Abstract
The effect of N-(2-chloro-4-pyridyl)-N′-phenylurea (CPPU) on fruit splitting of ‘Murcott’ tangor fruit (Citrus reticulate × Citrus sinensis) was analyzed throughout two successive seasons during 2020 and 2021. ‘Murcott’ trees were treated with various concentrations of CPPU, 1.0, 5.0, and 10.0 mg·L−1, which significantly reduced the splitting of fruit compared with the control fruit and the gibberellic acid (GA3)-treated fruit (20.0 mg·L−1). The fruit treated with 5.0 mg·L−1 CPPU exhibited higher levels of fruit weight, longitudinal diameter, transverse diameter, peel strength, and peel thickness compared to the control fruit. In addition, the tree sprayed with 5.0 mg·L−1 CPPU had the greatest number of cellular layers and cells at the stylar end calculated in fruit. However, there were no other fruit quality characteristics significantly affected by CPPU and GA3 treatments.
1. Introduction
Fruit splitting has been a long-term pre-harvest problem in the citrus industry, especially observed in some split-susceptible species, such as Clementine ‘Ellendale’ [1], ‘Fino’ and ‘Marisol’ [2], sweet orange ‘Navel’ [3], ‘Valencia’ [4], and the mandarin hybrids ‘Murcott’ [5], ‘Nova’ [6], and ‘Shogun’ [7]. Splitting causes about a 30% loss of citrus production [8,9]. Although these are split-prone cultivars, the morphological characteristics resulting in fruit splitting differ and can be generalized as fruit with thin peel [2,3,4,5], fruit with secondary fruitlets [1,6,10,11], and fruit with a particular oblate shape [3,12]. Therefore, the structural integrity of fruit peel is hampered under internal pressure during the fruit developmental stage. It exhibits a split at the stylar end or navel end because these two spots have the weakest structures [13].
Few studies have been conducted on increasing the peel thickness and strength of split-susceptible citrus species by applying mineral nutrients or plant growth regulators (PGRs). Previous studies indicated that synthetic auxin, 2,4-dichlorophenoxy acetic acid (2,4-D), 1-naphthaleneacetic acid (NAA), S-abscisic acid (S-ABA), aminoethoxyvinylglycine (AVG), 1-methylcyclopropene (1-MCP), 3,5,6-trichloro-2-piridil oxyacetic acid (3,5,6-TPA), and gibberellic acid (GA3) are effective in terms of reducing citrus fruit splitting [2,6,8,10,14,15]. An earlier investigation has proven that an increase in endogenous GA3 and cytokinin led to thicker and rougher peel found in ‘Shamouti’ orange [16]. Mupambi et al. [2] pointed out that both 15 and 25 mg·L−1 of 2,4-D effectively reduced fruit splitting in ‘Marisol Clementine’ mandarin; however, these two concentrations also caused peel coarseness which affected the appearance of the fruit. In addition, Stander et al. [17] indicated that foliar application of 10 mg·L−1 2,4-D after the fruit drop only increased peel thickness and strength, resulting in a 50% reduction in fruit splitting in ‘Marisol’ and ‘Mor’, and had no effects on peel coarseness.
N-(2-chloro-4-pyridyl)-N′-phenylurea (CPPU), a synthetic inhibitor of cytokinin oxidase, has been used pre-harvest in horticultural crops to increase fruit size [18,19], to maintain the vivid green bract of red-fleshed pitaya (Hylocereus sp.) [20], and to promote precocious flowering in H. undatus [21]. The endogenous cytokinin was reported to affect the peel thickness of orange [16], and ‘Murcott’ is susceptible to splitting due to its thin peel characteristic [5]. Thus, this study aimed to evaluate the effect of the pre-harvest application of CPPU on ‘Murcott’ fruit splitting over two production seasons.
2. Materials and Methods
2.1. Site Description
The study was carried out in a commercial ‘Murcott’ orchard in Gukeng Township, Yunlin County, Taiwan (lat. 23.37° N, long. 120.35° E, elevation 50 m), throughout two consecutive seasons (2020–21 and 2021–22). Fifteen-year-old ‘Murcott’ (Citrus reticulate × Citrus sinensis) trees were spaced 4.0 × 2.5 m between and within rows. This area has a sub-tropical climate with annual precipitation between 1.5 and 2.0 m. The experimental trees received the same horticultural practices except for the differing experimental treatments throughout the study.
2.2. Field Experiment
The aqueous solution was made containing different CPPU (Kyowa Hakko Kogyo Co., Tokyo, Japan) concentrations (1.0, 5.0, and 10.0 mg·L−1), and water and GA3 (20.0 mg·L−1) were used as the negative–positive controls. The whole tree was directly foliar-sprayed using a power knapsack sprayer with 5.0 L·tree−1 every 7 days, three weeks in a row after full bloom (AFB, late March to early April). The trees chosen were uniform in size and vigor, and the treatments were applied to the same trees in the next season. A single tree was treated as an experimental unit, and the experimental design was a completely randomized design (CRD) with four replications.
2.2.1. Fruit Splitting Calculation
After the physiological fruit drop, each experimental unit was randomly tagged with 200 fruit per tree and monitored every week (starting on 1 July) until 31 December, which is represented in Figure 1. The splitted fruit were picked and collected each month, and the degree of fruit splitting was presented as the number of split fruit of the total tagged fruit (number of split fruit/total tagged fruit) × 100%.

Figure 1.
Different foliar applications on ‘Murcott’ fruit in field. (A) Control, (B) GA3, (C) 1.0 mg L−1 CPPU, (D) 5.0 mg L−1 CPPU, and (E) 10.0 mg L−1 CPPU.
2.2.2. Fruit Size
Twenty tagged fruit were randomly marked and measured in terms of fruit length (L) (mm) and fruit width (W) (mm) with a digital electronic caliper ruler (Mitutoyo 530-115, Kawasaki, Japan) every month until harvest. The average size of twenty fruit in each treatment was considered the fruit size of each replication.
2.2.3. Fruit Fresh Weight
The 20 fruit per treatment (as described in Section 2.2.2) were harvested on the same day and transported to the lab (RT at 25 °C and RH 50%) immediately for physiochemical analysis. The fruit were weighed using a digital balance (SCALTEC SBA-51, Goettingen, Germany), and the average was calculated.
2.2.4. Peel Strength
Five of the twenty tagged fruit were randomly picked for fruit peel strength measurement, and each fruit was placed on a stand of a texture analyzer (Stevens-LFRA, Middleboro, MA, USA). Peel strength was measured using a penetrating probe (2.0 mm) at a puncturing speed of 5.0 mm·s−1 and a 5.0 mm depth at one point around the equator and one point at the stylar end, and the readings were averaged and expressed in units as kg·cm−2.
2.2.5. Peel Thickness
Another ten fruit were used to assess peel thickness, with the fruit cut in half longitudinally, and peel thickness was measured using a digital electronic caliper ruler (Mitutoyo 530-115, Kawasaki, Japan) at the equator and stylar end. The average thickness (mm) of the ten fruit per treatment was obtained.
2.2.6. Total Soluble Solid (TSS), Titratable Acidity (TA), and TSS to TA Ratio
The same samples used in Section 2.2.5 were further analyzed for chemical characteristics. Each fruit was squeezed with a handheld juicer and a few drops of juice from each fresh fruit were placed on a refractometer (ATAGO PAL-1, Tokyo, Japan); ten readings were averaged per treatment. The total soluble solid (TSS) value was expressed in degrees Brix (°Brix). The titratable acidity (TA) was calculated by titrating fruit juice with 0.1 N NaOH to an end point of pH 8.2, and it was expressed as a percentage of citric acid [22]. The TSS to TA ratio was calculated by dividing the TSS value by the TA value.
2.3. Peel Tissue Sectioning
The last five fruit from each treatment and the split fruit were used and modified for the free tissue section [23]. The pericarp sample of the stylar end of each fruit was taken with dimensions of 0.2 × 0.2 × 0.2 cm3. First, the samples were fixed by 70% formalin–acetic acid–alcohol (5.0 mL formalin + 5.0 mL acetic acid + 70.0 mL alcohol, FAA). Next, all samples were immersed in a graded series of ethanol-tert-butanol solutions, and pure, water-free alcohol was added for dehydration. Third, the samples were infiltrated with a suitable histological wax at 60 °C in the oven. Fourth, each specimen was consecutively sectioned into 9 ± 2 μm thickness with a rotary microtome (Olympus Microm HM315, Boise, ID, USA). Fifth, each section was placed on the slide for mounting. The xylene-free specimen was stained with Safranin-O and Fast-green.
2.4. Quantitative Analysis of Sectioned Tissue
All slides were viewed under an optical microscope (Olympus BX51, Tokyo, Japan), and images were pictured by using a digital camera (Olympus E330, Tokyo, Japan) and magnified at 300% to calculate the cellular layers and cells using ImageJ software (https://ij.imjoy.io/, accessed on 30 August 2024).
2.5. Statistical Analysis
The data were subjected to analysis of variance (ANOVA) by using SAS (version 9.2; SAS Institute, Cary, NC, USA). Mean values were separated by using a least significant difference (LSD) test at a 5% significance level (p ≤ 0.05).
3. Results
3.1. Fruit Splitting Rate
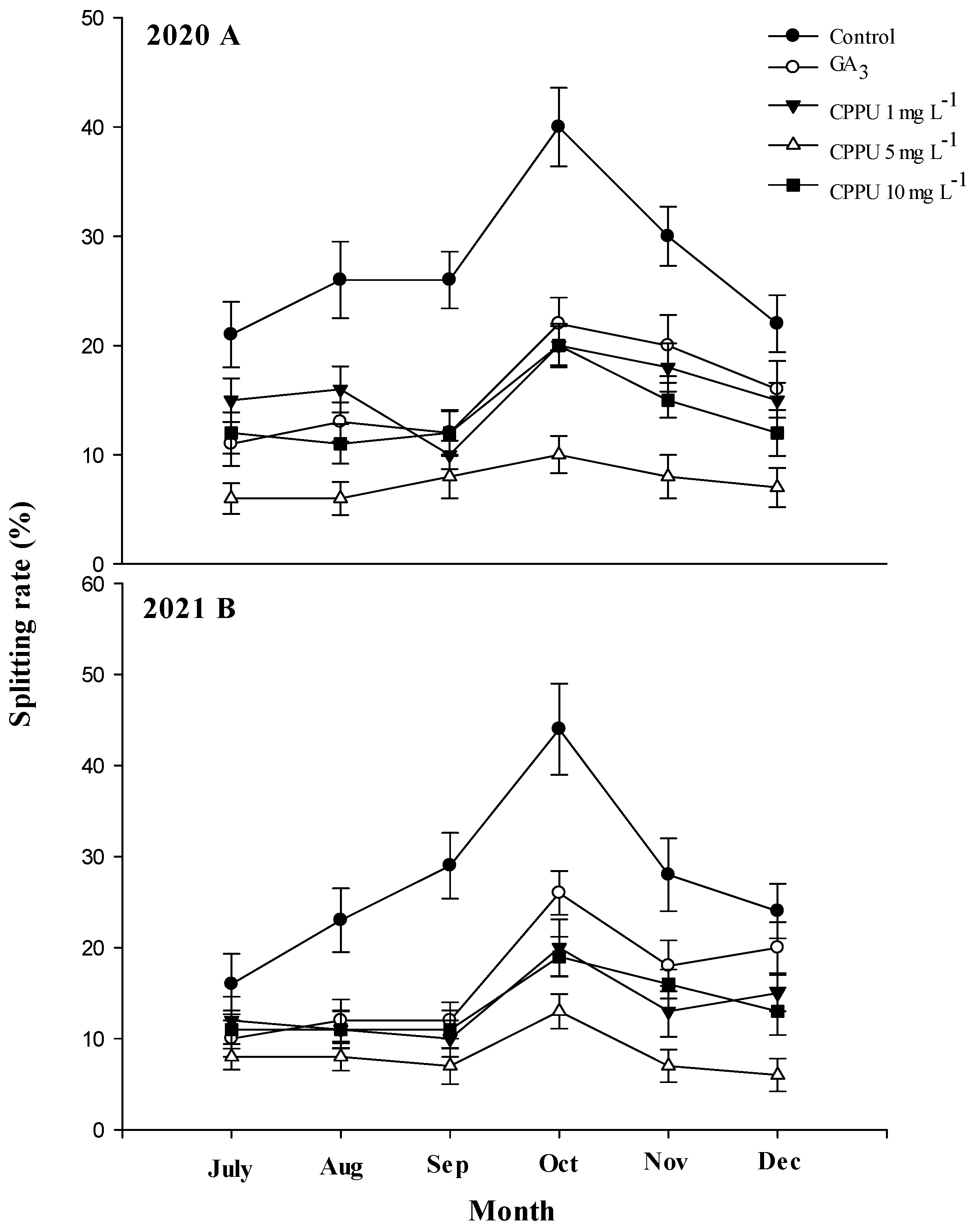
The occurrence of splitting fruit was observed from July to December in 2020 and 2021. In both years, the highest fruit splitting ratio was calculated in October in all treatments. A significant reduction in fruit splitting was observed in both years when ‘Murcott’ trees were treated with CPPU and GA3. However, the negative control trees, grown without CPPU, exhibited the highest fruit splitting ratios (Figure 2). Although CPPU and GA3 significantly reduced the percentage of fruit splitting compared with the non-treated trees, the trees treated with 5.0 mg·L−1 CPPU showed a greater reduction in fruit splitting than all other treatments (Figure 2). The water-treated trees had an average of 27.5% fruit splitting in 2020 compared with 8.8% in the trees treated with 5.0 mg·L−1 CPPU. Similarly, an average of 34.3% splitting was calculated from the water-treated trees compared with an average of 7.5% splitting when CPPU was applied at 5.0 mg·L−1 in 2021.

Figure 2.
The average split rate in ‘Murcott’ fruit in 2020 (A) and 2021 (B). The bars represent the means of four replications ± S.E. (n = 4).
3.2. Physical Characteristics
Fruit weight increased when the trees were subjected to all chemical treatments. However, the application of CPPU (5.0 mg·L−1) resulted in a significant increase in fruit weight compared to the water-treated trees in 2020 and 2021 (Table 1). The trees sprayed with 5.0 mg·L−1 CPPU had the greatest fruit length in 2020, but the greatest fruit length was found when the trees were treated with GA3 in 2021. In both years, the greatest fruit width and peel thickness were obtained under the 5.0 mg·L−1 CPPU treatment. In addition, a significantly higher mean value of peel strength was found in the trees treated with 5.0 mg·L−1 CPPU compared with the water-treated trees in both years. The maximum peel strength was observed in the trees subjected to 5.0 mg·L−1 CPPU in 2020 and 2021 (Table 1).

Table 1.
Effects of CPPU and GA3 on physicochemical characteristics of ‘Murcott’ fruit.
3.3. Chemical Characteristics
CPPU did not significantly affect TSS in ‘Murcott’ fruit in 2020 and 2021 (Table 1). In 2020, the lowest level of this characteristic was recorded when CPPU was applied at 1.0 mg·L−1 and the lowest TSS was recorded when the trees were subjected to CPPU 10.0 mg·L−1 in 2021. There were no significant differences among all treatments regarding TA values. Moreover, there were no significant differences in TSS/TA among all treatments.
3.4. Tissue Section
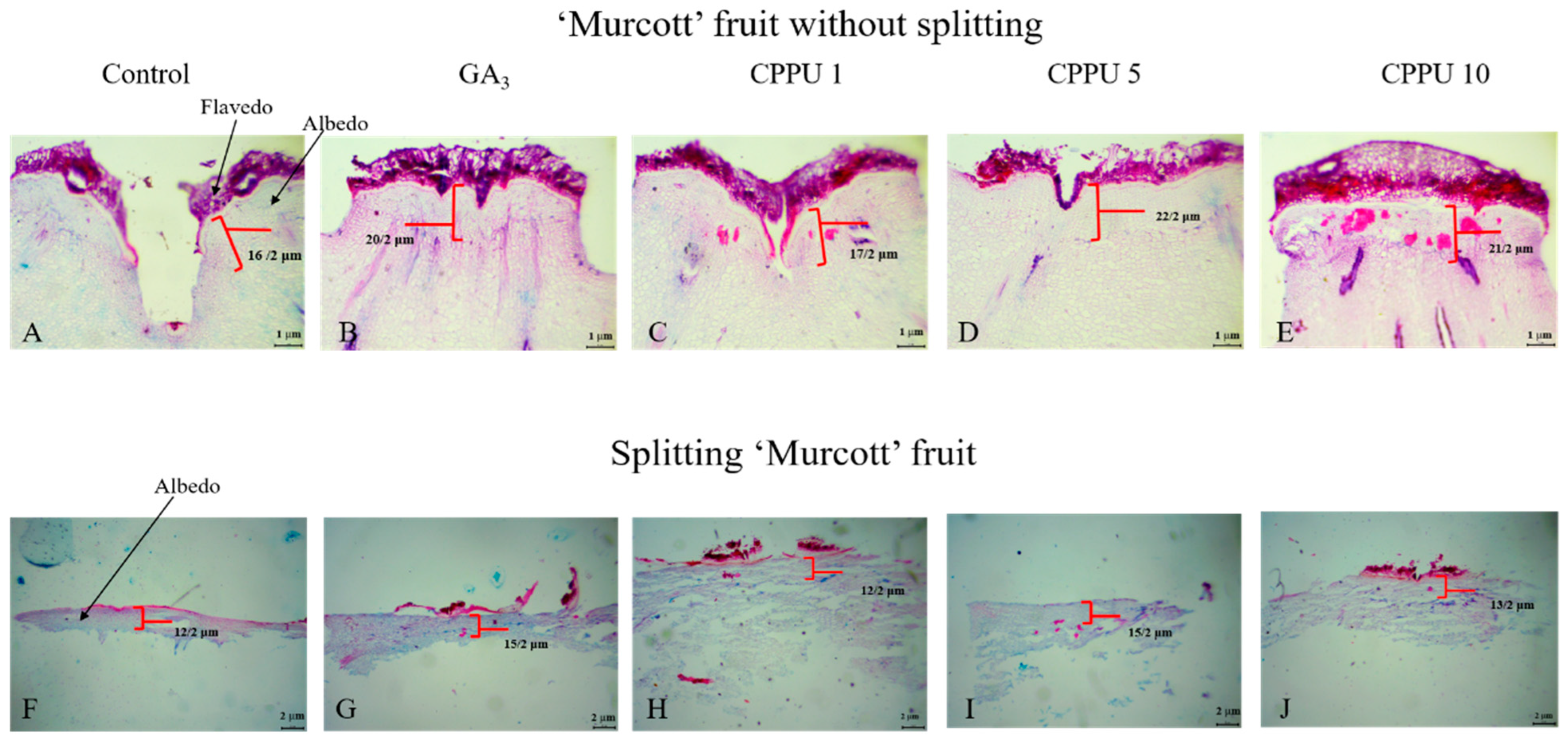
Regardless of fruit splitting, GA3- and CPPU-treated fruit resulted in a greater number of cellular layers in the exocarp compared with untreated fruit. The fruit without splitting exhibited 16, 20, 17, 22, and 21 layers per 2 μm in control, GA3 (20 mg·L−1), CPPU 1.0, 5.0, and 10.0 mg·L−1 fruit, respectively (Figure 3A–E). On the other hand, the split fruit showed fewer cellular layers in the exocarp, which were 12, 15, 12, 15, and 15 per 2 μm in control, GA3 (20 mg·L−1), CPPU 1.0, 5.0, and 10.0 mg·L−1 fruit, respectively (Figure 3F–J).

Figure 3.
The number of cell layers of the albedo on the stylar end of ‘Murcott’ fruit. A schematic representation of ‘Murcott’ fruit pericarp in a transverse section. Scale bars: 1 µm (A–E) and 2 µm (F–J).
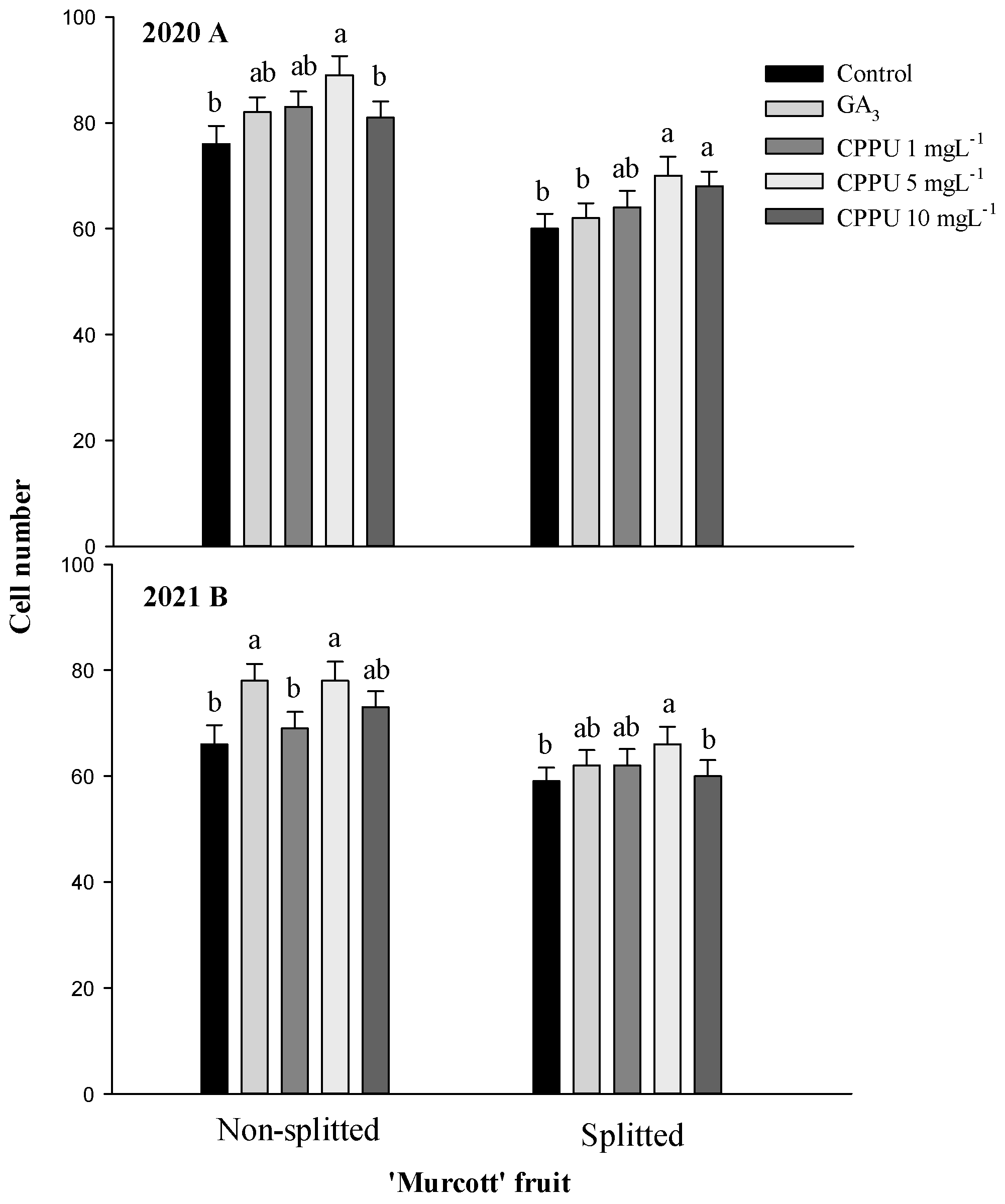
In addition, the non-split fruit had greater cell numbers in the exocarp than the split fruit in both years (Figure 4). ‘Murcott’ trees subjected to CPPU 5.0 mg·L−1 had significantly greater cell numbers in the exocarp compared to the control trees with and without fruit splitting.

Figure 4.
The effect of CPPU and GA3 treatments on cell numbers in the exocarp of split and non-split ‘Murcott’ fruit. The bars represent the means of four replications ± S.E. (n = 4). Means with the same letter are not significantly different according to the LSD test at 5%.
4. Discussion
The application of gibberellins (GA3) and forchlorfenuron (CPPU) significantly reduced the ratio of ‘Murcott’ fruit splitting in this study, and this finding is similar to those found in citrus [2,6,8,12,15,24], sweet cherry [25], pitaya fruit [20], grapes [26,27], and pomegranates [28]. The maximum peak of fruit splitting occurred in all treatments in October in both years, which might be correlated with rainfall patterns during the growing season in Taiwan.
GA3 and CPPU are PGRs responsible for cell expansion and division, leading to increased fruit size, such as in sweet cherries [25], grapes [26,27], and blueberries [29]. Although the GA3 application did not significantly influence ‘Murcott’ fruit size, such as fresh weight and fruit width, among all treatments in both 2020 and 2021, the trees treated with GA3 resulted in the highest fruit length among all treatments in 2021. An interfruit competition directly influenced fruit size, which has been shown in ‘Ellendale’ mandarin [8]. On the other hand, the trees subjected to CPPU also resulted in an increased fruit size in our study in 2020 and 2021. In particular, 5.0 mg·L−1 CPPU significantly influenced the fruit size and fresh weight of ‘Murcott’ fruit, which supports previous studies in kiwifruit [30,31], grapes [27], and blueberries [29]. However, there was no significant difference in fruit weight observed between CPPU-treated and non-treated litchi [32,33].
Previous studies showed that GA3 did not affect the content of TSS or TA in ‘Midknight Valencia’ orange [17], which is similar to our findings in this study. In addition, no significant difference in TSS or TTA values of fruit juice was observed in CPPU-treated fruit, as compared to the non-treated control fruit, when observing litchi fruit [32,33]. The opposite results were reported for the exogenous application of CPPU, which resulted in an increase in carbohydrates observed in tomato fruit [34], kiwifruit [35], and macadamia fruit [36]. A possible explanation may be that CPPU causes significant changes in the cell layer and cell number in the peel instead of in the segment, leading to the percentage of carbohydrates constantly accumulating in the pulp or juice sac. A similar finding was reported in CPPU-treated litchi fruit [32], which may support the above rationale. However, this hypothesis requires further investigation.
In addition, GA3 did not significantly affect peel strength or peel thickness, but sprayed 5.0 mg·L−1 CPPU effectively increased fruit peel strength and peel thickness in this study. Although CPPU functions like cytokinin in increasing fruit size, it is effective in enhancing peel thickness and fruit firmness, as reported in kiwifruit [37], grapes [26], litchi [32,33], and pitaya fruit [20].
Antognozzi et al. [35] indicated that CPPU stimulates cell division and elongation in ‘Hayward’ kiwifruit due to an increase in cell number and size in the outer and inner pericarp. Thereafter, Zoffoli et al. [26] reported that CPPU effectively increases pedicel thickness and cuticle content in table grapes. In addition, Fahima et al. [32] reported that CPPU application can increase pericarp cell division and cell number in brachysclereids, leading to a thicker pericarp and therefore reducing the susceptibility to micro-crack formation in litchi fruit. Furthermore, CPPU application resulted in more cellular layers and greater cell numbers in the exocarp whether ‘Murcott’ fruit was split or not, which supports a finding that CPPU-treated kiwifruit ‘Hongyang’ had more subepidermal cells and cell layers [38].
Previous studies indicated that CPPU application resulted in the regulation of endogenous phytohormone synthesis such as auxin, gibberellins, and cytokinin which controls fruit development [39,40]. A similar finding was reported in kiwifruit due to CPPU-induced cytokinin and gibberellin biosynthesis [38]. Furthermore, Wu et al. [38] indicated that CPPU-treated fruit exhibited a significant increase in gene expression which encodes key GA and CK biosynthetic enzymes in kiwifruit, and similar results were found in the regulation of fruit development in tomato [41,42].
5. Conclusions
It is obvious that the application of CPPU after full bloom not only increases the fruit size but also the peel thickness and strength of ‘Murcott’ fruit. The susceptibility of citrus to splitting is most likely because of the thin and weak peel at the stylar or navel end. Thus, how to increase the greater cell numbers and layers of exocarp may play an important role in reducing the splitting occurrence of ‘Murcott’ fruit. Further research is thus required to evaluate the concentration and frequency of CPPU application and the responses of other split-susceptible citrus species.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae10101023/s1, Figure S1: The effect of CPPU and GA3 treatments on cell numbers in the exocarp. (A) Control, (B) GA3, (C) 1.0 mg L−1 CPPU, (D) 5.0 mg L−1 CPPU, and (E) 10.0 mg L−1 CPPU.
Author Contributions
Methodology, P.-T.C.; Validation, Y.-L.J.; Investigation, Y.-L.J.; Writing—original draft, P.-T.C.; Writing—review & editing, P.-T.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Council of Agriculture Executive Yuan, Taiwan, Grant No. 109AS-7.3.4-FD-Z1, and the APC was funded by Grant No. 109AS-7.3.4-FD-Z1.
Data Availability Statement
The original contributions presented in the study are included in the Supplementary Materials, further inquiries can be directed to the corresponding author.
Conflicts of Interest
On behalf of all authors, the corresponding author states that there are no conflicts of interest.
References
- Rabe, E.; Van der Walt, W.; Kleynhans, S. Factors Influencing Fruit Splitting in Ellendale; Annual Report S.A. Co-Op. Citrus Exchange Ltd.: Nelspruit, South Africa, 1989. [Google Scholar]
- Mupambi, G. Studies to Reduce the Size of the Navel-End Opening of Navel Oranges. Ph.D. Dissertation, University Stellenbosch, Stellenbosch, South Africa, 2010. [Google Scholar]
- De Cicco, V.; Intrigliolo, F.; Ippolito, A.; Vanadia, S.; Guiffrida, A. Factors in Navelina orange splitting. Proc. Int. Soc. Citricult. 1988, 1, 535–540. [Google Scholar]
- Bower, J.P.; Gilfillan, I.M.; Skinner, H. Fruit splitting in ‘Valencia’ and its relationship to the pectin status of the rind. Proc. Int. Soc. Citricult. 1992, 1, 511–514. [Google Scholar]
- Goldschmidt, E.E.; Galili, D. Fruit splitting in ‘Murcott’ tangerines: Control by reduced water supply. Proc. Int. Soc. Citricult. 1992, 2, 657–660. [Google Scholar]
- Almela, V.; Zaragoza, S.; Primo-Millo, E.; Agusti, M. Hormonal control of splitting in ‘Nova’ mandarin fruit. J. Hort. Sci. 1994, 69, 969–973. [Google Scholar]
- Sdoodee, S.; Chiarawipa, R. Fruit splitting occurrence of Shogun mandarin (Citrus reticulata Blanco cv. Shogun) in southern Thailand and alleviation by calcium and boron sprays. Songklanakarin J. Sci. Technol. 2005, 27, 719–730. [Google Scholar]
- Rabe, E.; Van Rensburg, P.J.J.; Van Der Walt, H.; Bower, J. Factors influencing preharvest fruit splitting in Ellendale (C. reticulata). HortScience 1990, 25, 1135f. [Google Scholar] [CrossRef]
- Barry, G.H.; Bower, J.P. Manipulation of fruit set and stylar-end fruit split in ‘Nova’ mandarin hybrid. Sci. Hortic. 1997, 70, 243–250. [Google Scholar] [CrossRef]
- García-Luis, A.; Duarte, A.M.M.; Porras, I.; García-Lidon, A.; Guardiola, J.L. Fruit splitting in ‘Nova’ hybrid mandarin in relation to the anatomy of the fruit and fruit set treatments. Sci. Hortic. 1994, 57, 215–231. [Google Scholar] [CrossRef]
- Rabe, E.; Van Rensburg, P.J.J. Gibberellic acid sprays, girdling, flower thinning and potassium applications affect fruit splitting and yield in the ‘Ellendale’ tangor. J. Hortic. Sci. 1996, 71, 195–203. [Google Scholar] [CrossRef]
- García-Luis, A.; Duarte, A.M.M.; Kanduser, M.; Guardiola, J.L. The anatomy of fruit in relation to the propensity of citrus fruit to split. Sci. Hortic. 2001, 87, 33–52. [Google Scholar] [CrossRef]
- Coit, J. Citrus Fruits; Macmillan: New York, NY, USA, 1915; p. 520. [Google Scholar]
- Greenberg, J.; Kaplan, I.; Fainzack, M.; Egozi, Y.; Giladi, B. Effects of auxin sprays on yield, fruit size, splitting and the incidence of creasing of ‘Nova’ mandarin. Acta Hortic. 2006, 727, 249–254. [Google Scholar] [CrossRef]
- Sandhu, S.; Bal, J.S. Quality Improvement in Lemon (Citrus Limon (L.) Burm.) through Integrated Management of Fruit Cracking. Afr. J. Agric. Res. 2013, 8, 3552–3557. [Google Scholar]
- Erner, Y.; Goren, R.; Monselise, S.P. Reduction of peel roughness of ‘Shamouti’ orange with growth regulators. J. Am. Soc. Hortic. Sci. 1976, 101, 513–515. [Google Scholar] [CrossRef]
- Stander, O.P.J.; Theron, K.I.; Cronjé, P.J.R. Foliar 2,4-D Application after physiological fruit drop reduces fruit splitting of Mandarin. HortTechnology 2014, 24, 717–723. [Google Scholar] [CrossRef]
- Kopečný, D.; Briozzo, P.; Popelková, H.; Šebela, M.; Končitíková, R.; Spíchal, L.; Nisler, J.; Madzak, C.; Frébort, I.; Laloue, M. Phenyl-and benzylurea cytokinins as competitive inhibitors of cytokinin oxidase/dehydrogenase: A structural study. Biochimie 2010, 92, 1052–1062. [Google Scholar] [CrossRef]
- Cruz-Castillo, J.G.; Baaldicchi, A.; Frioni, T.; Marocchi, F.; Moscatello, S.; Proietti, S.; Battistelli, A.; Famiani, F. Pre-anthesis CPPU low dosage application increases ‘Hayward’ kiwifruit weight without affecting the other qualitative and nutritional characteristics. Food Chem. 2014, 158, 224–228. [Google Scholar] [CrossRef]
- Jiang, Y.L.; Chen, L.Y.; Lee, T.C.; Chang, P.T. Improving postharvest storage of Fresh red-fleshed pitaya (Hylocereus polyrhizus sp.) fruit by pre-harvest application of CPPU. Sci. Hortic. 2020, 273, 109646. [Google Scholar] [CrossRef]
- Khaimov, A.; Mizrahi, Y. Effects of day-length, radiation, flower thinning and growth regulators on flowering of the vine cacti Hylocereus undatus and Selenicereus megalanthus. J. Hortic. Sci. Biotechnol. 2006, 60, 371–383. [Google Scholar] [CrossRef]
- Chang, P.T.; Hsieh, C.C.; Jiang, Y.L. Responses of ‘Shih Huo Chuan’ pitaya (Hylocereus polyrhizus (Weber) Britt. & Rose) to different degrees of shading nets. Sci. Hortic. 2016, 198, 154–162. [Google Scholar]
- Atkinson, J.A.; Wells, D.M. An updated protocol for high throughput plant tissue sectioning. Front. Plant Sci. 2017, 8, 1721. [Google Scholar] [CrossRef]
- Greenberg, J.; Holtzman, S.; Fainzack, M.; Egozi, Y.; Giladi, B.; Oren, Y.; Kaplan, I. Effects of NAA and GA3 sprays on fruit size and the incidence of creasing of ‘Washington’ navel orange. Acta Hortic. 2010, 884, 273–279. [Google Scholar] [CrossRef]
- Cline, J.A.; Trought, M. Effect of gibberellic acid on fruit cracking and quality of Bing and Sam sweet cherries. Can. J. Plant Sci. 2007, 87, 545–550. [Google Scholar] [CrossRef]
- Zoffoli, J.P.; Latorre, B.A.; Naranjo, P. Preharvest applications of growth regulators and their effect on postharvest quality of table grapes during cold storage. Postharvest Biol. Technol. 2009, 51, 183–192. [Google Scholar] [CrossRef]
- Marzouk, H.A.; Kassem, H.A. Improving yield, quality, and shelf life of Thompson seedless grapevine by preharvest foliar applications. Sci. Hortic. 2011, 130, 425–430. [Google Scholar] [CrossRef]
- Sahu, P.S.A.; Sahu, A. Effect of pre-harvest sprays of forchlorfenuron and boron on fruit cracking and quality of pomegranate (Punica granatum L.) cv. Kandhari. Int. J. Chem. Stud. 2018, 6, 2998–3002. [Google Scholar]
- Retamales, J.B.; Lobos, G.A.; Romero, S.; Godoy, R.; Moggia, C. Repeated applications of CPPU on highbush blueberry cv. Duke increase yield and enhance fruit quality at harvest and during postharvest. Chil. J. Agri. Res. 2014, 74, 157–161. [Google Scholar] [CrossRef]
- Cruz-Castillo, J.G.; Woolley, D.; Lawes, G. Kiwifruit size and CPPU response are influenced by the time of anthesis. Sci. Hortic. 2002, 95, 23–30. [Google Scholar] [CrossRef]
- Kim, J.; Takami, Y.; Mizugami, T.; Beppu, K.; Fukuda, T.; Kataoka, I. CPPU application on size and quality of hardy kiwifruit. Sci. Hortic. 2006, 110, 219–222. [Google Scholar] [CrossRef]
- Fahima, A.; Levinkron, S.; Maytal, Y.; Hugger, A.; Lax, I.; Huang, X.; Eyal, Y.; Lichter, A.; Goren, M.; Stern, R.A.; et al. Cytokinin treatment modifies litchi fruit pericarp anatomy leading to reduced susceptibility to post-harvest pericarp browning. Plant Sci. 2019, 283, 41–50. [Google Scholar] [CrossRef]
- Liu, X.S.; Luo, Y.C.; Wang, H.C.; Huang, X.M. Post-bloom CPPU application is effective at improving fruit set and suppressing coloration but ineffective at increasing fruit size in litchi. Horticulturae 2022, 8, 1096. [Google Scholar] [CrossRef]
- Albacete, A.; Cantero-Navarro, E.; Balibrea, M.E.; Grosskinsky, D.K.; De la Cruz Gonzalez, M.; Martinez-Adujar, C.; Smigocki, A.C.; Roitsch, T.; Perez-Alfocea, F. Hormonal and metabolic regulation of tomato fruit sink activity and yield under salinity. J. Exp. Bot. 2014, 65, 6081–6095. [Google Scholar] [CrossRef] [PubMed]
- Antognozzi, E.; Battistelli, A.; Famiani, F.; Moscatello, S.; Stanica, F.; Tombesi, A. Influence of CPPU on carbohydrate accumulation and metabolism in fruits of Actinidia deliciosa (A. Chev.). Sci. Hortic. 1996, 65, 37–47. [Google Scholar] [CrossRef]
- Zeng, H.; Yang, W.; Lu, C.; Lin, W.; Zou, M.; Zhang, H.; Wan, J.; Huang, X. Effect of CPPU on carbohydrate and endogenous hormone levels in young macadamia fruit. PLoS ONE 2016, 11, e0158705. [Google Scholar] [CrossRef] [PubMed]
- Patterson, K.; Mason, K.; Gould, K. Effects of CPPU (N-(2-chloro-4-pyridyl)-N′-phenylurea) on fruit growth, maturity, and storage quality of kiwifruit. N. Z. J. Crop Hortic. Sci. 1993, 21, 253–261. [Google Scholar] [CrossRef]
- Wu, L.; Lan, J.; Xiang, X.; Jin, Z.; Khan, S.; Liu, Y. Transcriptome sequencing and endogenous phytohormone analysis reveal new insights in CPPU controlling fruit development in kiwifruit (Actinidia chinensis). PLoS ONE 2020, 15, e0240355. [Google Scholar] [CrossRef]
- Devoghalaere, F.; Doucen, T.; Guitton, B.; Keeling, J.; Payne, W.; Ling, T.J. A genomics approach to understanding the role of auxin in apple (Malus × domestica) fruit size control. BMC Plant Biol. 2012, 12, 7. [Google Scholar] [CrossRef]
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of plant hormones and their interplay in development and ripening of fleshy fruits. J. Exp. Bot. 2014, 65, 4561–4575. [Google Scholar] [CrossRef]
- Martí, C.; Orzáez, D.; Ellul, P.; Moreno, V.; Carbonell, J.; Granell, A. Silencing of DELLA induces facultative parthenocarpy in tomato fruits. Plant J. 2007, 52, 865–876. [Google Scholar] [CrossRef]
- Mignolli, F.; Vidoz, M.L.; Picciarelli, P.; Mariotti, L. Gibberellins modulate auxin responses during tomato (Solanum lycopersicum L.) fruit development. Physiol. Plant. 2019, 165, 768–779. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).