
Genetic Diversity and Population Structure Analysis of Excellent Sugar Beet (Beta vulgaris L.) Germplasm Resources


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Phenotypic Trait Measurements
2.3. DNA Extraction and Genotyping
2.4. Data Analysis
3. Results
3.1. Genetic Diversity Using SSR Primers
3.2. Genetic Diversity Using InDel Primers
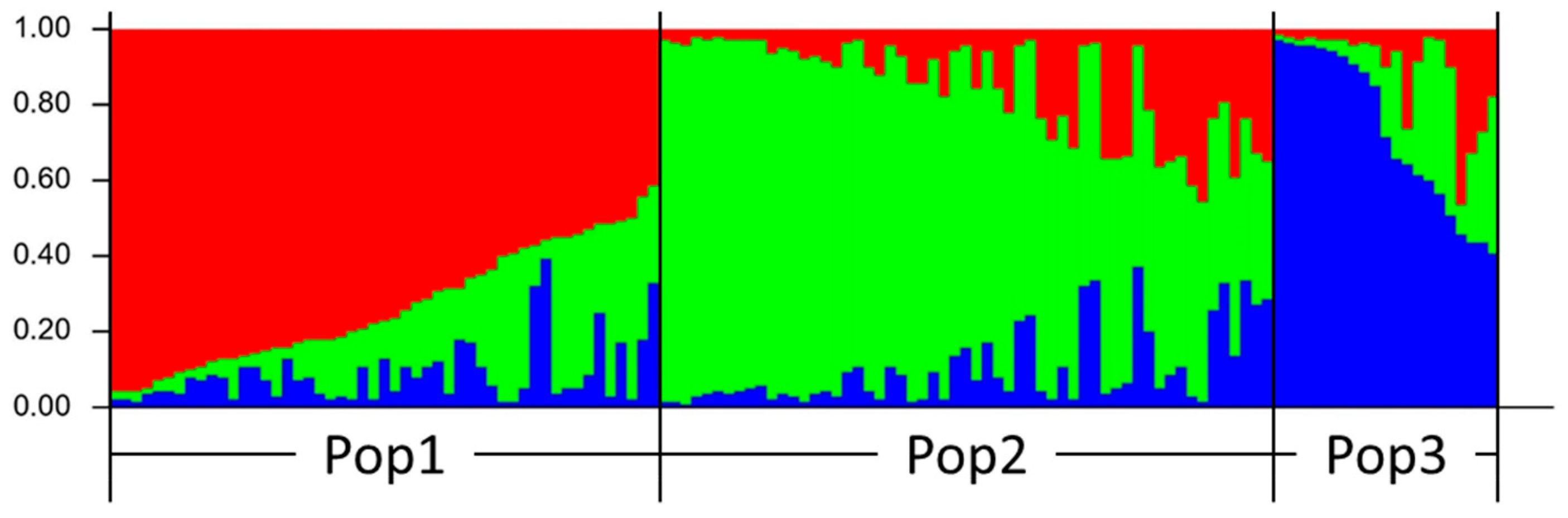
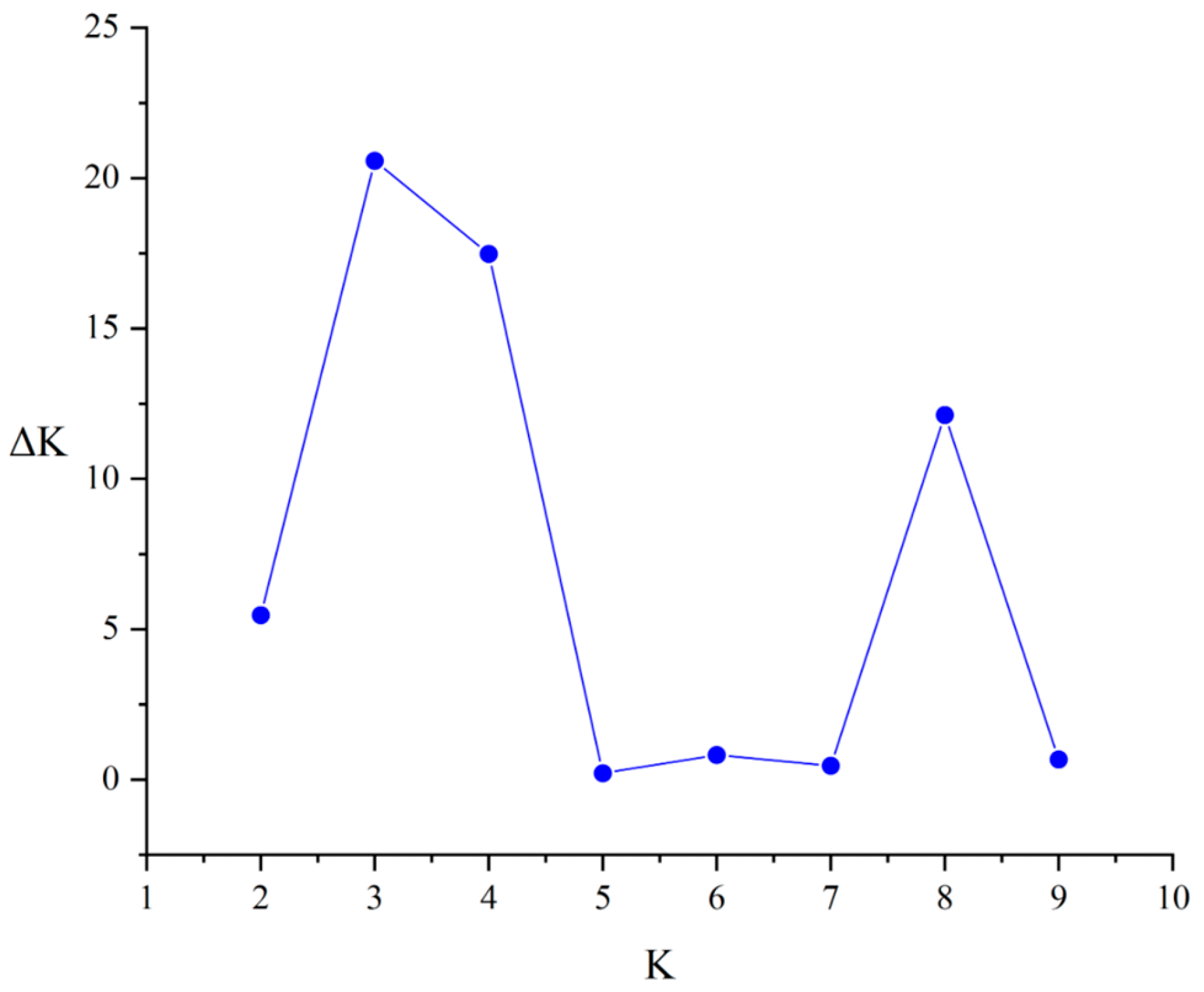
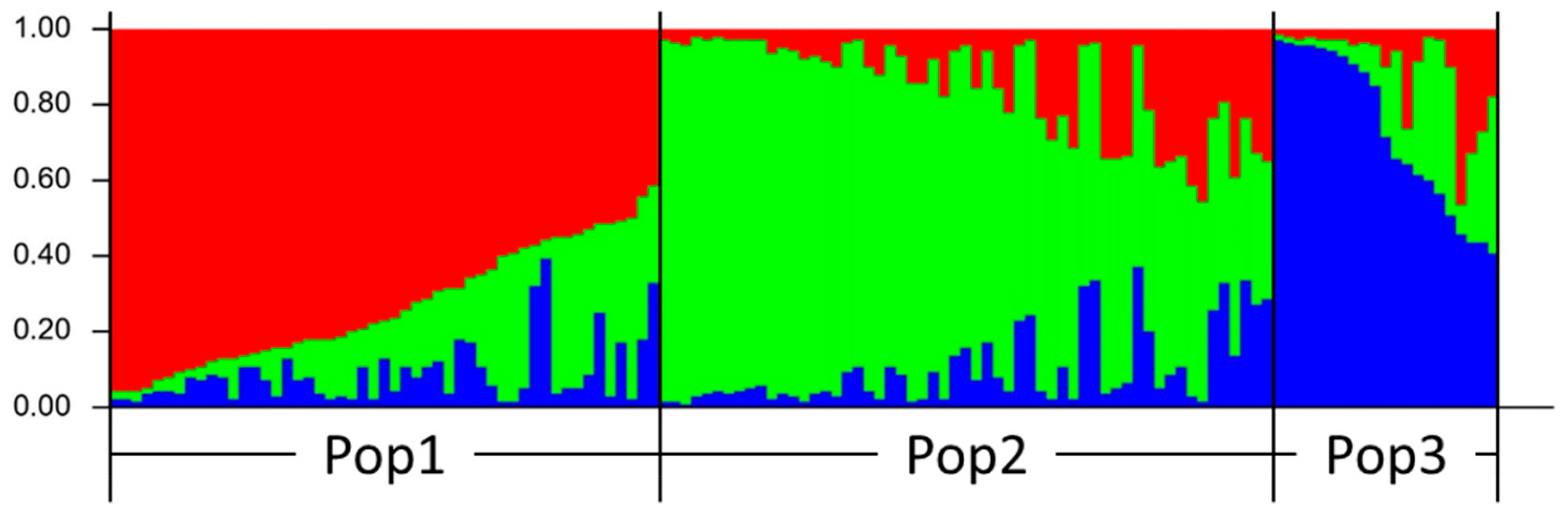
3.3. Population Structure Analysis
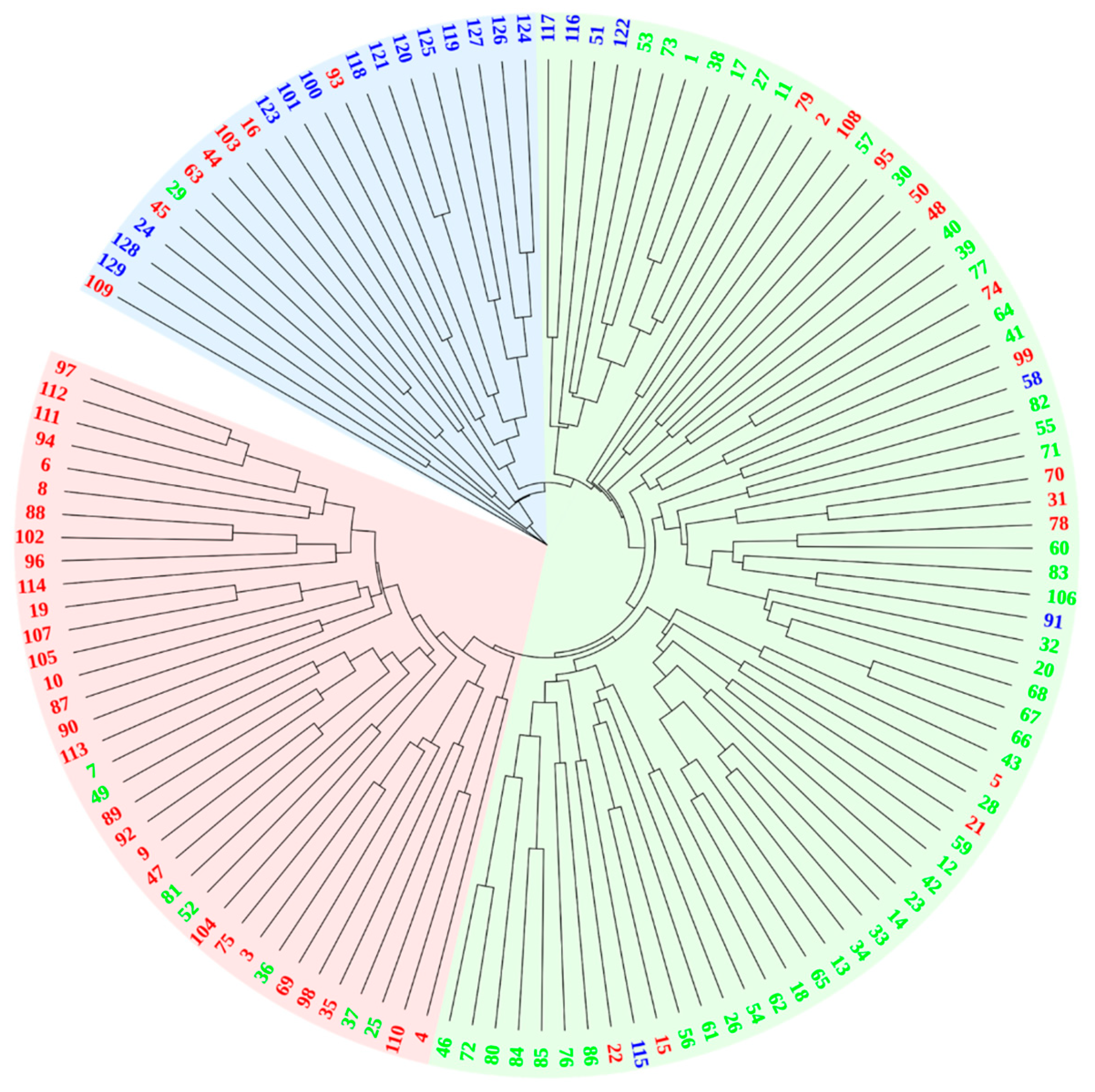
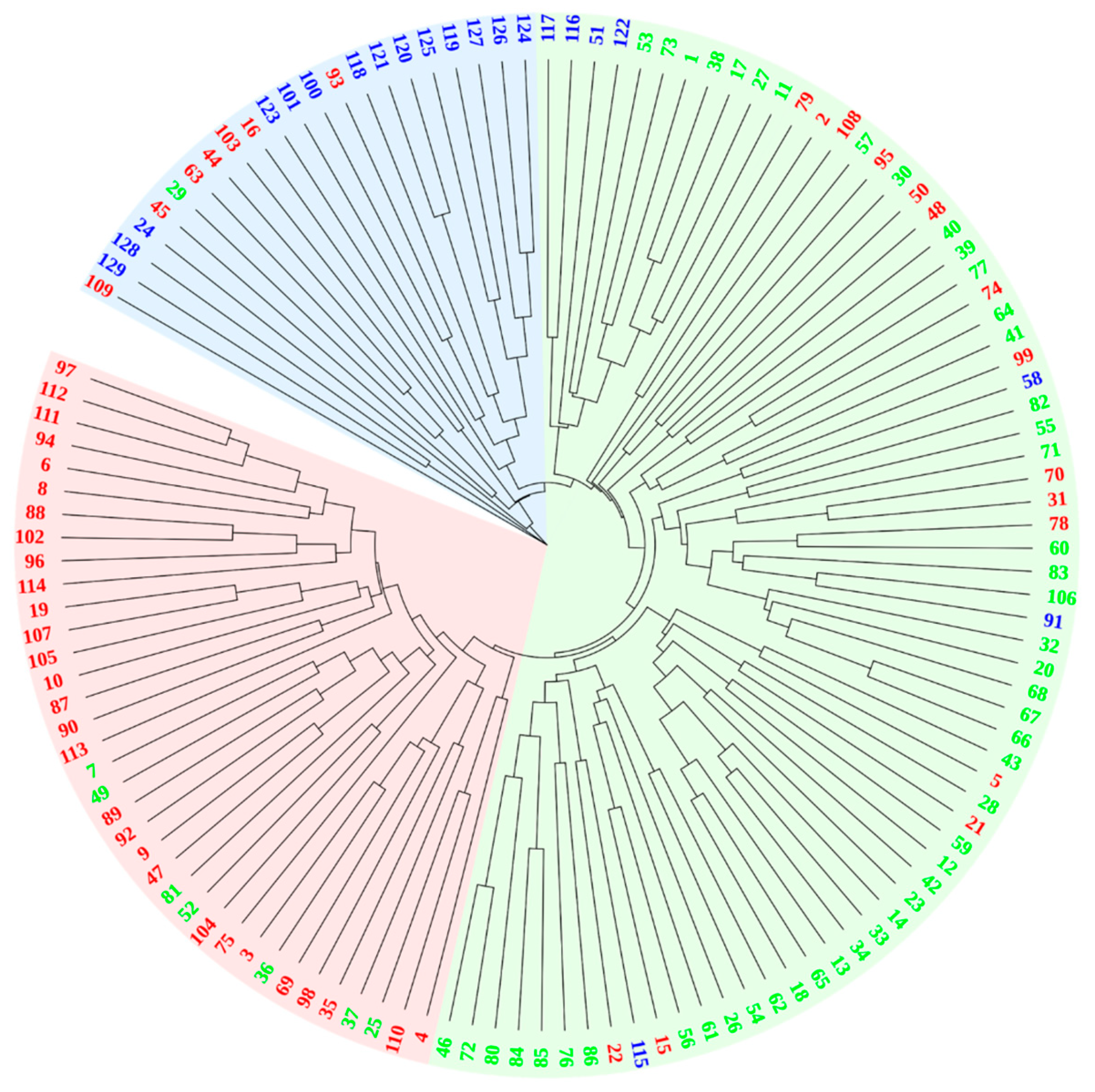
3.4. Genetic Distance and Cluster Structure
3.5. Variation Analysis of the Phenotypic Traits
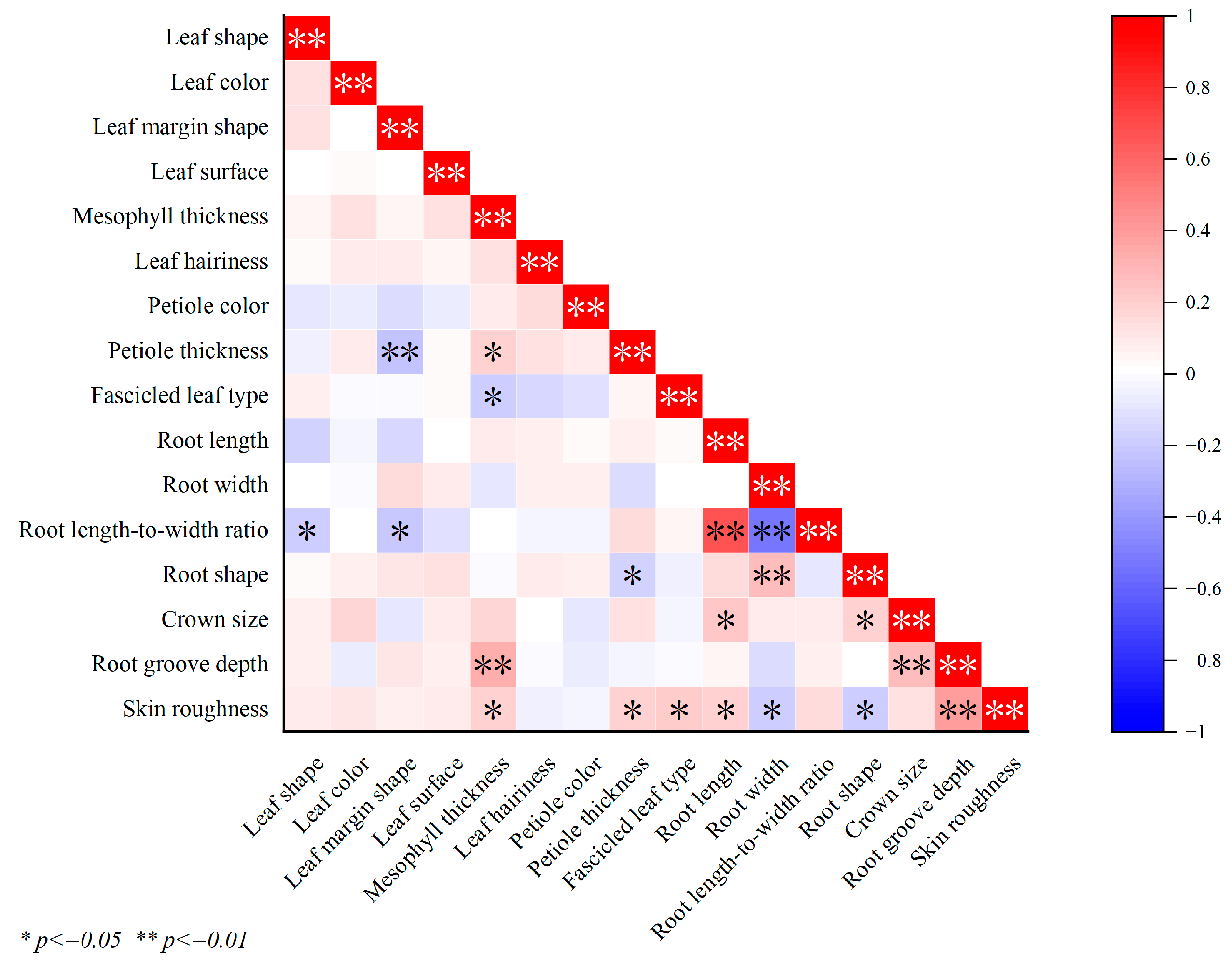
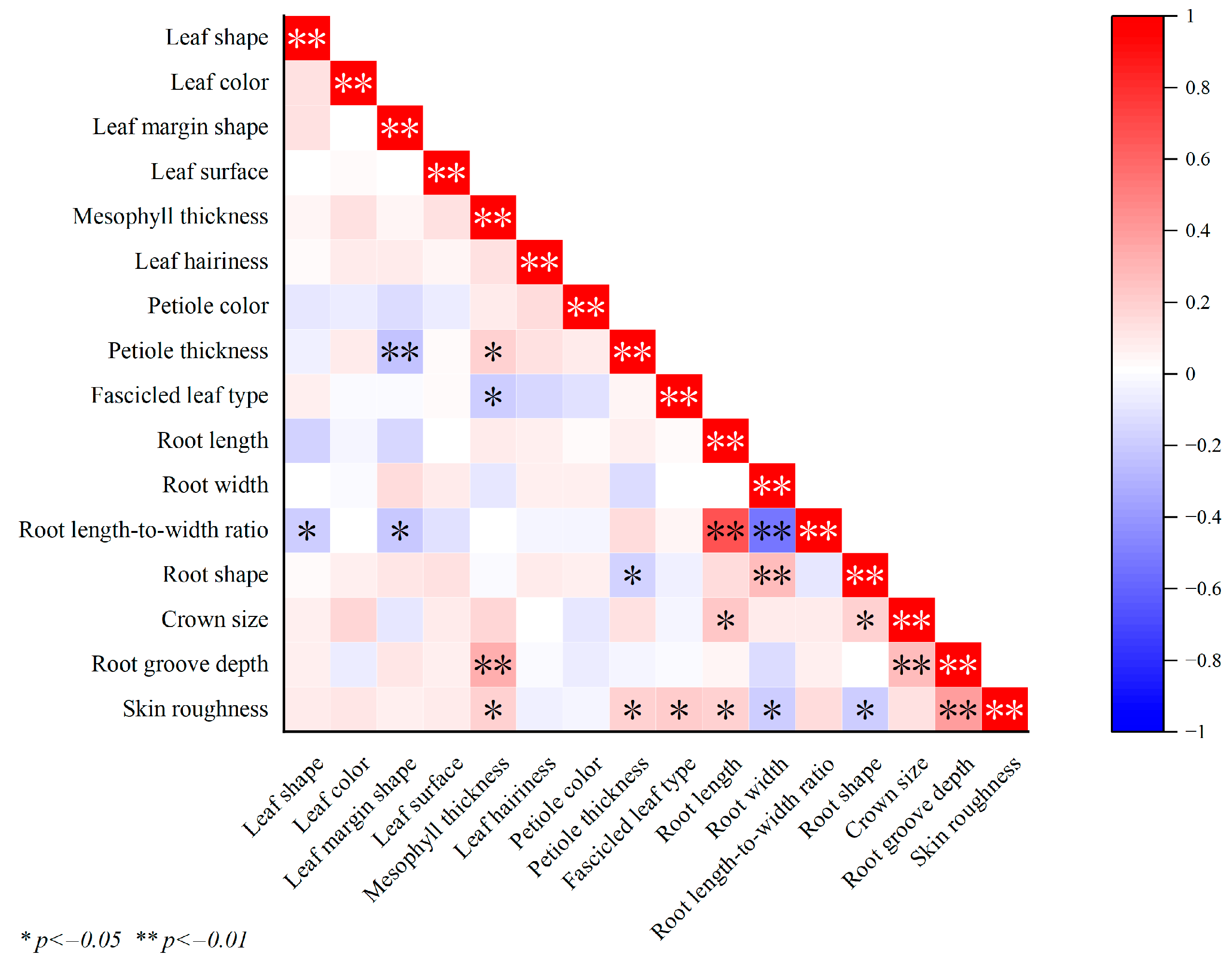
3.6. Correlation Analysis
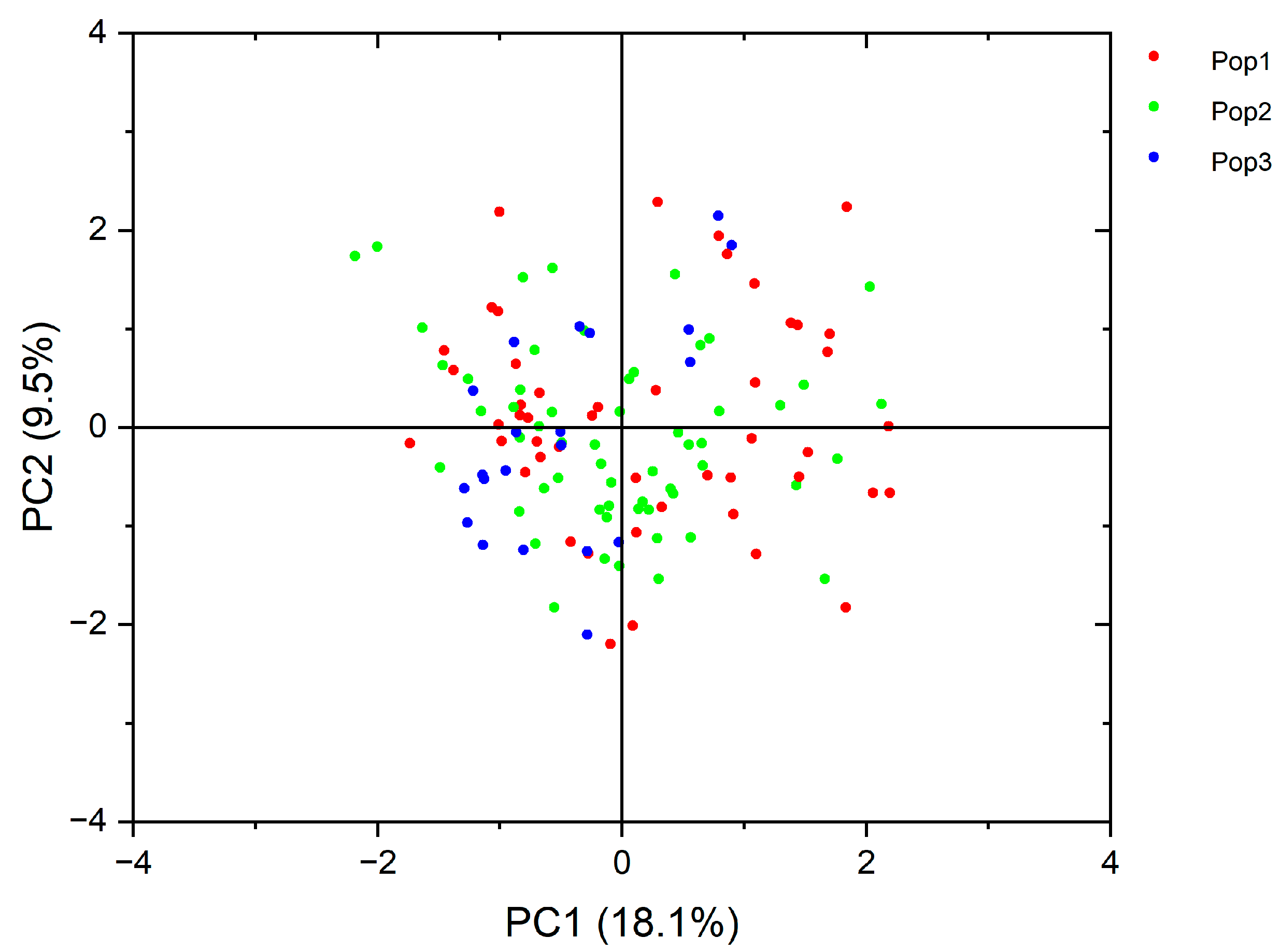
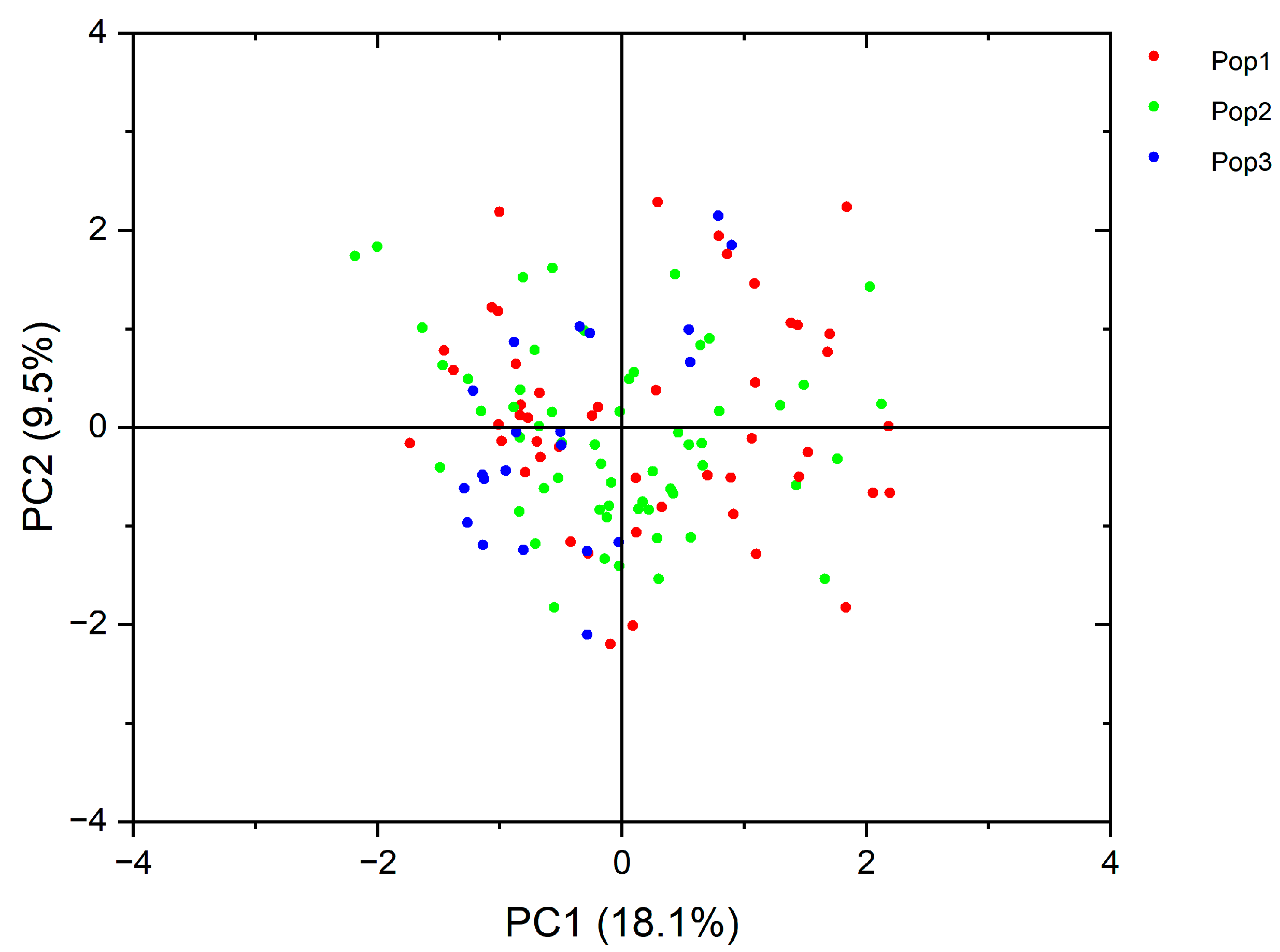
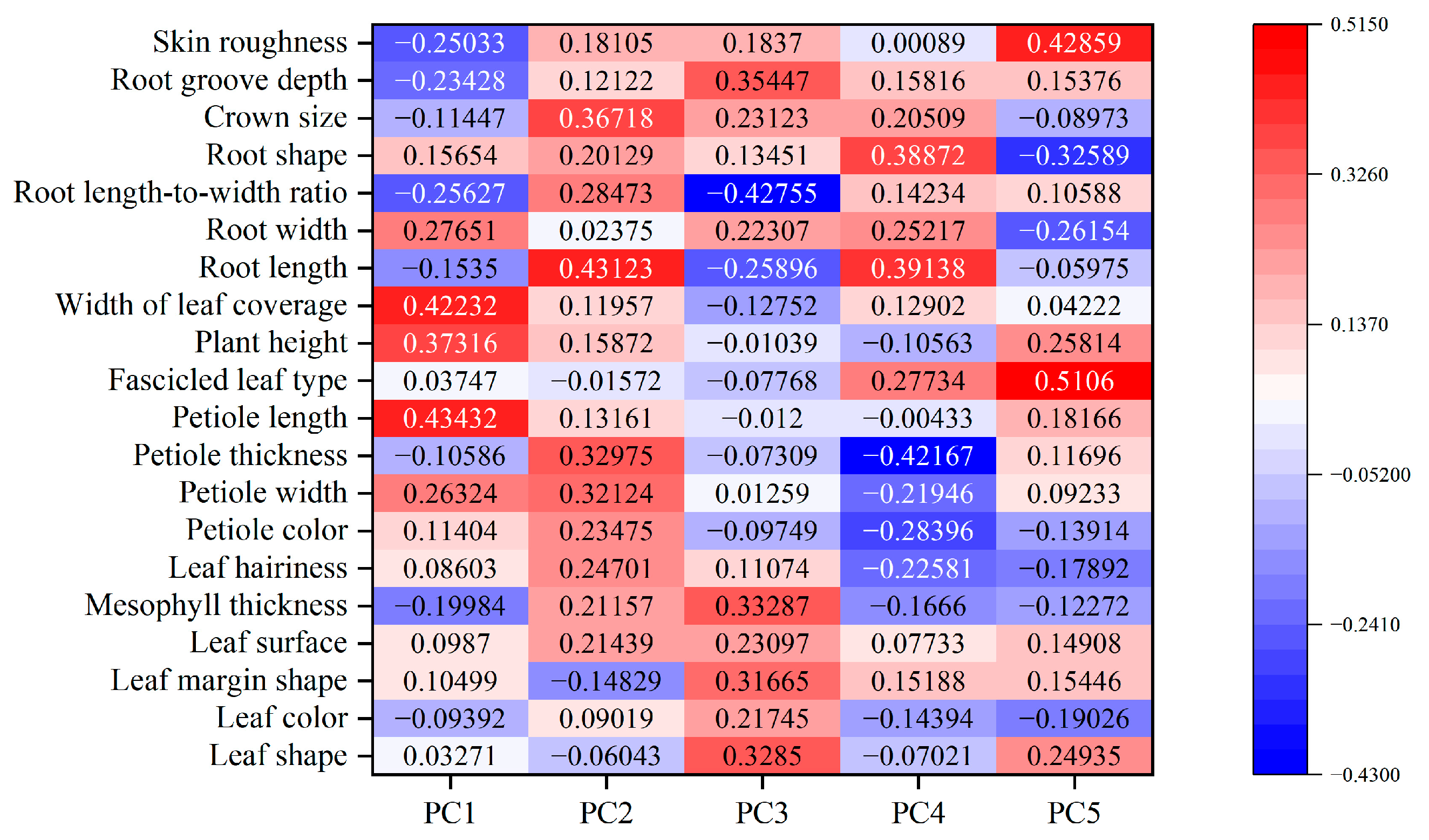
3.7. PCA Evaluation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wascher, F.L.; Stralis-Pavese, N.; McGrath, J.M.; Schulz, B.; Himmelbauer, H.; Dohm, J.C. Genomic distances reveal relationships of wild and cultivated beets. Nat. Commun. 2022, 13, 2021. [Google Scholar] [CrossRef] [PubMed]
- Wolfgang, A.; Temme, N.; Tilcher, R.; Berg, G. Understanding the sugar beet holobiont for sustainable agriculture. Front. Microbiol. 2023, 14, 1151052. [Google Scholar] [CrossRef] [PubMed]
- Stevanato, P.; Chiodi, C.; Broccanello, C.; Concheri, G.; Biancardi, E.; Pavli, O.; Skaracis, G.N. Sustainability of the Sugar Beet Crop. Sugar Tech 2019, 21, 703–716. [Google Scholar] [CrossRef]
- Duraisam, R.; Salelgn, K.; Berekete, A.K. Production of beet sugar and bio-ethanol from sugar beet and it bagasse: A review. Int. J. Eng. Trends Technol. 2017, 43, 222–233. [Google Scholar] [CrossRef]
- McGrath, J.M. Assisted Breeding in Sugar Beets. Sugar Tech 2010, 12, 187–193. [Google Scholar] [CrossRef]
- Li, J.; Lühmann, A.K.; Weißleder, K.; Stich, B. Genome-wide distribution of genetic diversity and linkage disequilibrium in elite sugar beet germplasm. BMC Genom. 2011, 12, 484. [Google Scholar] [CrossRef] [PubMed]
- Ćurčić, Ž.; Taški-Ajduković, K.; Nagl, N. Relationship between hybrid performance and genetic variation in self-fertile and self-sterile sugar beet pollinators as estimated by SSR markers. Euphytica 2017, 213, 108. [Google Scholar] [CrossRef]
- McGrath, J.M.; Panella, L.W. Sugar Beet Breeding. Plant Breed. Rev. 2018, 42, 167–218. [Google Scholar] [CrossRef]
- Monteiro, F.; Frese, L.; Castro, S.; Duarte, M.C.; Paulo, O.S.; Loureiro, J.; Romeiras, M.M. Genetic and Genomic Tools to Asssist Sugar Beet Improvement: The Value of the Crop Wild Relatives. Front. Plant Sci. 2018, 9, 74. [Google Scholar] [CrossRef]
- Veloso, M.M.; Simões-Costa, M.C.; Guimarães, J.B.; Ribeiro, C.M.; Evaristo, I.; Espírito-Santo, D.; Pinto-Ricardo, C.; Paulo, O.S.; Duarte, M.C. Genetic Diversity and Population Structure of Wild Beets (Beta spp.) from the Western Iberian Peninsula and the Azores and Madeira Islands. Diversity 2021, 13, 593. [Google Scholar] [CrossRef]
- Fugate, K.K.; Campbell, L.G.; Covarrubias-Pazaran, G.; Rodríguez-Bonilla, L.; Zalapa, J. Genetic diversity is enhanced in Wild × Cultivated hybrids of sugarbeet (Beta vulgaris L.) despite multiple selection cycles for cultivated traits. Genet. Resour. Crop Evol. 2021, 68, 2549–2563. [Google Scholar] [CrossRef]
- Ramanatha Rao, V.; Hodgkin, T. Genetic diversity and conservation and utilization of plant genetic resources. Plant Cell Tissue Organ Cult. 2002, 68, 1–19. [Google Scholar] [CrossRef]
- Guo, S.; Ji, P.; Wang, J.; He, Y.; Zhang, Y.; Zhang, F.; Yun, Y.; Zhang, G. Estimation of Genetic Diversity between and within Biparental Clones and Full-Sib Families of the Chinese Pine Using SSR Markers. Horticulturae 2023, 9, 1205. [Google Scholar] [CrossRef]
- Lee, H.Y.; Ro, N.Y.; Jeong, H.J.; Kwon, J.K.; Jo, J.; Ha, Y.; Jung, A.; Han, J.W.; Venkatesh, J.; Kang, B.C. Genetic diversity and population structure analysis to construct a core collection from a large Capsicum germplasm. BMC Genet. 2016, 17, 142. [Google Scholar] [CrossRef]
- Mai, T.T.P.; Hardner, C.M.; Alam, M.M.; Henry, R.J.; Topp, B.L. Phenotypic characterisation for growth and nut characteristics revealed the extent of genetic diversity in wild macadamia germplasm. Agriculture 2021, 11, 680. [Google Scholar] [CrossRef]
- Placide, R.; Shimelis, H.; Laing, M.; Gahakwa, D. Application of principal component analysis to yield and yield related traits to identify sweet potato breeding parents. J. Trop. Agric. 2015, 92, 1–15. Available online: https://www.researchgate.net/publication/274958783 (accessed on 10 September 2023).
- Liu, D.; Wang, X.; Li, W.; Li, J.; Tan, W.; Xing, W. Genetic Diversity Analysis of the Phenotypic Traits of 215 Sugar Beet Germplasm Resources. Sugar Tech 2022, 24, 1790–1800. [Google Scholar] [CrossRef]
- Liu, S.; Zhong, H.; Zhang, F.; Wang, X.; Wu, X.; Wang, J.; Shi, W. Genetic Diversity and Core Germplasm Research of 144 Munake Grape Resources Using 22 Pairs of SSR Markers. Horticulturae 2023, 9, 917. [Google Scholar] [CrossRef]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genic microsatellite markers in plants: Features and applications. Trends Biotechnol. 2005, 23, 48–55. [Google Scholar] [CrossRef]
- Abu-Ellail, F.F.B.; Salem, K.F.M.; Saleh, M.M.; Alnaddaf, L.M.; Al-Khayri, J.M. Molecular Breeding Strategies of Beetroot (Beta vulgaris ssp. vulgaris var. conditiva Alefeld). Adv. Plant Breed. Strateg. Veg. Crops 2021, 8, 157–212. [Google Scholar] [CrossRef]
- Stevanato, P.; Broccanello, C.; Biscarini, F.; Del Corvo, M.; Sablok, G.; Panella, L.; Stella, A.; Concheri, G. High-throughput RAD-SNP genotyping for characterization of sugar beet genotypes. Plant Mol. Biol. Report. 2014, 32, 691–696. [Google Scholar] [CrossRef]
- Simko, I.; Eujayl, I.; van Hintum, T.J.L. Empirical evaluation of DArT, SNP, and SSR marker-systems for genotyping, clustering, and assigning sugar beet hybrid varieties into populations. Plant Sci. Int. J. Exp. Plant Biol. 2012, 184, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [Google Scholar] [CrossRef]
- Taški-Ajduković, K.; Nagl, N.; Ćurčić, Ž.; Zorić, M. Estimation of genetic diversity and relationship in sugar beet pollinators based on SSR markers. Electron. J. Biotechnol. 2017, 27, 1–7. [Google Scholar] [CrossRef]
- Smulders, M.J.M.; Esselink, G.D.; Everaert, I.G.; de Riek, J.; Vosman, B. Characterisation of sugar beet (Beta vulgaris L. ssp. vulgaris) varieties using microsatellite markers. BMC Genet. 2010, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Pathak, A.; Kumar, R.; Joshi, B. Genetic diversity of sugar beet genotypes evaluated by microsatellite DNA markers. J. Environ. Biol. 2017, 384, 777–783. [Google Scholar] [CrossRef]
- Li, J.; Schulz, B.; Stich, B. Population structure and genetic diversity in elite sugar beet germplasm investigated with SSR markers. Euphytica 2010, 175, 35–42. [Google Scholar] [CrossRef]
- Andrello, M.; Henry, K.; Devaux, P.; Verdelet, D.; Desprez, B.; Manel, S. Insights into the genetic relationships among plants of Beta section Beta using SNP markers. Theor. Appl. Genet. 2017, 130, 1857–1866. [Google Scholar] [CrossRef]
- Xiang, X.; Li, C.; Li, L.; Bian, Y.; Kwan, H.S.; Nong, W.; Cheung, M.K.; Xiao, Y. Genetic diversity and population structure of Chinese Lentinula edodes revealed by InDel and SSR markers. Mycol. Prog. 2016, 15, 37. [Google Scholar] [CrossRef]
- Wu, K.; Yang, M.; Liu, H.; Tao, Y.; Mei, J.; Zhao, Y. Genetic analysis and molecular characterization of Chinese sesame (Sesamum indicum L.) cultivars using Insertion-Deletion (InDel) and Simple Sequence Repeat (SSR) markers. BMC Genet. 2014, 15, 35. [Google Scholar] [CrossRef]
- Lü, Y.; Cui, X.; Li, R.; Huang, P.; Zong, J.; Yao, D.; Li, G.; Zhang, D.; Yuan, Z. Development of genome-wide insertion/deletion markers in rice based on graphic pipeline platform. J. Integr. Plant Biol. 2015, 57, 980–991. [Google Scholar] [CrossRef]
- Mahmoodi, R.; Dadpour, M.R.; Hassani, D.; Zeinalabedini, M.; Vendramin, E.; Leslie, C.A. Composite core set construction and diversity analysis of Iranian walnut germplasm using molecular markers and phenotypic traits. PLoS ONE 2021, 16, e0248623. [Google Scholar] [CrossRef]
- Abbasi, Z.; Arzani, A.; Majidi, M.M. Evaluation of Genetic Diversity of Sugar Beet (Beta vulgaris L.) Crossing Parents Using Agro-morphological Traits and Molecular Markers. J. Agric. Ence Technol. 2014, 16, 1397–1411. Available online: https://jast.moares.ac.ir/article-23-6906-en.pdf (accessed on 10 September 2023).
- Chalbi, A.; Chikh-Rouhou, H.; Mezghani, N.; Slim, A.; Fayos, O.; Bel-Kadhi, M.S.; Garcés-Claver, A. Genetic Diversity Analysis of Onion (Allium cepa L.) from the Arid Region of Tunisia Using Phenotypic Traits and SSR Markers. Horticulturae 2023, 9, 1098. [Google Scholar] [CrossRef]
- Cui, P. Descriptors and Date Standard for Beet (Beta vulgaris L.); China Agriculture Press: Beijing, China, 2006; pp. 8–42. [Google Scholar]
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Springer: Berlin/Heidelberg, Germany, 1991; pp. 283–293. [Google Scholar] [CrossRef]
- Fugate, K.K.; Fajardo, D.; Schlautman, B.; Ferrareze, J.P.; Bolton, M.D.; Campbell, L.G.; Wiesman, E.; Zalapa, J. Generation and Characterization of a Sugarbeet Transcriptome and Transcript-Based SSR Markers. Plant Genome 2014, 7, plantgenome2013-11. [Google Scholar] [CrossRef]
- Yeh, F. Population genetic analysis of co-dominant and dominant markers and quantitative traits. Belg. J. Bot 1997, 129, 157. [Google Scholar]
- Ross-Ibarra, J. Genetic Data Analysis II. Methods for Discrete Population Genentic Data. Econ. Bot. 2002, 56, 216. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.; Vonholdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Bedeian, A.G.; Mossholder, K.W. On the use of the coefficient of variation as a measure of diversity. Organ. Res. Methods 2000, 3, 285–297. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; p. 117. [Google Scholar]
- Ortiz-Burgos, S. Shannon-Weaver Diversity Index. In Encyclopedia of Estuaries; Kennish, M.J., Ed.; Springer: Dordrecht, The Netherlands, 2016; pp. 572–573. [Google Scholar]
- Patterson, N.J.; Price, A.L.; Reich, D. Population Structure and Eigenanalysis. PLoS Genet. 2006, 2, e190. [Google Scholar] [CrossRef] [PubMed]
- Kleine, M.; Voss, H.H.; Cai, D.; Jung, C. Evaluation of nematode-resistant sugar beet (Beta vulgaris L.) lines by molecular analysis. Theor. Appl. Genet. 1998, 97, 896–904. [Google Scholar] [CrossRef]
- Saccomani, M.; Stevanato, P.; Trebbi, D.; McGrath, J.M.; Biancardi, E. Molecular and morpho-physiological characterization of sea, ruderal and cultivated beets. Euphytica 2009, 169, 19–29. [Google Scholar] [CrossRef]
- Taguchi, K.; Kuroda, Y.; Okazaki, K.; Yamasaki, M. Genetic and phenotypic assessment of sugar beet (Beta vulgaris L. subsp. vulgaris) elite inbred lines selected in Japan during the past 50 years. Breed Sci 2019, 69, 255–265. [Google Scholar] [CrossRef]
- Abekova, A.M.; Yerzhebayeva, R.S.; Bastaubayeva, S.O.; Konusbekov, K.; Bazylova, T.A.; Babissekova, D.I.; Amangeldiyeva, A.A. Assessment of sugar beet genetic diversity in the Republic of Kazakhstan by using RAPD markers and agromorphological traits. Sabrao J. Breed. Genet. 2022, 54, 67–78. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Na | Ne | I | Ho | He | Nei’s Expected Heterozygosity | Nm | Genetic Diversity | PIC |
|---|---|---|---|---|---|---|---|---|---|
| 14118 | 4 | 3.116 | 1.224 | 1.000 | 0.682 | 0.679 | 0.660 | 0.684 | 0.624 |
| 17623 | 5 | 3.124 | 1.349 | 0.961 | 0.683 | 0.680 | 0.603 | 0.680 | 0.640 |
| 2305 | 5 | 2.941 | 1.237 | 0.946 | 0.663 | 0.660 | 0.632 | 0.660 | 0.602 |
| 11965 | 5 | 4.104 | 1.455 | 0.985 | 0.759 | 0.756 | 0.466 | 0.756 | 0.713 |
| 27374 | 4 | 2.560 | 1.080 | 0.891 | 0.612 | 0.609 | 0.637 | 0.615 | 0.544 |
| BQ588629 | 5 | 3.325 | 1.339 | 0.992 | 0.702 | 0.699 | 0.611 | 0.699 | 0.651 |
| L37 | 4 | 2.765 | 1.114 | 1.000 | 0.641 | 0.638 | 0.904 | 0.638 | 0.566 |
| L7 | 6 | 3.655 | 1.498 | 0.953 | 0.729 | 0.726 | 0.441 | 0.735 | 0.699 |
| 7492 | 4 | 2.595 | 1.097 | 0.929 | 0.617 | 0.615 | 0.634 | 0.632 | 0.574 |
| L47 | 3 | 2.702 | 1.042 | 0.992 | 0.632 | 0.630 | 0.927 | 0.630 | 0.555 |
| L57 | 6 | 2.492 | 1.107 | 0.977 | 0.601 | 0.599 | 1.107 | 0.599 | 0.523 |
| L35 | 5 | 3.196 | 1.264 | 0.935 | 0.690 | 0.687 | 0.413 | 0.713 | 0.663 |
| L48 | 4 | 2.878 | 1.182 | 0.953 | 0.655 | 0.653 | 0.639 | 0.658 | 0.600 |
| L64 | 4 | 3.247 | 1.240 | 0.898 | 0.695 | 0.692 | 0.444 | 0.697 | 0.638 |
| W21 | 5 | 2.372 | 1.078 | 0.781 | 0.581 | 0.578 | 0.380 | 0.615 | 0.561 |
| 2170 | 3 | 2.090 | 0.787 | 0.906 | 0.524 | 0.522 | 1.418 | 0.529 | 0.419 |
| 15915 | 5 | 2.773 | 1.182 | 0.960 | 0.642 | 0.639 | 0.561 | 0.665 | 0.610 |
| 17923 | 8 | 3.445 | 1.468 | 0.977 | 0.713 | 0.710 | 0.552 | 0.710 | 0.664 |
| 24552 | 5 | 2.825 | 1.128 | 0.961 | 0.649 | 0.646 | 0.682 | 0.652 | 0.582 |
| 26319 | 9 | 4.870 | 1.810 | 0.953 | 0.798 | 0.795 | 0.364 | 0.798 | 0.773 |
| 57236 | 4 | 3.315 | 1.249 | 0.954 | 0.701 | 0.698 | 0.538 | 0.698 | 0.637 |
| SSD6 | 6 | 3.302 | 1.450 | 0.992 | 0.700 | 0.697 | 0.617 | 0.697 | 0.661 |
| TC94 | 3 | 2.492 | 0.982 | 1.000 | 0.601 | 0.599 | 1.267 | 0.599 | 0.516 |
| BVV23 | 3 | 2.950 | 1.090 | 0.961 | 0.664 | 0.661 | 0.666 | 0.661 | 0.587 |
| W15 | 4 | 2.655 | 1.074 | 0.921 | 0.626 | 0.623 | 0.625 | 0.635 | 0.562 |
| BVV21 | 4 | 2.587 | 1.080 | 0.952 | 0.616 | 0.613 | 0.656 | 0.636 | 0.571 |
| TC122 | 7 | 4.079 | 1.627 | 0.985 | 0.758 | 0.755 | 0.469 | 0.755 | 0.722 |
| Average | 4.815 | 3.054 | 1.231 | 0.952 | 0.664 | 0.662 | 0.599 | 0.668 | 0.610 |
| Locus | Na | Ne | I | Ho | He | Nei’s Expected Heterozygosity | Nm | Genetic Diversity | PIC |
|---|---|---|---|---|---|---|---|---|---|
| ND31 | 3 | 2.486 | 1.002 | 0.734 | 0.600 | 0.598 | 0.380 | 0.598 | 0.531 |
| ND75 | 3 | 1.876 | 0.729 | 0.636 | 0.469 | 0.467 | 0.533 | 0.467 | 0.378 |
| ND109 | 3 | 2.269 | 0.898 | 0.892 | 0.561 | 0.559 | 0.982 | 0.559 | 0.465 |
| ND113 | 3 | 1.825 | 0.663 | 0.682 | 0.454 | 0.452 | 0.768 | 0.452 | 0.353 |
| ND220 | 3 | 2.768 | 1.053 | 0.813 | 0.641 | 0.639 | 0.341 | 0.639 | 0.562 |
| ND223 | 3 | 1.750 | 0.702 | 0.558 | 0.430 | 0.429 | 0.467 | 0.429 | 0.360 |
| ND253 | 5 | 2.424 | 1.042 | 0.859 | 0.590 | 0.588 | 0.636 | 0.588 | 0.508 |
| ND258 | 3 | 2.856 | 1.074 | 0.954 | 0.652 | 0.650 | 0.688 | 0.650 | 0.576 |
| ND275 | 4 | 2.204 | 0.925 | 0.852 | 0.548 | 0.546 | 0.808 | 0.546 | 0.459 |
| ND277 | 3 | 1.804 | 0.791 | 0.566 | 0.447 | 0.446 | 0.435 | 0.446 | 0.404 |
| ND10 | 4 | 1.985 | 0.741 | 0.758 | 0.498 | 0.496 | 0.735 | 0.496 | 0.384 |
| ND11 | 3 | 1.808 | 0.658 | 0.659 | 0.449 | 0.447 | 0.701 | 0.447 | 0.351 |
| ND18 | 2 | 1.162 | 0.268 | 0.057 | 0.140 | 0.140 | 0.015 | 0.140 | 0.130 |
| ND108 | 4 | 2.844 | 1.115 | 0.914 | 0.651 | 0.648 | 0.566 | 0.648 | 0.576 |
| ND120 | 3 | 2.015 | 0.716 | 0.969 | 0.506 | 0.504 | 6.319 | 0.504 | 0.381 |
| ND121 | 3 | 2.579 | 1.004 | 0.876 | 0.615 | 0.612 | 0.629 | 0.612 | 0.530 |
| ND129 | 4 | 3.008 | 1.175 | 0.845 | 0.670 | 0.668 | 0.431 | 0.668 | 0.602 |
| ND142 | 3 | 2.428 | 0.956 | 0.954 | 0.590 | 0.588 | 1.070 | 0.588 | 0.499 |
| ND143 | 4 | 1.581 | 0.696 | 0.426 | 0.369 | 0.368 | 0.345 | 0.368 | 0.336 |
| ND162 | 4 | 2.577 | 1.052 | 0.984 | 0.614 | 0.612 | 0.942 | 0.612 | 0.533 |
| ND229 | 3 | 2.062 | 0.762 | 0.985 | 0.517 | 0.515 | 5.382 | 0.515 | 0.398 |
| ND231 | 4 | 2.931 | 1.104 | 0.930 | 0.661 | 0.659 | 0.600 | 0.659 | 0.586 |
| ND233 | 3 | 2.711 | 1.046 | 0.915 | 0.634 | 0.631 | 0.658 | 0.631 | 0.558 |
| ND243 | 5 | 2.701 | 1.104 | 0.915 | 0.632 | 0.630 | 0.663 | 0.630 | 0.564 |
| ND244 | 3 | 2.913 | 1.084 | 0.954 | 0.659 | 0.657 | 0.662 | 0.657 | 0.583 |
| ND246 | 2 | 1.928 | 0.674 | 0.667 | 0.483 | 0.481 | 0.564 | 0.481 | 0.365 |
| ND267 | 3 | 2.606 | 1.018 | 0.977 | 0.619 | 0.616 | 0.879 | 0.616 | 0.540 |
| ND270 | 4 | 2.325 | 0.959 | 0.868 | 0.572 | 0.570 | 0.799 | 0.570 | 0.479 |
| ND279 | 3 | 2.226 | 0.873 | 0.922 | 0.553 | 0.551 | 1.140 | 0.551 | 0.451 |
| ND280 | 2 | 1.555 | 0.542 | 0.434 | 0.358 | 0.357 | 0.388 | 0.357 | 0.293 |
| ND281 | 3 | 2.806 | 1.065 | 0.930 | 0.646 | 0.644 | 0.614 | 0.644 | 0.571 |
| ND283 | 2 | 1.961 | 0.683 | 0.813 | 0.492 | 0.490 | 1.062 | 0.490 | 0.370 |
| ND286 | 3 | 2.865 | 1.074 | 0.961 | 0.654 | 0.651 | 0.705 | 0.651 | 0.576 |
| Average | 3.242 | 2.298 | 0.886 | 0.796 | 0.545 | 0.543 | 0.660 | 0.543 | 0.462 |
| Trait | Max (cm) * | Min (cm) ** | Average (cm) | SD | CV (%) |
|---|---|---|---|---|---|
| Petiole width | 2.760 | 1.075 | 1.596 | 0.266 | 16.64 |
| Petiole length | 38.940 | 18.030 | 26.663 | 4.562 | 17.11 |
| Plant height | 69.450 | 41.625 | 53.566 | 5.211 | 9.73 |
| Width of leaf coverage | 103.260 | 54.620 | 79.442 | 11.990 | 15.09 |
| Traits | Characteristic Description (Proportion of Distribution, %) | H′ | |||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | ||
| Leaf shape | Halberd | Share | Tongue | 1.074 | |||
| 23.26 | 38.76 | 37.98 | |||||
| Leaf color | Light green | Green | Dark green | 1.057 | |||
| 20.93 | 43.41 | 35.66 | |||||
| Leaf margin shape | Full margin | Small wave | Medium wave | Big wave | 1.237 | ||
| 5.43 | 29.46 | 26.36 | 38.76 | ||||
| Leaf surface | Smooth | Wavy | Slight crease | More creases | 1.146 | ||
| 2.33 | 23.26 | 44.96 | 29.46 | ||||
| Mesophyll thickness | Thin | Medium | Thick | 1.041 | |||
| 58.91 | 33.33 | 31.01 | |||||
| Leaf hairiness | Little | Much | 0.138 | ||||
| 96.90 | 3.10 | ||||||
| Petiole color | White | White-green | Light green | Green | Pink | Purplish red | 1.034 |
| 3.88 | 60.47 | 27.13 | 6.98 | 0.00 | 1.55 | ||
| Petiole thickness | Slight | Medium | Thick | 1.065 | |||
| 28.68 | 45.74 | 25.58 | |||||
| Fascicled leaf type | Erect | Semi-crawl | Crawl | 0.810 | |||
| 61.24 | 34.11 | 4.65 | |||||
| Root length | Very short | Short | Medium | Long | Very long | 1.395 | |
| 3.10 | 13.18 | 37.21 | 31.01 | 15.50 | |||
| Root width | Narrow | Medium | Broad | 1.082 | |||
| 26.36 | 32.56 | 41.09 | |||||
| Root length-to-width ratio | Small | Medium | Large | 1.051 | |||
| 30.23 | 22.48 | 47.29 | |||||
| Root shape | Cuneiform | Conical | Spindle | 1.000 | |||
| 51.94 | 31.78 | 16.28 | |||||
| Crown size | Small | Medium | Large | 1.068 | |||
| 43.41 | 33.33 | 23.26 | |||||
| Root groove depth | None | Not obvious | Shallow | Deep | 1.240 | ||
| 27.91 | 37.98 | 28.68 | 5.43 | ||||
| Skin roughness | Very smooth | Smoother | Very rough | 0.965 | |||
| 49.61 | 38.76 | 11.63 | |||||
| Trait | Petiole Width | Petiole Length | Plant Height |
|---|---|---|---|
| Petiole length | 0.361 ** | ||
| Plant height | 0.392 ** | 0.788 ** | |
| Width of leaf coverage | 0.441 ** | 0.642 ** | 0.444 ** |
| Principal Component Number | Eigenvalue | Contribution Rate (%) | Cumulative Contribution Rate (%) |
|---|---|---|---|
| 1 | 3.615 | 18.075 | 18.075 |
| 2 | 1.903 | 9.516 | 27.590 |
| 3 | 1.853 | 9.263 | 36.854 |
| 4 | 1.480 | 7.399 | 44.252 |
| 5 | 1.380 | 6.899 | 51.151 |
| 6 | 1.173 | 5.866 | 57.017 |
| 7 | 1.072 | 5.359 | 62.376 |
| 8 | 1.068 | 5.340 | 67.716 |
| 9 | 0.968 | 4.838 | 72.555 |
| 10 | 0.850 | 4.252 | 76.807 |
| 11 | 0.804 | 4.020 | 80.827 |
| 12 | 0.691 | 3.454 | 84.281 |
| 13 | 0.678 | 3.389 | 87.669 |
| 14 | 0.620 | 3.102 | 90.771 |
| 15 | 0.466 | 2.328 | 93.099 |
| 16 | 0.434 | 2.172 | 95.271 |
| 17 | 0.395 | 1.977 | 97.249 |
| 18 | 0.295 | 1.476 | 98.725 |
| 19 | 0.142 | 0.710 | 99.435 |
| 20 | 0.113 | 0.565 | 100.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, F.; Pi, Z.; Li, S.; Wu, Z. Genetic Diversity and Population Structure Analysis of Excellent Sugar Beet (Beta vulgaris L.) Germplasm Resources. Horticulturae 2024, 10, 120. https://doi.org/10.3390/horticulturae10020120
Peng F, Pi Z, Li S, Wu Z. Genetic Diversity and Population Structure Analysis of Excellent Sugar Beet (Beta vulgaris L.) Germplasm Resources. Horticulturae. 2024; 10(2):120. https://doi.org/10.3390/horticulturae10020120
Chicago/Turabian StylePeng, Fei, Zhi Pi, Shengnan Li, and Zedong Wu. 2024. "Genetic Diversity and Population Structure Analysis of Excellent Sugar Beet (Beta vulgaris L.) Germplasm Resources" Horticulturae 10, no. 2: 120. https://doi.org/10.3390/horticulturae10020120
APA StylePeng, F., Pi, Z., Li, S., & Wu, Z. (2024). Genetic Diversity and Population Structure Analysis of Excellent Sugar Beet (Beta vulgaris L.) Germplasm Resources. Horticulturae, 10(2), 120. https://doi.org/10.3390/horticulturae10020120