM: Philadelphia, PA, USA, 1975. [Google Scholar]
- Deng, J.; Shao, X.M.; Yu, Z.S. Hydrodynamic studies on two traveling wavy foils in tandem arrangement. Phys. Fluids 2007, 19, 113104. [Google Scholar] [CrossRef]
- Boschitsch, B.M.; Dewey, P.A.; Smits, A.J. Propulsive performance of unsteady tandem hydrofoils in an in-line configuration. Phys. Fluids 2014, 26, 051901. [Google Scholar] [CrossRef]
- Ashraf, I.; Godoy-Diana, R.; Halloy, J.; Collignon, B.; Thiria, B. Synchronization and collective swimming patterns in fish (Hemigrammus bleheri). J. R. Soc. Interface 2016, 13, 20160734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, F.B.; Wang, W.; Wu, J.; Sui, Y. Swimming performance and vorticity structures of a mother–calf pair of fish. Comput. Fluids 2016, 124, 1–11. [Google Scholar] [CrossRef]
- Kurt, M.; Moored, K. Unsteady Performance of Finite-Span Pitching Propulsors in Side-by-Side Arrangements. In Proceedings of the 2018 Fluid Dynamics Conference, Atlanta, GA, USA, 24 June 2018; p. 3732. [Google Scholar]
- Ramananarivo, S.; Fang, F.; Oza, A.; Zhang, J.; Ristroph, L. Flow interactions lead to orderly formations of flapping wings in forward flight. Phys. Rev. Fluids 2016, 1, 071201. [Google Scholar] [CrossRef]
- Zhu, X.; He, G.; Zhang, X. Flow-mediated interactions between two self-propelled flapping filaments in tandem configuration. Phys. Rev. Lett. 2014, 113, 238105. [Google Scholar] [CrossRef]
- Dai, L.; He, G.; Zhang, X.; Zhang, X. Stable formations of self-propelled fish-like swimmers induced by hydrodynamic interactions. J. R. Soc. Interface 2018, 15, 20180490. [Google Scholar] [CrossRef] [PubMed]
- Park, S.G.; Sung, H.J. Hydrodynamics of flexible fins propelled in tandem, diagonal, triangular and diamond configurations. J. Fluid Mech. 2018, 840, 154–189. [Google Scholar] [CrossRef]
- Kurt, M.; Mivehchi, A.; Moored, K.W. Two-dimensionally stable self-organization arises in simple schooling swimmers through hydrodynamic interactions. arXiv 2021, arXiv:2102.03571. [Google Scholar]
- Novati, G.; Verma, S.; Alexeev, D.; Rossinelli, D.; Van Rees, W.M.; Koumoutsakos, P. Synchronisation through learning for two self-propelled swimmers. Bioinspir. Biomim. 2017, 12, 036001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, M.; Iollo, A. Modeling and simulation of fish-like swimming. J. Comput. Phys. 2011, 230, 329–348. [Google Scholar] [CrossRef]
- Gazzola, M.; Hejazialhosseini, B.; Koumoutsakos, P. Reinforcement learning and wavelet adapted vortex methods for simulations of self-propelled swimmers. SIAM J. Sci. Comput. 2014, 36, B622–B639. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Chang, X.; Tian, R.; Wang, N.; Zhang, L.; Liu, W. A numerical simulation method for bionic fish self-propelled swimming under control based on deep reinforcement learning. Proc. Inst. Mech. Eng. Part C J. Mech. Eng. Sci. 2020, 234, 3397–3415. [Google Scholar] [CrossRef]
- Gazzola, M.; Tchieu, A.A.; Alexeev, D.; de Brauer, A.; Koumoutsakos, P. Learning to school in the presence of hydrodynamic interactions. J. Fluid Mech. 2016, 789, 726–749. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Novati, G.; Koumoutsakos, P. Efficient collective swimming by harnessing vortices through deep reinforcement learning. Proc. Natl. Acad. Sci. USA 2018, 115, 5849–5854. [Google Scholar] [CrossRef] [Green Version]
- Colabrese, S.; Gustavsson, K.; Celani, A.; Biferale, L. Flow navigation by smart microswimmers via reinforcement learning. Phys. Rev. Lett. 2017, 118, 158004. [Google Scholar] [CrossRef] [Green Version]
- Colabrese, S.; Gustavsson, K.; Celani, A.; Biferale, L. Smart inertial particles. Phys. Rev. Fluids 2018, 3, 084301. [Google Scholar] [CrossRef] [Green Version]
- Biferale, L.; Bonaccorso, F.; Buzzicotti, M.; Clark Di Leoni, P.; Gustavsson, K. Zermelo’s problem: Optimal point-to-point navigation in 2D turbulent flows using reinforcement learning. Chaos Interdiscip. J. Nonlinear Sci. 2019, 29, 103138. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Ling, F.; Heydari, S.; Kanso, E.; Heess, N.; Merel, J. Learning to swim in potential flow. Phys. Rev. Fluids 2021, 6, 050505. [Google Scholar] [CrossRef]
- Alageshan, J.K.; Verma, A.K.; Bec, J.; Pandit, R. Machine learning strategies for path-planning microswimmers in turbulent flows. Phys. Rev. E 2020, 101, 043110. [Google Scholar] [CrossRef] [PubMed]
- Tsang, A.C.H.; Tong, P.W.; Nallan, S.; Pak, O.S. Self-learning how to swim at low Reynolds number. Phys. Rev. Fluids 2020, 5, 074101. [Google Scholar] [CrossRef]
- Daddi-Moussa-Ider, A.; Löwen, H.; Liebchen, B. Hydrodynamics can determine the optimal route for microswimmer navigation. Commun. Phys. 2021, 4, 1–11. [Google Scholar]
- Zhu, Y.; Tian, F.B.; Young, J.; Liao, J.C.; Lai, J.C. A numerical study of fish adaption behaviors in complex environments with a deep reinforcement learning and immersed boundary-lattice Boltzmann method. Sci. Rep. 2021, 11, 1–20. [Google Scholar]
- Tian, F.B. A numerical study of linear and nonlinear kinematic models in fish swimming with the DSD/SST method. Comput. Mech. 2015, 55, 469–477. [Google Scholar] [CrossRef]
- Zhou, C.; Shu, C. Simulation of self-propelled anguilliform swimming by local domain-free discretization method. Int. J. Numer. Methods Fluids 2012, 69, 1891–1906. [Google Scholar] [CrossRef]
- Krüger, T.; Kusumaatmaja, H.; Kuzmin, A.; Shardt, O.; Silva, G.; Viggen, E.M. The Lattice Boltzmann Method; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Ma, J.; Wang, Z.; Young, J.; Lai, J.C.; Sui, Y.; Tian, F.B. An immersed boundary-lattice Boltzmann method for fluid-structure interaction problems involving viscoelastic fluids and complex geometries. J. Comput. Phys. 2020, 415, 109487. [Google Scholar] [CrossRef]
- Huang, W.X.; Tian, F.B. Recent trends and progress in the immersed Boundary method. Proc. Inst. Mech. Eng. Part C J. Mech. Eng. Sci. 2019, 233, 7617–7636. [Google Scholar] [CrossRef]
- Xu, Y.Q.; Tang, X.Y.; Tian, F.B.; Peng, Y.H.; Xu, Y.; Zeng, Y.J. IB–LBM simulation of the haemocyte dynamics in a stenotic capillary. Comput. Methods Biomech. Biomed. Eng. 2014, 17, 978–985. [Google Scholar]
- Tian, F.B.; Bharti, R.P.; Xu, Y.Q. Deforming-Spatial-Domain/Stabilized Space–Time (DSD/SST) method in computation of non-Newtonian fluid flow and heat transfer with moving boundaries. Comput. Mech. 2014, 53, 257–271. [Google Scholar] [CrossRef]
- Tian, F.B. FSI modeling with the DSD/SST method for the fluid and finite difference method for the structure. Comput. Mech. 2014, 54, 581–589. [Google Scholar] [CrossRef]
- Tian, F.B.; Wang, Y.; Young, J.; Lai, J.C. An FSI solution technique based on the DSD/SST method and its applications. Math. Model. Methods Appl. Sci. 2015, 25, 2257–2285. [Google Scholar] [CrossRef]
- Mittal, R.; Iaccarino, G. Immersed boundary methods. Annu. Rev. Fluid Mech. 2005, 37, 239–261. [Google Scholar] [CrossRef] [Green Version]
- Sotiropoulos, F.; Yang, X. Immersed boundary methods for simulating fluid–structure interaction. Prog. Aerosp. Sci. 2014, 65, 1–21. [Google Scholar] [CrossRef]
- Xu, L.; Tian, F.B.; Young, J.; Lai, J.C. A novel geometry-adaptive Cartesian grid based immersed boundary-lattice Boltzmann method for fluid–structure interactions at moderate and high Reynolds numbers. J. Comput. Phys. 2018, 375, 22–56. [Google Scholar] [CrossRef]
- Xu, L.; Wang, L.; Tian, F.B.; Young, J.; Lai, J.C. A geometry-adaptive immersed boundary-lattice Boltzmann method for modelling fluid–structure interaction problems. In IUTAM Symposium on Recent Advances in Moving Boundary Problems in Mechanics; Springer: Berlin/Heidelberg, Germany, 2019; pp. 161–171. [Google Scholar]
- Young, J.; Tian, F.B.; Liu, Z.; Lai, J.C.; Nadim, N.; Lucey, A.D. Analysis of unsteady flow effects on the Betz limit for flapping foil power generation. J. Fluid Mech. 2020, 902, A30. [Google Scholar] [CrossRef]
- Tian, F.B.; Luo, H.; Zhu, L.; Liao, J.C.; Lu, X.Y. An efficient immersed boundary-lattice Boltzmann method for the hydrodynamic interaction of elastic filaments. J. Comput. Phys. 2011, 230, 7266–7283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mnih, V.; Kavukcuoglu, K.; Silver, D.; Rusu, A.A.; Veness, J.; Bellemare, M.G.; Graves, A.; Riedmiller, M.; Fidjeland, A.K.; Ostrovski, G. Human-level control through deep reinforcement learning. Nature 2015, 518, 529. [Google Scholar] [CrossRef] [PubMed]
- Hausknecht, M.; Stone, P. Deep Recurrent Q-Learning for Partially Observable MDPs. In Proceedings of the 2015 AAAI Fall Symposium Series, Ithaca, NY, USA, 27 August 2015. [Google Scholar]
- Tampuu, A.; Matiisen, T.; Kodelja, D.; Kuzovkin, I.; Korjus, K.; Aru, J.; Aru, J.; Vicente, R. Multiagent cooperation and competition with deep reinforcement learning. PLoS ONE 2017, 12, e0172395. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Pang, J.-H.; Tian, F.-B. Stable Schooling Formations Emerge from the Combined Effect of the Active Control and Passive Self-Organization. Fluids 2022, 7, 41. https://doi.org/10.3390/fluids7010041
Zhu Y, Pang J-H, Tian F-B. Stable Schooling Formations Emerge from the Combined Effect of the Active Control and Passive Self-Organization. Fluids. 2022; 7(1):41. https://doi.org/10.3390/fluids7010041
Chicago/Turabian StyleZhu, Yi, Jian-Hua Pang, and Fang-Bao Tian. 2022. "Stable Schooling Formations Emerge from the Combined Effect of the Active Control and Passive Self-Organization" Fluids 7, no. 1: 41. https://doi.org/10.3390/fluids7010041
APA StyleZhu, Y., Pang, J.-H., & Tian, F.-B. (2022). Stable Schooling Formations Emerge from the Combined Effect of the Active Control and Passive Self-Organization. Fluids, 7(1), 41. https://doi.org/10.3390/fluids7010041