Abstract
Aquilaria sinensis (Lour.) Spreng is a known medicinal plant producing agarwood. To date, studies on the floral biology of A. sinensis have been limited. In this study, the floral micro- and ultra-structures, pollen viability, stigma receptivity, and artificial pollination of A. sinensis were investigated. The results show that the flower’s development can be divided into five stages, including the flower bud differentiation stage (2–7 d), the flower bud stage (7–13 d), the flowering stage (14 d), the pollination stage (14–15 d), and the fruiting stage (15–25 d). The floral organs mainly include: 4–6 split calyces, 10 petals, 10 stamens, and 1 pistil. The anther is oblong, with four pollen sacs the pollen is round, with maximum viability 6 h after flowering; and stigma receptivity is at its best 6 h before flowering. Artificial pollination is successful in the field. These findings will provide useful information for producing and breeding A. sinensis.
1. Introduction
Agarwood is a resin-containing wood formed by injured plants of the genus Aquilaria spp. or Gyrinops spp., and it has been widely used in medicine, perfume, and incense throughout the world, including in Asia, the Middle East, and Europe [,]. The wild resources of Aquilaria spp. are mainly distributed in Asia (i.e., Philippines, Indonesia, Malaysia, Thailand, Vietnam, and Myanmar), with 15 agarwood species in Southeast Asia and 3 in South Asia [,]. In China, Aquilaria sinensis and Aquilaria yunnanensis have been recorded in the wild, while A. sinensis has been widely cultivated as a unique agarwood source for medicine use [,]. Currently, wild A. sinensis is mainly distributed in the provinces of Guangdong, Guangxi, Fujian, and Hainan; it has been listed in the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) appendix, which largely results from its overexploitation [,,]. Since 2007, artificial cultivation of A. sinensis has been increasingly carried out in the tropical and subtropical areas of China (i.e., Guangdong, Guangxi, Hainan, Yunnan, and Fujian), while research on germplasm innovation and new variety selection has been performed [].
Unveiling floral biology (e.g., flower characteristics, longevity of flower organs, and degree of self-compatibility) plays an important role in a breeding system [,,]. The study of flowering habits is of great importance to hybrid breeding and superior germplasm breeding, as seen in pears, coconuts, and tea [,,,]. Previous studies have found that there are significant differences in floral biology among different Aquilaria spp. Specifically, there are significant differences in the duration of reproduction stages (i.e., the flower differentiation stage, the flowering stage, and the fruiting stage), with 44.6 d for A. filaria, 49.4 d for A. crassna, 51.4 d for A. microcarpa, 59.3 d for A.beccariana, 77.7 d for A. hirta, and 82.4 d for A. malaccensis []. A. sinensis is a terminal or axillary umbel, and the flowers are bisexual with four pollen sacs; generally, buds appear in March, flowers bloom in April, and fruit ripens from June to August [,]. The sexual reproduction of A. crassna occurs annually, and the flowers are hermaphroditic and borne on terminal branches [].
Self-incompatibility is one of the most common mechanisms used by plants to prevent self-fertilization and promote outcrossing, which can maintain heterozygosity and heterogeneity []. Additionally, the alternation of life cycles largely depends on sexual reproduction, for which normal flower development is necessary []. Previous studies have found that the pollination of A. crassna and seed dispersal of A. sinensis are carried out by hornets, noctuids, and pyralids [,,]. To date, studies on the floral biology of A. sinensis are still limited. In this study, the floral biology (i.e., flower development, structure of anthers and pollen, pollen viability, and stigma receptivity) was examined, from the flower differentiation stage to the fruiting stage.
2. Materials and Methods
2.1. Plant Material
Seed-propagated plants of Aquilaria sinensis (Lour.) Spreng are grown at the Xinglong Southern Medicine Garden (21 m a.s.l.; 18°44′ N, 110°11′ E; mean annual temperature 24.8 °C; mean annual rainfall 2400 mm) in Hainan province, China. In 2022, 30 plants were randomly selected for this study regarding floral biology, including flower development, structure of anthers and pollen, pollen viability, and stigma receptivity.
2.2. Floral Morphology and Anatomy
The flowers’ morphology was recorded using an camera (E-M5, Olympus, Hong Kong, China). The structure of the floral anatomy (i.e., stigmas, petals, and calyces) was observed using a stereomicroscope (SZ61-SET; Olympus).
2.3. Floral Micro-Structures
The flower samples collected from the plants were immediately stored in an FAA fixative with 90 mL of 50% alcohol, 5 mL of formalin, and 5 mL of glacial acetic acid. The samples were embedded and sliced into 4 μm slices. The samples were then flattened at 40 °C, retrieved, and baked at 60 °C. The dewaxing agent (G1128, Servicebio, Wuhan, China) was used to dewax, red dye solution (G1031, Servicebio, Wuhan, China) was used to stain, and gradient ethanol was used to prepare solid green dye solution (G1031,Servicebio, Wuhan, China). Finally, the samples were cleared with xylene for 5 min, sealed with a neutral gum sheet, and observed using an optical microscope (Eclipse E100, Nikon, Guangzhou, China).
2.4. Ultra-Structure of Anthers and Pollen
The flower samples containing anthers and pollen were firstly fixed with EM fixative solution (G1102-100 M, Servicebio, Wuhan, China) at 22 °C for 2 h. The fixed samples were washed with 0.1 mol/L phosphate buffer (pH 7.4) three times at 22 °C for 15 min and transferred into 1% OsO4 (Ted Pella Inc., Beijing, China) in 0.1 mol/L phosphate buffer (pH 7.4) at 22 °C for 2 h. Then, the samples were successively dehydrated with 30%, 50%, 70%, 80%, 90%, and 100% ethanol for 15 min, as well as isoamyl acetate for 15 min. Finally, the anthers and pollen were observed using a scanning electron microscope (SU8100, Hitachi, Shanghai, China).
2.5. Measurement of Pollen Viability
The pollen viability was measured using a TTC (2,3,5-triphenyl-2h-tetrazolium chloride) method []. Firstly, the newly collected pollen was briefly placed on a slide and immediately brought back to the lab; then, the collected pollen was stained with a 0.5% TTC solution, placed in an incubator at 35 °C for 15 min, and observed using a microscope.
2.6. Measurement of Stigma Receptivity
Stigma receptivity was measured using a biphenidine-hydrogen peroxide method []. The stigma was collected in the early flowering stage and briefly placed in a concave slide. Then, a mixed solution (benzidine: hydrogen peroxide: water = 4:11:22) was added to the slide, and stigma receptivity was observed using a microscope.
2.7. Validation of Artificial Pollination
In order to validate the possibility of artificial pollination, the stigma was pollinated by hand ca. 6 h before flowering. Firstly, 48 h after pollination, the petals and calyces were removed and the pistil was fixed with FAA for 24 h. The fixed samples were then washed with ddH2O water and immersed in 3 mol/L NaOH at 60 °C for 1.5 h. The samples were then washed with ddH2O water and stained with 0.1% aniline blue dye solution in the dark for 3 h. Then, the samples were pressed, imbibed with glycerol, and covered with a cover slip. Finally, the pistil was observed using an Otter optics BK-FL fluorescence microscope.
2.8. Statistical Analysis
All experiments were performed using three biological replicates. SPSS 22.0 software was used for ANOVA and Tukey′s post test, with p < 0.05 considered statistically significant between flower development stages.
3. Results
3.1. Floral Morphology at Different Development Stages
As shown in Figure 1, obvious changes in the floral morphology of A. sinensis were observed at different development stages. Based on the floral morphology, the flower’s development can be divided into five stages, including: the flower bud differentiation stage (2–7 d), the flower bud stage (7–13 d), the flowering stage (14 d), the pollination stage (14–15 d), and the fruiting stage (15–25 d).

Figure 1.
Floral morphology of Aquilaria sinensis at different development stages.
3.2. Floral Morphological Structure
A. sinensis inflorescences are arranged in an umbrella shape and positioned at the top or axillary of stems, with 4–15 flowers per inflorescence and a flower stalk length of 8.29 ± 1.58 mm (Table 1 and Figure 2a). There are 4–6 split calyces that are ovate, with an 8.42 ± 0.77 mm length (Table 1 and Figure 2b,g). There are 10 petals that are scaly and inserted at the calyces’ throat (Figure 2c,g). There are 10 stamens with a 4.81 ± 0.45 mm length and one pistil with a 3.83 ± 0.34 mm length (Table 1 and Figure 2c,g). After pollination, the anthers gradually wither (Figure 2d,e), the petals begin to close (Figure 2e), and the stigmas gradually wither (Figure 2f).

Table 1.
Floral organs of A. sinensis.

Figure 2.
Floral morphological structure of A. sinensis. (a) inflorescence, (b) flower, (c) longitudinal section of flower, (d) anther, (e) withered anther, (f) withered stigma, and (g) floral organs.
3.3. Micro-Structure of Stigmas and Anthers
As shown in Figure 3, significant changes in the structure of stigmas and anthers were observed during flowering. The stigma was white before flowering, turned yellow when pollinated after flowering for 1 h, and gradually lost water and withered after flowering for 4 days (Figure 3a–f). The anther stayed open before flowering, generated scattered pollen after flowering for 1 h, and the amount of pollen gradually decreased and withered after flowering for 4 days (Figure 3g–l).
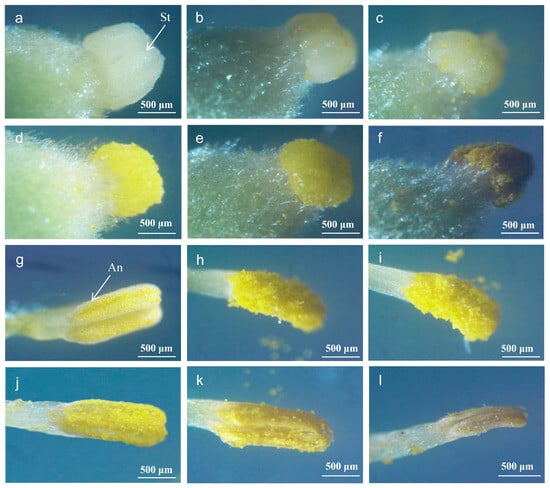
Figure 3.
Micro-structure of A. sinensis stigma and anther during flowering. An: anther, St: stigma. Stigma: (a) before flowering, (b) 1 h after flowering, (c) 6 h after flowering, (d) 12 h after flowering, (e) 24 h after flowering, and (f) 4 days after flowering. Anther: (g) before flowering, (h) 1 h after flowering, (i) 6 h after flowering, (j) 12 h after flowering, (k) 24 h after flowering, and (l) 4 days after flowering.
3.4. Micro-Structure of Stamens and Pistils
In the stamen’s flower bud differentiation stage, the flower bud’s central growth cone is surrounded by two to three small, round protrusions of the stamen primordium (Figure 4(a1)). In the pistil’s case, the central growth cone is surrounded by a ring-like protrusion of the pistil primordium; the ovary develops and expands, and the ovule primordium appears inside the ovary. The pistil primordium differentiates into flower primitives, such as petal primordium (Figure 4(b1)).
During the flower’s development, the stamen primordium differentiates to oblong anthers and short filaments, and the three layers of cells in the stamen primordium further divide and differentiate into different tissues. The outermost peripetalous cells expand the surface area of the anther and form its outermost epidermis; the intermediate layer of cells divides flatly and differentiates to form the most primitive sporogenic cells; and the primary sporogenic cells divide several times mitotically to form microspore mother cells. The bottom of the pistil gradually forms a cup-like structure before the ovary becomes egg-shaped (Figure 4(a2,b2)).
Once the anther has developed, a bidirectional structure of locules, walls, and vascular zones are formed in the anther (Figure 4(a3)). After meiosis forms a tetrad, the four cells separate and become free in the pollen sac through lysis of the callus wall, forming a single, thin-walled pollen grain with a thick cytoplasm and a centrally located nucleus. The cells of the chorionic felt layer undergo vesiculation and begin to autolyze in situ. The pollen is now fully mature, and the pollen sac tapetum cells are completely lyzed (Figure 4(a3–a5)). The ovules within the ovary form and gradually mature on one stigma (Figure 4(b3,b4)).
At the flowering stage, the ovules inside the ovary develop and mature, and the anthers begin to release pollen on top of the stigma (Figure 4(a6,b5,b6)).
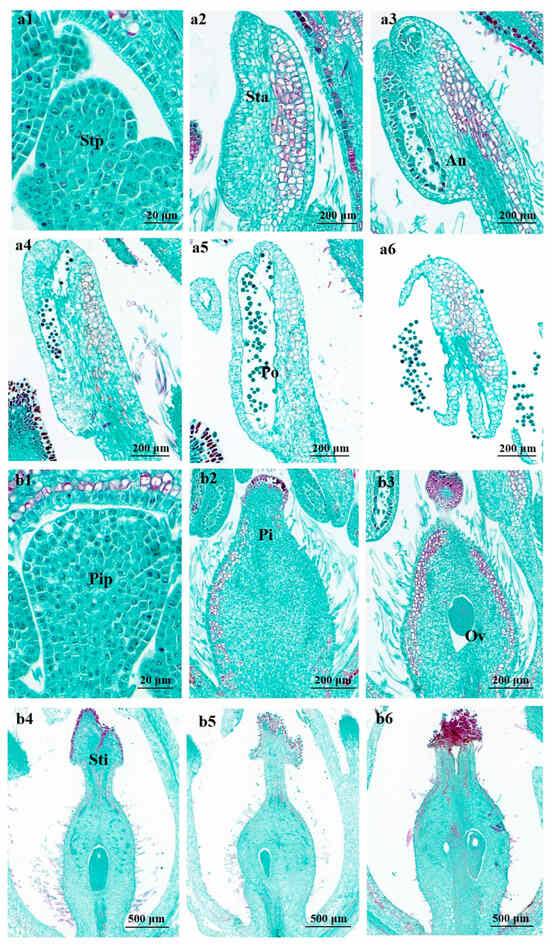
Figure 4.
Micro-structure of stamen and pistil of A. sinensis during flowering. Stp: stamen primordium, Sta: stamen, Sti: stigma, An: anther, Po: pollen, Pip: pistil primordium, Pi: pistil, Ov: ovary. Image a and b represent a stamen and pistil, respectively: (a1,b1) flower bud differentiation at 2 days and (a2–a6, b2–b6) flower bud differentiation at 7 days after flowering 2 days.
3.5. Ultra-Structure of Anthers and Pollen
The anthers are oblong with a 1.3 mm length and four pollen sacs (Figure 5a). The pollen grain is round with a 22 μm diameter (Figure 5b); the outer surface has a reticulate pattern, with cone-like protrusions, an acuminate apex, and a round and obtuse base (Figure 5c).

Figure 5.
Ultra-structure of anther and pollen of A. sinensis. (a) anther, (b) pollen grain, and (c) surface of pollen grain.
3.6. Pollen Viability during Flowering
The pollen viability was 45.30% at 0 h, significantly increased and maximized to 88.67% at 6 h, and then significantly decreased from 59.00% at 12 h to 0.00% at 20 h (Figure 6). This indicates that the maximum viability of pollen is at ca. 6 h, and the pollen loses viability after flowering for 20 h. Since the fruit set largely depends on pollen viability [], the maintenance of pollen viability plays an important role in the pollination reproduction of A. sinensis.
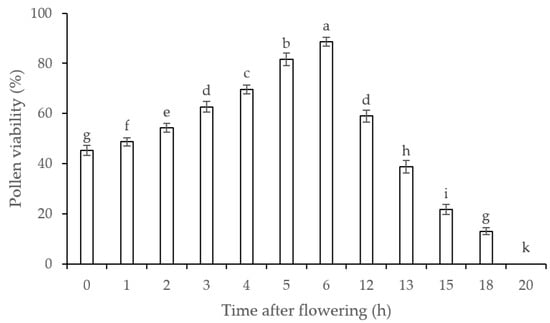
Figure 6.
Pollen viability of A. sinensis after flowering. Values are averages with their standard deviations (n = 3). Different lowercase letters represent a significant difference (p < 0.05) among different treatments.
3.7. Stigma Receptivity during Flowering
Weak stigma receptivity was measured by the presence of small bubbles and a light-blue stain before 24 h of flowering (Figure 7a); strong stigma receptivity was measured by the presence of many bubbles and a blue stain before 12 h of flowering (Figure 7b); the best stigma receptivity was measured by the presence of the most bubbles and a dark-blue stain before 6 h of flowering (Figure 7c). Stigma receptivity became weak, with fewer bubbles present, after 6 h of flowering (Figure 7d).
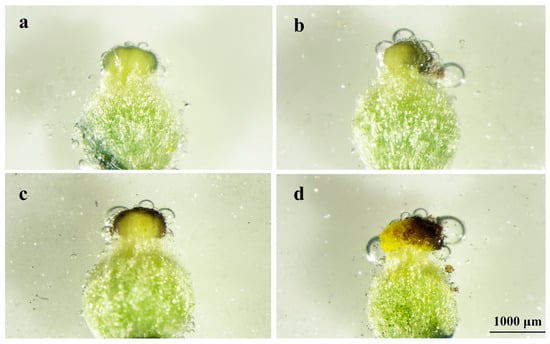
Figure 7.
Stigma receptivity of A. sinensis after flowering. (a) 24 h before flowering, (b) 12 h before flowering, (c) 6 h before flowering, and (d) 6 h after flowering.
3.8. Comparison of Artificial Pollination with Natural Pollination
As shown in Figure 8, the fluorescent micro-structure of pistils 48 h after natural and artificial pollination were both clearly observed. This proves that artificial pollination can be successfully applied in the reproduction of A. sinensis.
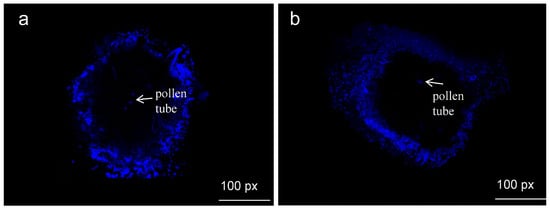
Figure 8.
Micro-structure of natural and artificial pollination of A. sinensis after 48 h. (a) natural pollination, (b) artificial pollination.
4. Discussion
4.1. Optimal Pollination Stage for A. sinensis
Many species of flowering plants use crossing to produce hybrid offspring in nature, which is borrowed in artificial pollination processes to generate novel cultivars with advantageous desired agronomic traits or tolerance to abiotic and biotic stress. [] The duration of pollen viability and stigma receptivity is closely related to species []. The pollen of passion-fruit display some viability 5 h before flowering; then, pollen viability increases continuously, reaching a peak 3 h after flowering, and then drops to only 10.63% at 9 h and to 0% at 20 h []. The pollen viability of Cymbidium hybridum lasts for 35 days []. In our study, the A. sinensis pollen had lost its viability 20 h after flowering; in other words, the pollen lifespan was relatively short. The scanning electron microscopy image showed no significant change in pollen appearance during the test period of this study, indicating that pollen viability could not be judged by observing its appearance alone.
The stigma is the best site for examining the hydration and germination of pollen, which have an important influence on fruit setting and are also important factors restricting fruit setting. There are also large time differences in stigma receptivity loss: with Artemisia wudanica T. N., Liou & W. Wang retained its stigma receptivity for 6 days after flowering []; C. hybridum retained its stigma receptivity for 45 days after flowering []; and Lilium davidii var. unicolour Cohan lost stigma receptivity the day of flowering []. The stigmas of A. sinensis were already receptive before flowering. Before and after flowering, a weak–strong–weak receptivity pattern was observed. The fruit setting rate was strongly correlated with pollen viability, indicating that both pollen viability and stigma receptivity had a significant effect on the fruiting rate of A. sinensis. In addition to the influence of pollen and stigmas, other factors can have important effects on the fruit setting rate, such as temperature and humidity.
Studying the pollen viability and stigma receptivity can improve the fruit setting rate, which also has an important impact on pollination efficiency and is of great significance in artificial selection and hybrid breeding. The study of plant flower structures and reproductive systems is essential for understanding plant life history and for developing a fundamental basis for research []. For seed plants, the necessary stage of fertilization is pollination, and plant characteristics such as corolla size, pistil length, and anther size affect the reproductive success of the plant [].
4.2. Pollination Characteristics of A. sinensis
The structure and function of breeding systems and life history characteristics are the two main directions of research on plant evolution and ecology. Plant breeding systems affect the sexual characteristics of offspring through the flower pattern and opening mode and the sequence and duration of pistil opening. Adaptation to different mating systems, degree of self-compatibility, and mating systems are the core parts of a plant’s breeding system. Diverse floral expressions and breeding systems often occur in the same taxa (species, genera, families, and orders) or in different geographical regions for the same taxa [,,,].
The breeding organs of plants adapt to pollinators through flower design and flower display and promote successful mating between plants. Wind, water, and insects are the main media for outcross pollination. Previous studies have revealed that most members of the family Thymelaeaceae are entomophilous []. Aquilaria sinensis is a cross-pollinating plant that requires pollinators. Previous studies have found that noctuids and pyralids are pollinators of pollinator-dependent A. sinensis []. We found many thrips in the flower of A. sinensis. At the same time, thrips were reported as pollinators in other species of Aquilaria [].
4.3. Artificial Pollination and Compatibility of A. sinensis
Self-incompatibility, or the inability of a fertile seed plant to produce a zygote after self-pollination, serves as a widely occurring outcrossing mechanism to prevent inbreeding in angiosperms. To adapt to cross-pollination, plants have evolved many mechanisms to avoid selfing [,]. Higher levels of selfing and outcrossing between related individuals will reduce the effective size of the population, thus increasing inbreeding and reducing the relative fitness of inbred offspring, which is called inbreeding depression []. Inbreeding depression has obvious effects on fruiting, seed weight, germination time, and germination rate, among other factors. For example, selfing can lead to a decrease in the quantity and quality of seed setting []. It is known that A. filaria, A. malaccensis, and A. microcarpa have self-incompatibility []. According to our study, artificial pollination of a stamen 6 h after flowering and of a pistil 6 h before flowering can improve the fruit setting rate. However, we found no fruit for self-pollinated A. sinensis, so we speculate that A. sinensis also has self-incompatibility characteristics.
5. Conclusions
From the above investigations, the floral biology of Aquilaria sinensis exhibits unique characteristics, such as its flower development, which can be divided into five stages; floral organs including calyces, petals, stamens, and pistil; and the presentation of pollen viability and stigma receptivity in a timely manner. Artificial pollination is successful in this field. These findings will provide useful information for producing and breeding A. sinensis.
Author Contributions
The contributions of the respective authors are as follows: Conceptualization, B.W., Y.Y. and J.W.; Data curation, B.W., G.M., S.L. and X.H.; Formal analysis, B.W. and G.M.; Funding acquisition, B.W.; Investigation, B.W., G.M., X.H., B.C., H.L. and L.H.; Methodology, B.W. and Y.Y.; Project administration, B.W.; Resources, Y.Y.; Software, G.M.; Supervision, Y.Y. and J.W.; Validation, G.M.; Writing—original draft, B.W., G.M., S.L. and X.H.; Writing—review and editing, Y.Y. and J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Hainan Provincial Natural Science Foundation of China (321QN0946), the CAMS Innovation Fund for Medical Sciences (CIFMS 2021-I2M-1-032), and the National Key Research and Development Program of China (2022YFC3501504).
Data Availability Statement
Data are contained within the article.
Acknowledgments
Hui Meng (Hainan Branch of the Institute of Medicinal Plant Development, China) assisted in data analysis, and Mengfei Li (Gansu Agricultural University, China) revised the manuscript. The authors would like to thank these experts for their kind help.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Soehartono, T.; Newton, A.C. Reproductive ecology of Aquilaria spp. in Indonesia. For. Ecol. Manag. 2001, 152, 71. [Google Scholar] [CrossRef]
- Naef, R. The volatile and semi-volatile constituents of agarwood, the infected heartwood of Aquilaria species: A review. Flavour Fragr. J. 2011, 26, 73–87. [Google Scholar] [CrossRef]
- Lee, S.Y.; Ng, W.L.; Mahat, M.N.; Nazre, M.; Mohamed, R. DNA Barcoding of the Endangered Aquilaria (Thymelaeaceae) and Its Application in Species Authentication of Agarwood Products Traded in the Market. PLoS ONE 2016, 11, e0154631. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zhao, W.; Sun, P.; Wei, J. Species and Conservation Status of the Endangered Agarwood-Producing Genus Aquilaria. Mod. Chin. Med. 2017, 19, 1057–1063. [Google Scholar]
- Committee of Chinese Flora. Flora of China; Science Press: Beijing, China, 1999; pp. 289–290. [Google Scholar]
- Xu, Y.H.; Zhang, Z.; Wang, M.X.; Wei, J.H.; Chen, H.J.; Gao, Z.H.; Sui, C.; Luo, H.M.; Zhang, X.L.; Yang, Y.; et al. Identification of genes related to agarwood formation: Transcriptome analysis of healthy and wounded tissues of Aquilaria sinensis. BMC Genom. 2013, 14, 227. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.W.; Zhao, J.P.; Liu, Y.B.; Qiu, Y.X.; Xie, Q.L.; Li, M.J.; Khan, I.A.; Wang, W. Advance in studies on chemical constituents, pharmacology and quality control of Aquilaria sinensis. Digit. Chin. Med. 2018, 1, 316–330. [Google Scholar] [CrossRef]
- Li, L.H.; Shou, H.Y.; Ma, Q.W. Distribution of Aquilaria sinensis (Thymelaeaceae). Med. Plant 2012, 3, 8. [Google Scholar]
- CITES. Available online: http://checklist.cites.org (accessed on 8 December 2023).
- Zhou, Y.F.; Wei, J.H.; Sun, P.W.; Liu, Y.Y.; Yang, Y.; Liu, P.W.; Ding, Z.M.; Wang, S.; Wang, C.H. The Development Situation and Consideration of Agarwood Scientific & Technological Innovation and Industrial Development in China. Mod. Chin. Med. 2017, 19, 1051–1056. [Google Scholar]
- Wyatt, R. Pollinator-plant interactions and the evolution of breeding systems. In Pollination Biology; Academic Press: Cambridge, MA, USA, 1983; pp. 51–86. [Google Scholar]
- Lan, Q.Y.; Fang, C.Y.; He, H.Y.; Chen, Z.Y. Mature embryo culture and plantlet regeneration of the Aquilaria sinensis. Genom. Appl. Biol. 2001, 20, 231–232. [Google Scholar]
- Friedman, J. The evolution of annual and perennial plant life histories: Ecological correlates and genetic mechanisms. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 461–481. [Google Scholar] [CrossRef]
- Hathurusinghe, B.M.; Pushpakumara, D.K.N.G.; Bandaranayake, P.C.G. Macroscopic and microscopic study on floral biology and pollination of Cinnamomum verum Blume (Sri Lankan). PLoS ONE 2023, 18, e0271938. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.L.; Wei, Y.R.; Pang, H.G.; Xu, J.F.; Li, Y.L.; Zhang, H.X.; Zhang, J.G.; Zhang, Y.X. Genome-wide identification of the PEBP genes in pears and the putative role of PbFT in flower bud differentiation. PeerJ 2020, 8, e8928. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, T.; Hatta, M.A.M. Improving coconut using modern breeding technologies: Challenges and opportunities. Plants 2022, 11, 3414. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hao, X.Y.; Lu, Q.H.; Zhang, W.F.; Zhang, H.J.; Wang, L.; Yang, Y.J.; Xiao, B.; Wang, X.C. Genome-wide identification and expression analysis of flowering-related genes reveal putative floral induction and differentiation mechanisms in tea plant (Camellia sinensis). Genomics 2020, 112, 2318–2326. [Google Scholar] [CrossRef] [PubMed]
- Shou, H.Y.; Ma, Q.W.; Liu, D.; Li, F.L. Cytological observations of microsporogenesis and male gametophyte development of Aquilaria sinensis (Lour.) Spreng. J. Beijing For. Univ. 2010, 32, 92–96. [Google Scholar]
- Zheng, L.A.; Wang, L.Y.; Han, R.Q.; Li, W.J.; Cheng, X.L. Growth Characteristics and Planting Techniques of Aquilaria sinensis. South China Agric. 2021, 15, 27–29. [Google Scholar]
- López-Sampson, A.; Page, T. Reproductive phenology, floral biology and breeding system of aquiliaria crassna in tropical Australia. J. Trop. For. Sci. 2019, 31, 211–221. [Google Scholar] [CrossRef]
- Muenchrath, D.; Campbell, A.; Merrick, L.; Lübberstedt, T.; Fei, S.Z. Controlled Hybridization: Self-incompatibility, Male-sterility, and Sex-inheritance. In Crop Genetics, 3rd ed.; Walter, S., Kendall, L., Eds.; Iowa State University Digital Press: Ames, IA, USA, 2023; pp. 42–66. [Google Scholar]
- Wei, S.; Ma, L. Comprehensive Insight into Tapetum-Mediated Pollen Development in Arabidopsis thaliana. Cells 2023, 12, 247. [Google Scholar] [CrossRef]
- Qin, R.M.; Wen, P.; Corlett, R.T.; Zhang, Y.Y.; Wang, G.; Chen, J. Plant-defense mimicry facilitates rapid dispersal of short-lived seeds by hornets. Curr. Biol. 2022, 32, 3429–3435. [Google Scholar] [CrossRef]
- Peakall, R.; Bohman, B. Seed dispersal: Hungry hornets are unexpected and effective vectors. Curr. Biol. 2022, 32, R836–R838. [Google Scholar] [CrossRef]
- Chen, G.; Liu, C.; Sun, W. Pollination and seed dispersal of Aquilaria sinensis (Lour.) Gilg (Thymelaeaceae):an economic plant species with extremely small populations in China. Plant Divers. 2016, 38, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q. Experimental Guidance of Plant Physiology; China Agricultural Press: Beijing, China, 2000; p. 149. [Google Scholar]
- Dafni, A. Pollination Ecology; Oxford University Press: New York, NY, USA, 1992; pp. 1–57. [Google Scholar]
- Thu, M.K.; Lin, Y.; Chen, J. Flower Types, Pollen Morphology, and In Vitro Pollen Germination of Longan (Dimocarpus longan Lour.). J. Bot. Res. 2017, 1, 50–56. [Google Scholar]
- Liang, S.; Hu, M.; Lin, H.C.; He, H.J.; Ning, X.P.; Peng, P.P.; Lu, G.H.; Sun, S.L.; Wang, X.J.; Wang, Y.Q.; et al. Transcriptional regulations of pollen tube reception are associated with the fertility of the ginger species Zingiber zerumbet and Zingiber corallinum. Front. Plant Sci. 2023, 14, 1099250. [Google Scholar] [CrossRef] [PubMed]
- Shao, F.X.; Wang, S.; Chen, J.H.; Chen, J.; Hong, R.Y.; Tang, Y.; Wang, J. Stigma shape development and receptivity of Zhongqiu Sucui Chinese jujube. Acta Hortic. Sin. 2019, 46, 2309–2322. [Google Scholar]
- Cai, Z.Y.; Dong, L.; Wang, H.Q.; Qiu, W.W.; Su, W.Q.; Ren, H.; Wang, X.M.; Fang, W.K.; Huang, Z.B.; Deng, B.; et al. Pollen viability, stigma receptivity and their effect on fruit set of passion-fruit at different flower developmental stages. J. Fruit Sci. 2023, 40, 969–977. [Google Scholar]
- Chu, Y.; Fan, Y.R.; Zhang, S.Y.; Sun, Y.F.; Ning, H.J. Pollen vitality and stigma receptivity of Cymbidium hybridum and Chinese orchid. J. Zhejiang A F Univ. 2013, 30, 950–954. [Google Scholar]
- Liu, H.; Cai, P.; Wan, T.; Yi, W.; Zhang, J. Observation on the Flowering Dynamic, Pollen Viability and Stigma Receptivity of Artemisia wudanica. Chin. J. Grassl. 2020, 42, 43–47. [Google Scholar]
- Sun, H.; Shi, G.; Ran, B.; Zhang, L.; Jia, X. Pollen viability, stigma receptivity and breeding system of Lilium davidii var. unicolor. J. Desert Res. 2019, 39, 62–69. [Google Scholar]
- Liu, L.D.; Zhu, N.; Shen, J.H.; Zhao, H.X. Comparative studies on floral dynamics and breeding system between Eleutherococcus senticosus and E. sessiliflorus. Acta Ecol. Sin. 2002, 22, 1041–1048. [Google Scholar]
- Machado, I.C.; Lopes, A.V. Floral traits and pollination systems in the Caatinga, a Brazilian tropical dry forest. Ann. Bot. 2000, 94, 365–376. [Google Scholar] [CrossRef]
- Charlesworth, D. Evolution of Plant Breeding Systems. Curr. Biol. 2006, 16, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Richards, A.J. Plant Breeding Systems; Hong Kong Printed in Great Britain at The University Press: Hong Kong, China, 1997; pp. 20–23. [Google Scholar]
- He, Y.P.; Liu, J.Q. A review on recent advances in the studies of plant breeding system. Acta Phytoecol. Sin. 2003, 27, 151–163. [Google Scholar]
- Armbruster, W.S.; Baldwin, B.G. Switch from specialized to generalized pollination. Nature 1998, 394, 632. [Google Scholar] [CrossRef]
- Wattanachai, T.; Suwan, T.; Malaiporn, T.; Kazuo, O. Insect pollination of Aquilaria crassna (Thymelaeaceae): Effect of moths for the fruit setting in Thailand. J. Fac. Agric. Kyushu Univ. 2009, 54, 321–328. [Google Scholar]
- Soehartono, T.; Newton, A.C. Conservation and sustainable use of tropical trees in the genus Aquilaria II. The impact of gaharu harvesting in Indonesia. Biol. Conserv. 2001, 97, 29–41. [Google Scholar] [CrossRef]
- Fattorini, R.; Glover, B.J. Molecular mechanisms of pollination biology. Annu. Rev. Plant Biol. 2020, 71, 487–515. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Kubo, K.; Takayama, S. Non-self- and self-recognition models in plant self-incompatibility. Nat. Plants 2016, 2, 16130. [Google Scholar] [CrossRef]
- Dyer, W.T.T. The Effects of Cross and Self-Fertilisation in the Vegetable Kingdom. Nature 1877, 15, 329–332. [Google Scholar] [CrossRef]
- Achrem, M.; Stępień, E.; Kalinka, A. Epigenetic Changes Occurring in Plant Inbreeding. Int. J. Mol. Sci. 2023, 24, 5407. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).