Abstract
Camellia oleifera Abel is rich in oil, which is beneficial to human health. Seasonal drought and uneven canopy microclimate distributions result in low yields and picking efficiencies. To reveal the features of fruit development in the canopy and evaluate the impact of pruning on the canopy microclimate and fruit parameters, a field experiment was carried out in Zhuzhou, Hunan. The C. oleifera tree canopy was divided into four positions: inside the upper canopy (UI); inside the lower canopy (LI); outside the upper canopy (UO); and outside the lower canopy (LO). A combination of high-light efficiency tree pruning and mulching with camellia shells and weedproof film (CPM) was tested and compared with mulching (M). Compared with temperature and humidity, canopy light intensity is a key climate factor that affects the development of Camellia oleifera fruits. CPM improved the canopy light distribution and increased the light intensity inside the canopy of C. oleifera. There was no significant difference in the fruit cracking rate among the four canopy positions in the CPM treatment, which indicated that the fruit maturation period tended to be consistent across the canopy positions. Treatment with CPM resulted in 24.14% and 4.11% greater yield and oil content ratios, respectively, than the treatment with M. The fresh weight of the fruits from LO was the lowest, and the fatty acid accumulation was the lowest due to the poor light availability. CMP increased fruit quality, particularly in LI, which was 41.92%, 36.36%, 13.21%, and 12.79% greater in terms of fresh weight, yield, fresh seed rate, and oil content, respectively, than M. Unfortunately, there was no discernible difference in fatty acid components between CPM and M. Our research suggested that pruning combined with the mulching of organic material is an effective way to alleviate seasonal drought and uneven fruit maturation in C. oleifera trees, further increasing yield and fruit quality.
1. Introduction
Camellia oleifera Abel, an important economic forest species, is widely grown in subtropical regions, including China, India, Japan, and Southeast Asia. More than 90% of the global production is in China [1,2,3]. Camellia oil extracted from the seeds of C. oleifera is often regarded as an ‘oriental olive oil’ due to its extremely similar fatty acid profiles and physicochemical properties to olive oil [4,5]. Camellia oil contains large amounts of unsaturated fatty acids (more than 80% of total fatty acids), polyphenols, tea saponins, etc. These biologically active compounds are beneficial to human health [4,5,6,7,8]. Therefore, camellia oil is gaining popularity not only as a high-quality edible oil but also as a functional food. However, the key problems of improving the yield and seed oil content of C. oleifera and the efficiency of mechanized harvesting still need to be solved in C. oleifera production.
The uneven distribution of annual rainfall seriously threatens the production of C. oleifera. Seasonal drought mainly occurs from July to December (Supplementary Table S1). Data from previous studies suggested that the fruit volume expanded rapidly from July to August. After August, the fruit volume did not change significantly, and the oil accumulated quickly until maturity [9,10,11]. Although C. oleifera is classified as a drought-tolerant tree species, the fruit expansion period and oil accumulation period are highly sensitive to water, and these are the physiological periods during which water is needed. Soil water deficit seriously threatens fruit yield and quality. Covering the soil surface with mulch is a cropland management technique that can increase soil moisture, efficiently reduce soil temperature, and improve soil structure by restricting the transport of water from the soil to air and reducing evaporative losses, which are beneficial for improving plant water availability [12,13,14,15,16]. Commonly used covering materials are divided into three types: organic materials (e.g., straw); inorganic materials (e.g., plastic films); and other materials, such as stones and higher-fiber materials [13,17,18,19]. Organic mulch is an essential agricultural measure that not only effectively utilizes a large amount of agricultural waste to avoid polluting the environment through incineration but is also very effective for soil and water conservation [20,21]. In recent years, much research has been carried out to investigate the relationship between the organic mulch environment and crop development. Mulching with compost significantly increased the quality and yield (~1 t/ha) of grapes in low-yield areas, which was attributed to improved soil moisture and temperature [22]. In a two-year rotation field in the Loess Plateau region of China, the yearly crop yield increased with straw mulching in both cycles, while with plastic film mulching, the increase occurred only in cycle 2. Compared with that under plastic film mulching, the greenhouse gas emission intensity under straw mulching was 126% lower. The soil temperature exhibited the opposite trend between plastic film mulching and straw mulching, whereas the soil moisture increased in both mulching treatments [23]. Newspaper mulching could be an alternative option compared to rice straw mulching because it provides a greater yield but has lower soil–water consumption for rain-fed soybean cultivation [24]. Of the six mulching materials, i.e., an ecological pad, straw, straw and ecological pad, gravel, sawdust, and camellia shells, organic material worked best to boost oil yield during seasonal drought in C. oleifera hilly areas without irrigation. For mulching with an ecological pad, the yield was 58%, 61%, and 124% greater than that of unmulched conditions for ‘Hengdongdatao’, ‘Huashuo’, and ‘Xianglin 210’, respectively [14]. Therefore, improving the soil environment by organic mulching is very effective for fruit development.
The management of canopy architecture is one of the most important factors for the productivity of fruit crops [25]. Incorrect tree structure and poor canopy management practices affect the uniformity of fruit development, which threatens fruit yield and quality at harvest while also reducing the efficiency of mechanized harvesting. Therefore, additional details about the canopy microclimate and canopy management have received extensive attention from researchers [26,27,28]. The main canopy factors are temperature, light intensity, and relative humidity [29]. The light intensity and temperature in the upper and outer layers were greater than those in the central and lower layers. The relative humidity in the lower and inner layers was greater than that in the upper layer [30]. Additionally, light intensity was strongly correlated with yield and fruit quality, with a correlation coefficient greater than 0.8 in C. oleifera [31]. For the fruit set percentage, the seed yield in the outer (or upper) canopy was significantly greater than that in the inner (or lower) canopy. However, fruit quality differs among canopy layers in some species. The oil content in the seeds of the lower canopy was greater than that in the upper canopy in Xanthoceras sorbifolium, and the opposite was true in Sapindus mukorossi [30,32]. Canopy manipulation by pruning, plant growth inhibition, and dwarfing rootstocks plays an important role in regulating vegetative growth, flowering, and fruit yield [25]. The pruning of trees affects the quality of sunlight intercepted, as the crown profile determines the extent to which the leaf area is exposed to incoming radiation. Summer pruning reduced whole-canopy transpiration in apple trees [33]. Winter lateral pruning and summer toppling for high-density olive orchards contributed to higher yields and more compact tree shapes because of a good vegetable-reproductive balance [34]. The microclimate, especially the light intensity, was reported to be more uniformly distributed, which contributed to greater uniformity in fruit quality within open-centered canopy trees than within round-headed canopy trees [35]. In Camellia oleifera orchards, due to extensive management, uneven canopy structure leads to inconsistent fruit maturity, which is not conducive to improving harvesting efficiency. Moreover, there is currently a lack of varieties with more open canopy. However, studies on fruit maturity consistency at harvest are rare.
Considerable research has been carried out on the quality improvement and canopy environment of C. oleifera. To alleviate the drought stress faced by C. oleifera during the growing season, our laboratory evaluated the soil water retention effect of various mulching materials and their effects on yield and quality and determined that organic mulching was an ideal mulching material [14]. In addition, our laboratory has also carried out canopy microclimate research on multiple tree species and has shown that thinning flowers or fruits, adjusting the type of fruiting branch group, or changing the tree structure can regulate yield and quality [30,31,36,37]. Unfortunately, studies on the combination of aboveground and belowground regulatory techniques are rare. Therefore, the purpose of this research project was to investigate whether a combination of C. oleifera tree pruning and mulching could improve the canopy environment, yield, and quality and promote fruit maturation. This study was conducted over two years. In the first year (2021), the microclimatic and fruit development characteristics of the four canopy positions (inside the upper canopy, inside the lower canopy, outside the upper canopy, and outside the lower canopy) were determined. In February of the second year (2022), high-light efficiency pruning of the trees was carried out, and in June, the tree plate was covered with camellia shells and weedproof film. The goal of this effort was to determine whether the combination of pruning and mulching can affect yield and oil quality, as well as the uniformity of fruit ripening among the four canopy positions. We hope to provide a feasible solution for relieving seasonal drought and ensure that canopy environment distribution does not lead to reduced yield and uneven fruit ripening speeds in C. oleifera.
2. Materials and Methods
2.1. Experimental Area and Plant Materials
This study was carried out in Zhuzhou, Hunan, China (113°14′ E, 27°62′ N). This area has a characteristic subtropical monsoon climate with a mean annual precipitation of 1400 mm and an acid red soil type. The climate-related information for the 2021–2022 cropping years is shown in Supplement Table S1. In the experimental area, the average monthly sunshine duration was 134 h in 2021 and 135.2 h in 2022. The average temperatures were 19.4 °C and 18.9 °C, respectively. Rainfall is primarily concentrated from February to June. Additionally, there is a seasonal drought in this area, lasting an average of up to five months from July to November.
Thirty-five-year-old trees with similar growth conditions and growth potentials without disease from the C. oleifera cultivar ‘Huaxin’ were randomly selected for this study as the optimal cultivar that the State Forestry Administration authorized for use and promotion across the country. The average height of these trees was 2 m, and the average crown breadth was 2.2 m × 2.2 m, with a natural round-headed shape.
2.2. Canopy Division and Tree Canopy Environmental Characteristics
C. oleifera trees were used to record changes in the canopy environment, including temperature, humidity, and light, from July to October. The experiment consisted of four canopy positions, namely, inside the upper canopy, outside the upper canopy, inside the lower canopy, and outside the lower canopy (UI, UO, LI, LO; Figure 1). The fruits on the inside canopy position are oriented at the lower edge of the tree and are always fully shaded, while the fruits from the outside canopy are the upper fruits of the tree that are always fully exposed to sunlight. Canopy microclimate factors during fruit development were observed 3 days per month under clear sky conditions. The light intensity, air temperature, and air relative humidity were measured four times a day at 8:00, 11:00, 14:00, and 17:00. Light intensity was measured using a handheld luminometer (TES, 1339R). The air temperature and relative humidity in the canopy were measured using a digital thermometer (SMART, SENSOR AR837).
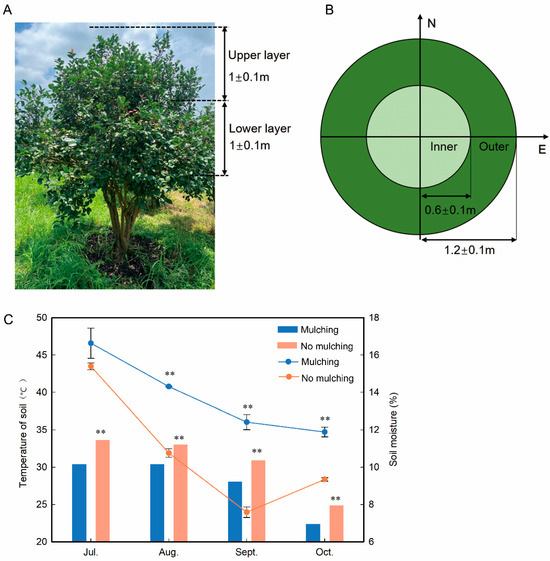
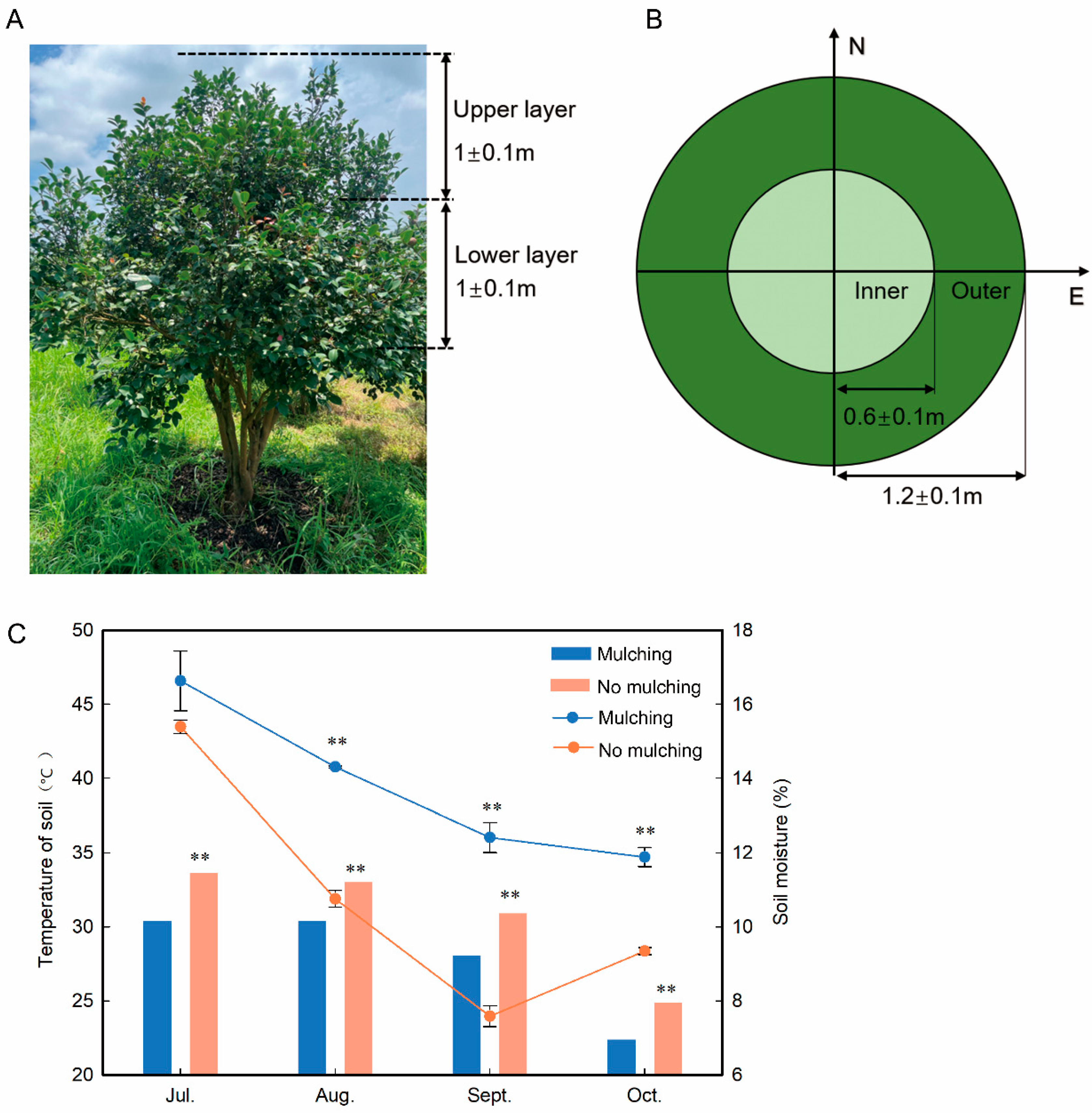
Figure 1.
Layout diagram of areas and soil temperature and moisture changes in C. oleifera trees. (A) front view of canopy partitioning; (B) vertical view of canopy partitioning; (C) effect of mulching with camellia shells and weedproof film on soil temperature and moisture compared with no mulching. N, north; E, east. One-way ANOVAs for mulching and no mulching are displayed, indicating ** for significance at p ≤ 0.01.
2.3. The Combination of High-Light Efficiency Pruning and Mulching of C. oleifera Trees
The study area experienced low total rainfall, especially from August until October. Our laboratory’s findings unequivocally demonstrated that mulching with camellia shells and weedproof film preserved soil moisture better than no mulching (Figure 1C). The average water content of the soil under mulching was 63.53% greater than that under no mulching in September 2020.
Therefore, the experimental setting included two treatments on the basis of mulching: mulching with camellia shells and weedproof film (M); and a combination of high-light efficiency pruning of shoots and mulching (CPM). Pruning was conducted on 28 February 2022. We removed the weak, diseased branches and overlapping shoots. Additionally, long shoots were slightly truncated. To ensure the annual yield, a small amount of flower thinning was carried out on the shoots with large flowers. Through these operations, the light utilization efficiency of the inner and lower layers of the tree canopy improved, but the tree shape did not change. Mulching was carried out on 30 June 2022, using camellia shells with a thickness of 7–10 cm and a weedproof film.
2.4. Fruit Sampling, Fruit Morphology, and Physiological Determination
After the trees bloomed, the developing fruits from the four canopy positions were collected on 3 July, and the fruits were collected every 7 or 10 days. Fruits were harvested on 1 and 3 November 2021 and 2022, respectively.
Forty-eight fruits were randomly selected, and 3 independent biological replicates with four fruits per replicate were collected from each canopy position at each sampling time point. After the determination of the fruit cracking rate, fresh fruit yield per tree, and quality (the average fresh weight of a single fruit, lateral diameter, longitudinal diameter, and the ratio of fresh fruit to seed), the seeds were placed in an oven (deactivated at 105 °C for 15 min and baked at 70 °C for 72 h). The Soxtec extraction method (FOSS Soxtec 8000, Denmark, Sweden) was used to extract the seed oil. The fatty acid composition of the oil in the seeds was measured via gas chromatography (YLSZJ-SB-287) [14]. The anthrone colorimetry method was used to measure the starch and soluble sugar concentrations in the extracts. The main parameters and calculation methods were expressed as follows [14]:
Fresh seed percentage (%) = Fresh seed weight/Fresh fruit weight × 100%;
Seed water content (%) = Seed dry weight/Kernel fresh weight × 100%;
Oil content ratio (%) = Oil content/Kernel dry weight × 100%.
2.5. Statistical Analysis
One-way analysis of variance (ANOVA) and Student’s t-test were used to analyze the data using IBM SPSS Statistics software version 26. Graphs were generated with GraphPad Prism software version 8.3.0 and SigmaPlot software version 12.5. p ≤ 0.05 was considered extremely significant in these analyses.
3. Results
3.1. Variation in the Microclimate within the Canopy during the Growing Season
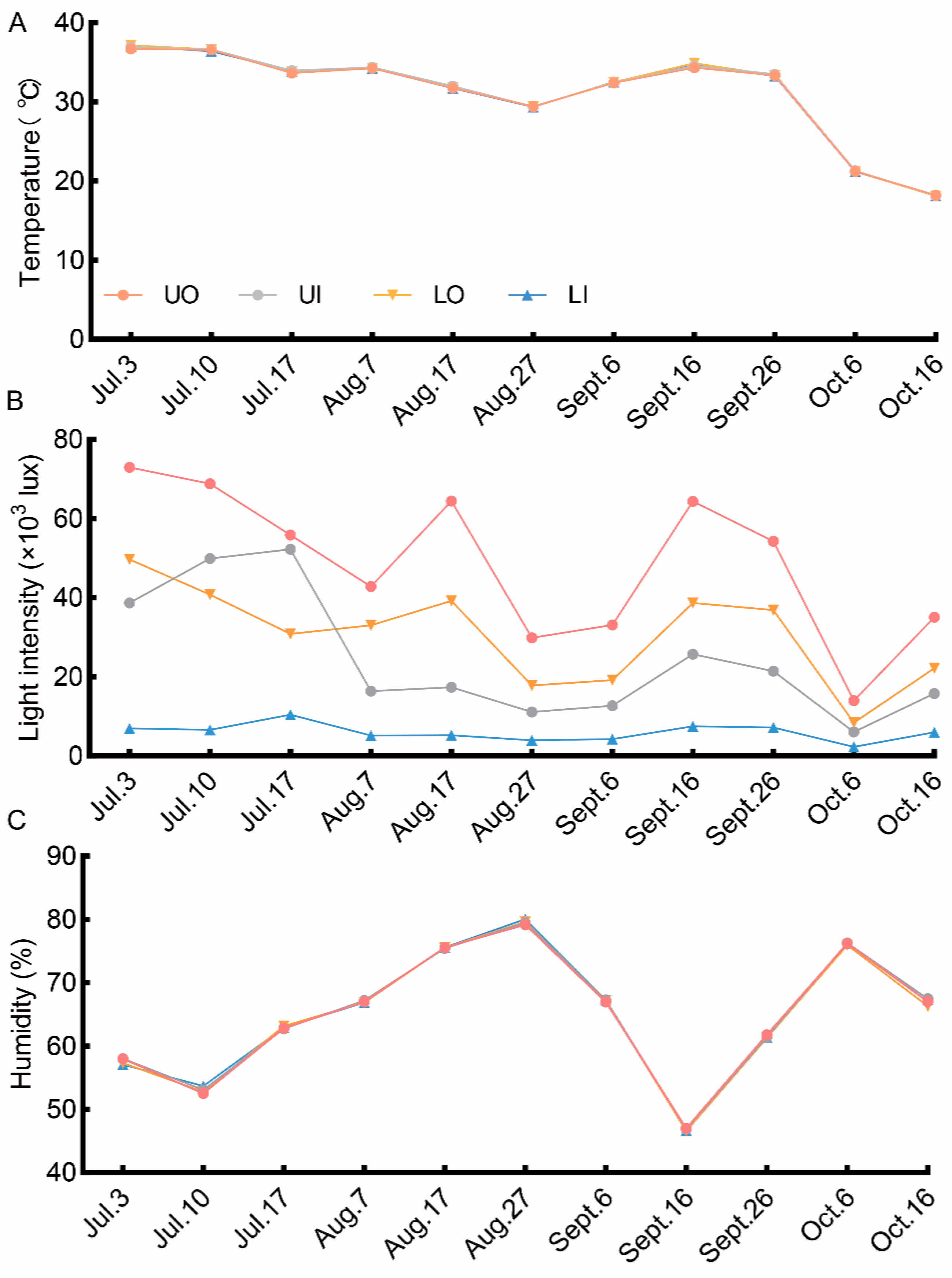
The analysis of the canopy environment measurements revealed that the dynamic variations in the four canopy positions followed the same general pattern, as shown in Figure 2. Humidity and temperature were similar at all canopy positions, while light intensity greatly differed depending on the canopy position. The order of canopy positions for maximum light intensity was UO > LO > UI > LI. Among all canopy positions, the upper canopy had the highest light intensity compared with the inside of the lower canopy. Therefore, the canopy position mainly affected the distribution of light.
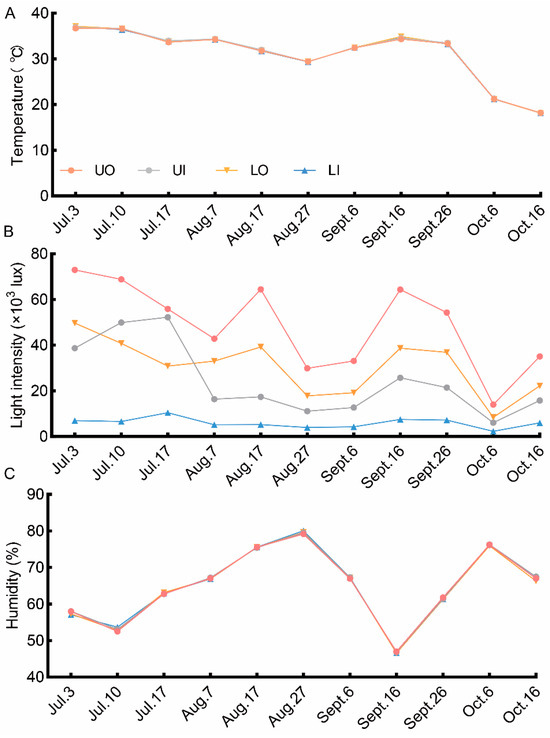
Figure 2.
Canopy environmental characteristics during the fruit development in 2021. (A) dynamic change in temperature at the four canopy positions; (B) dynamic change in light intensity at the four canopy positions; (C) dynamic change in humidity at the four canopy positions. UO, outside the upper canopy; UI, inside the upper canopy; LO, outside the lower canopy; LI, inside the lower canopy.
3.2. Characteristics of Fruit Development at Different Canopy Positions
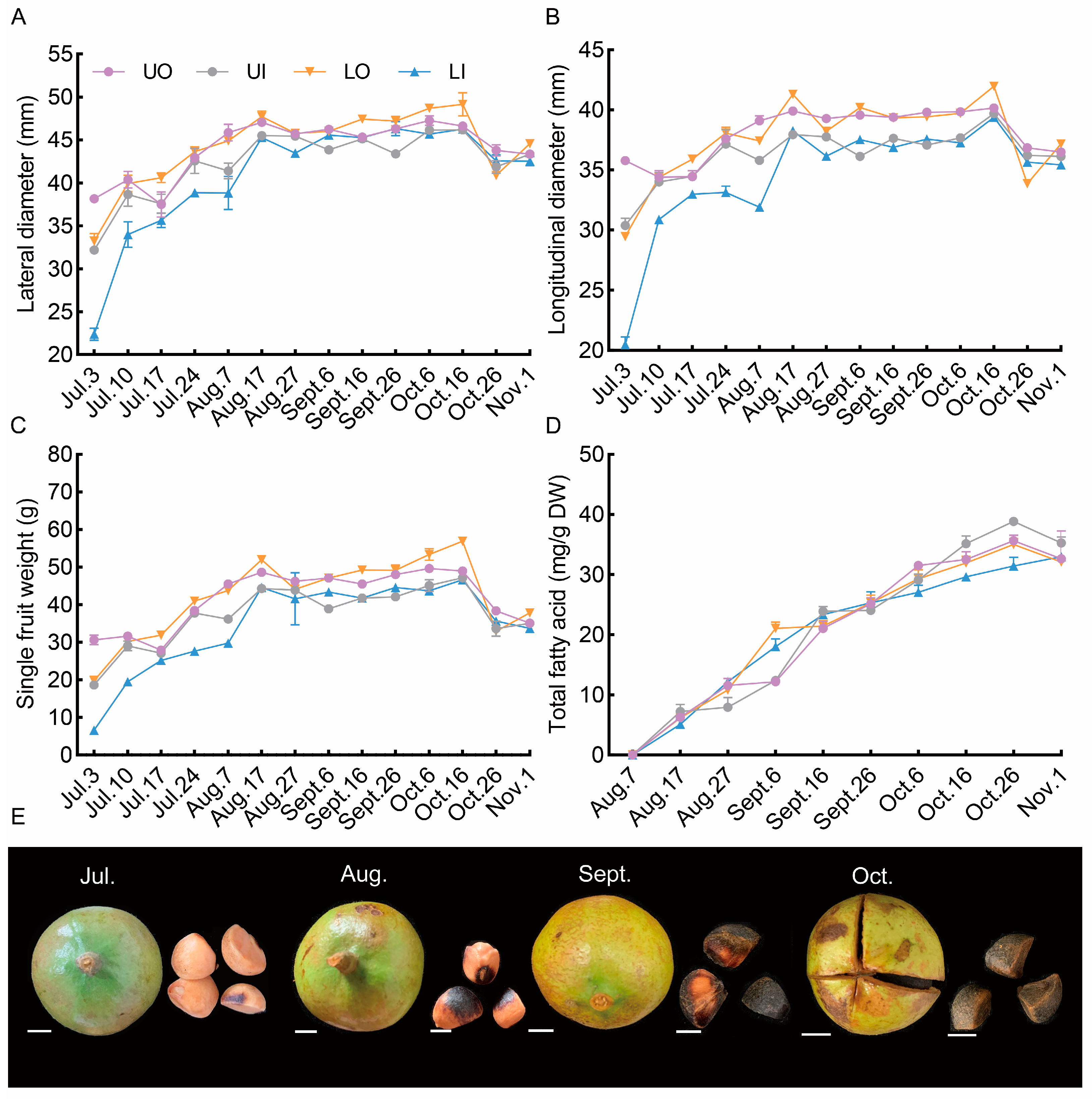
The fruit development characteristics among the four canopy positions are shown in Figure 3. Lateral diameter, longitudinal diameter, and single-fruit fresh weight exhibited similar dynamic changes at all canopy positions. The rapid expansion of fruit occurred before 17 August, after which the fruit morphology tended to stabilize. However, the fruit morphology declined slightly before harvest. The total fatty acid content increased rapidly, and the seed coat gradually became black and hard after August. This showed that quality fruit began to form in mid-August.
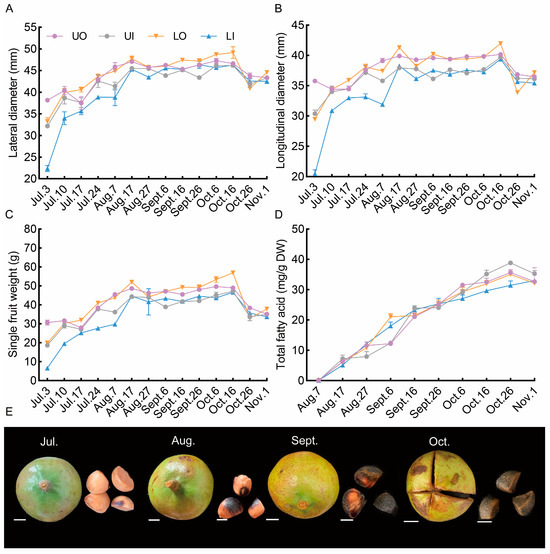
Figure 3.
Dynamic effect of different canopy positions on fruit development in 2021. (A) dynamic change in fruit lateral diameter; (B) dynamic change in fruit longitudinal diameter; (C) dynamic change in fruit weight; (D) dynamic accumulation of seed total fatty acids; (E) dynamic development of fruit and seeds. UO, outside of the upper canopy; UI, inside of the upper canopy; LO, outside of the lower canopy; LI, inside of the lower canopy. Bar = 10 mm.
The fruit morphology, single fruit fresh weight, and oil content of the seeds at the different canopy positions were strongly influenced by the microclimate (Figure 3). The fruits inside the canopy, especially the fruits inside the upper canopy, were smaller than those outside the canopy. As shown by the dynamic changes in oil accumulation, the oil content of the seeds inside the upper canopy continued to increase, while the seed oil content at the other three canopy positions tended to decrease after 26 October. These findings indicated that the fruit oil accumulation inside the lower canopy was slower than that of the outside canopy and that the fruits were not mature at harvest.
3.3. Response of the C. oleifera Canopy Microclimate to CPM
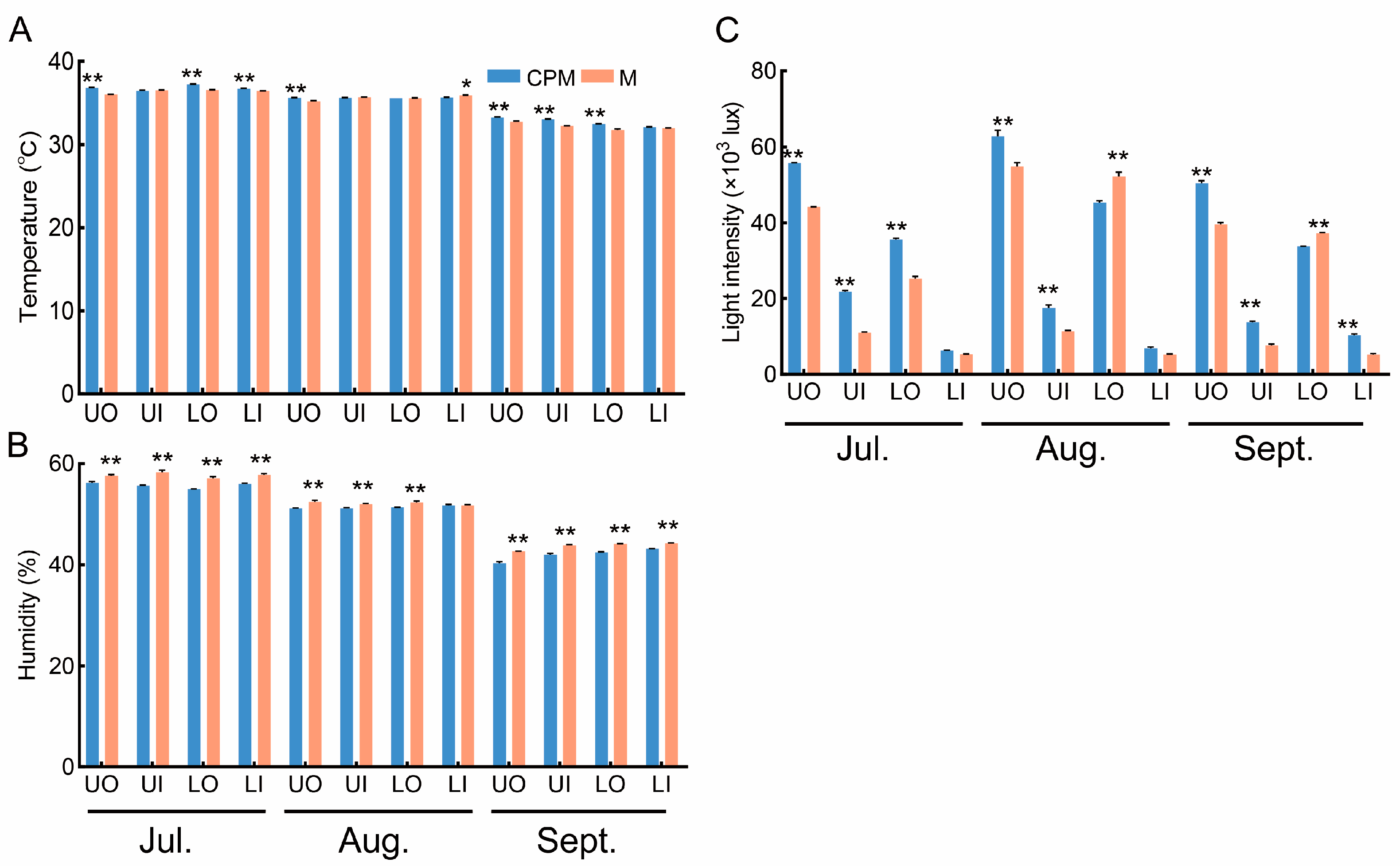
To improve fruit maturation uniformity, which is beneficial for mechanized harvesting, we carried out CPM on C. oleifera trees. The temperature, light intensity, and humidity under the different canopy positions were influenced by the CPM (Figure 4A–C). In comparison to that in the M treatment, the average temperature in the CPM treatment increased by 2.11%, 2.39%, 1.81%, and 1.02% for UO, UI, LO, and LI, respectively (Figure 4A). The humidity decreased by 5.46% (UO), 4.50% (UI), 3.82% (LO), and 3.22% (LI) in the CPM treatment compared with that in the M treatment (Figure 4B). The light intensity in the CPM treatment had the greatest increase (27.24%, 92.70%, 42.45%, and 85.15% for UO, UI, LO, and LI, respectively) compared with M (Figure 4B). Consequently, the light distribution across the four canopy positions, particularly the light intensity of the inner canopy, was considerably (p ≤ 0.05) improved by pruning the C. oleifera trees. Naturally, this affected the temperature and humidity of these canopy positions.
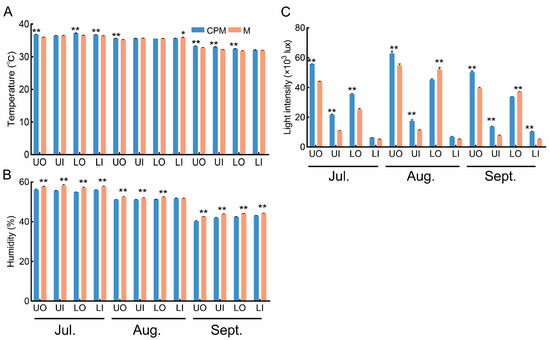
Figure 4.
The response of canopy environmental characteristics to CPM in 2022. (A) comparison of canopy temperature; (B) comparison of canopy light intensity; (C) comparison of canopy humidity. CPM, the combination of high-light efficiency pruning and mulching of C. oleifera trees; M, mulching of C. oleifera trees. UO, outside the upper canopy; UI, inside the upper canopy; LO, outside the lower canopy; LI, inside the lower canopy. ** indicates significance at p ≤ 0.01; * indicates significance at p ≤ 0.05.
3.4. CPM Promoted Fruit Cracking and Ripening in C. oleifera Trees
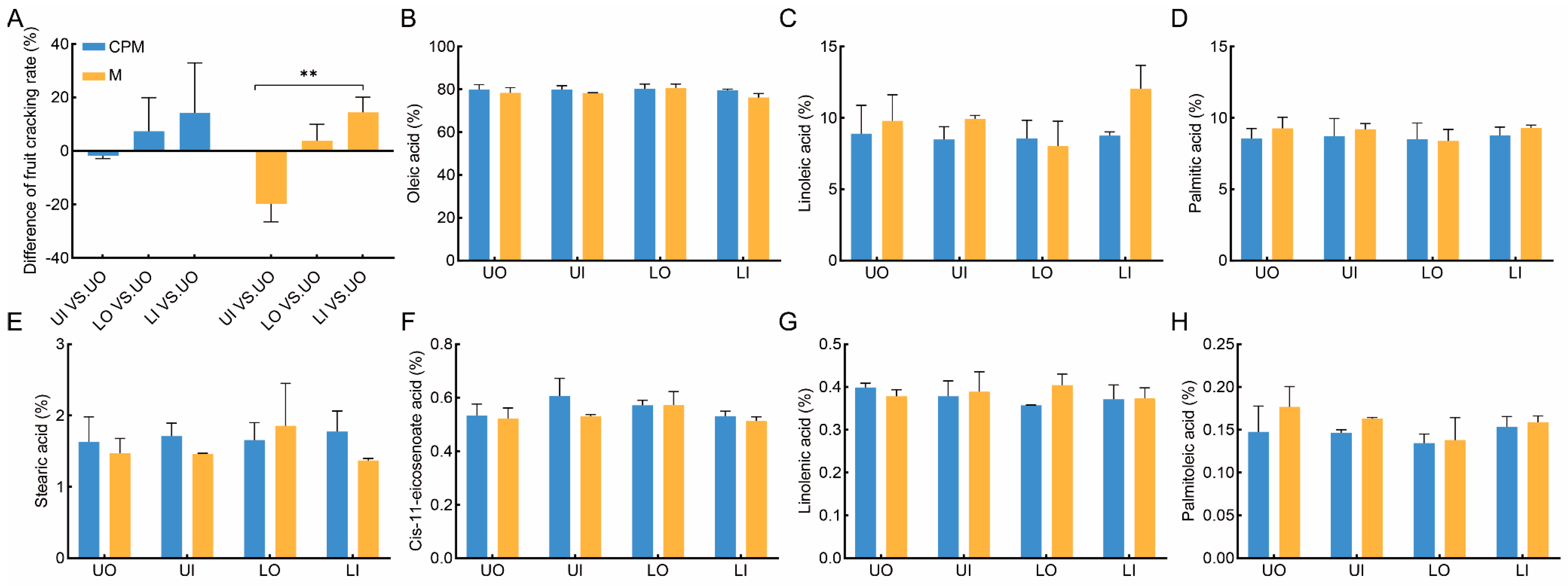
Fruit cracking is related to the maturity of the fruit. The greater the fruit cracking rate and gaps of fruit cracking, the greater the fruit maturity. The results of the fruit cracking rates for the four canopy positions are shown in Table 1. The fruit cracking rate significantly increased by 84.65% and 109.65% for LO and UI, respectively, for fruit cracking rate with gaps > 2. Unfortunately, compared to those in M, there were no significant differences in the fruit cracking rates of UO and LI (Table 1). In both the CPM and M, the fruit cracking rates in UI were 1.81% and 19.77% greater than those in UO; however, the fruit cracking rates in LO and LI were lower than those in UO, as shown in Figure 5A. This indicated that the maturity of the fruit in UI was greater than that in UO, but the maturity of the fruit in LI and LO was lower than that in UO. Compared to M, the difference in the fruit cracking rate was not significant between UI vs. UO and LI vs. UO in CPM (Figure 5A). These results showed that the high-light efficiency pruning of C. oleifera trees promoted consistent fruit cracking and fruit development at the four canopy positions.

Table 1.
Changes in the morphological indexes and maturity of C. oleifera ‘Huaxin’ in 2022.
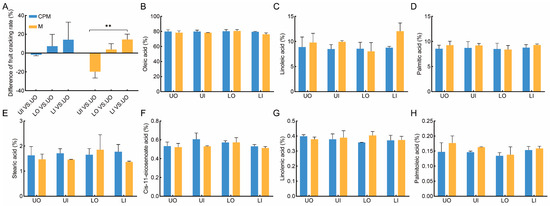
Figure 5.
The response of fruit ripening speed difference between canopies and the fatty acid compositions of kernels to CPM in 2022. (A) the difference in the fruit ripening speed; (B–H) the difference in oleic acid, linoleic acid, palmitic acid, stearic acid, cis-11-eicosenoate acid, linolenic acid, and palmitoleic acid. CPM, the combination of high-light efficiency pruning and mulching of C. oleifera trees; M, mulching of C. oleifera trees; UO, outside the upper canopy; UI, inside the upper canopy; LO, outside the lower canopy; LI, inside the lower canopy. One-way ANOVAs for M and CPM are displayed indicating. ** indicates significance at p ≤ 0.01.
3.5. Response of Yield and Quality to C. oleifera Tree Pruning
The effect of pruning on the fruit morphology index is shown in Table 1. Within the lower canopy, there was a significant difference in the fruit shape index in that fruits in CPM had a greater (p ≤ 0.05) longitudinal diameter and lateral diameter than those in M. The fresh fruit weight of the treated trees increased by 41.92%. However, there were no significant differences in the fruit shape index or fruit fresh weight in LO between CPM and M. These results indicated that the fruits inside the lower canopy had a more positive response to the treatment of the CPM treatment.
The results showed that the high-light efficiency pruning of C. oleifera trees strongly influenced yield and quality (Table 2 and Figure 5B–H). The fresh seed percentage of CPM was significantly greater (13.21%) in LI but was not significantly greater in UO, UI, or LO compared with M. The fruit yield per tree was greater in the CPM group than in the M group. At the time of harvest, the UO, UI, and LI of the CPM group had yields per tree of 2.52 kg, 1.41 kg, and 0.45 kg, respectively, which were 20%, 34.29%, and 36.36% greater than those of the M group. There was no significant difference in the seed water content or soluble sugar content at the four canopy positions between CPM and M. The soluble starch content significantly increased in the CPM group, especially in UI (4.04 mg/g under fresh weight conditions), which was 1.21 times greater than that in the M group.
Table 2.
Changes in fruit yield and fruit quality of C. oleifera ‘Huaxin’ in 2022.
The CPM of C. oleifera trees had different effects on the oil content ratio at different canopy positions (Table 2). Compared to that in the M treatment, the oil content ratio in the lower canopy was greater, but there was no significance or there was a decrease in the upper canopy in the CPM group. The major component was oleic acid, accounting for the largest proportion of the total fatty acid content, followed by linoleic acid and palmitic acid, while stearic acid, cis-11-eicosenoate acid, linolenic acid, and palmitoleic acid accounted for a very small proportion of the seed oil (Figure 5B–H). Erucic acid, which is harmful to human health, was not detected in any of the samples.
There was no significant difference (p ≤ 0.05) in the average percentage of oleic acid in the four canopy positions between the CPM group and the M group. Compared to those in the M group, the oleic acid concentrations in the CPM group were 79.85%, 79.95%, 80.25%, and 79.6% for UO, UI, LO, and LI, respectively, with no significant difference. Similar results were also found for the other six fatty acid components. These results indicated that the high-light efficiency pruning of C. oleifera trees did not affect the oil quality at the four canopy positions.
4. Discussion
4.1. Variation in the Canopy Microclimate throughout the Fruit Growing Season
The microclimate conditions of the canopy are influenced by the filtering of the environment by the leaves [38]. The redistribution of natural light intercepted by leaves in different canopy layers results in microclimate differences. Our results showed that temperature and humidity demonstrated no significant differences, but a significant difference was observed in light intensity among the four canopy positions (Figure 2). This was consistent with the conclusions of Wen et al. [31], Lu et al. [37]. The closer to the inner canopy position, the greater the humidity, the lower the temperature, which is inconsistent with the results of flavedo studies [39], Sapindus mukorossi Gaertn [30], and Xanthoceras sorbifolium [32] studies. This might be attributed to the small crown breadth of Camellia oleifera. The light intensity in the outside canopy position was significantly greater than that of the inside the canopy position due to the positional advantages. The highest light intensity was observed in UO, and the lowest light intensity was observed inside the lower canopy. These findings confirm the results obtained for Camellia oleifera [35], Xanthoceras sorbifolium Bunge [40], and peach [41].
4.2. The Impact of Canopy Position on the Fruit Development Characteristics
An exploration of the characteristics of fruit development is extremely important for guiding orchard management. The results of this study indicated that the fruits expanded rapidly from July to August, and fatty acids accumulated rapidly by mid-August, when the volume tended to stabilize in Hunan, China. This result provided further support for the view that seed oil accumulation occurred after the rapid expansion of the fruit [9]. Compared with the fruits from the inside canopy, the longitudinal diameter, lateral diameter, and single-fruit weight of fruits in the outside canopy position were greater due to the greater light availability (Figure 3). With the development of fruit, the accumulation of seed oil increases gradually and reaches a peak at maturity [9,10]. The current study revealed that kernel oil accumulation mainly occurred from August to October. Figure 3 shows an upward trend in the accumulation of total fatty acids in the kernels from inside the lower canopy position, whereas a downward trend was observed for the other three canopy positions near the harvest stage. Taken together, these findings indicated that the seed oil of the fruits in the other three canopy positions gradually degraded, and a suitable harvest time was missed; moreover, the fruits from inside the lower canopy position were still immature because of low light availability. Benelli et al. [42] reported that a canopy with 30% incident light of a tree not only reduced the mesocarp oil content but also delayed fruit maturity in a study of the effects of the light environment on the quality and maturity of olive plants. These results confirmed that irregular fruit maturation is associated with a poor light environment in the canopy. Previous studies have shown that appropriate drought conditions helped promote oil accumulation, but excessive drought threatened fatty acid synthesis and accumulation [43]. Remarkably, we also discovered that in September, the total fatty acid content of seeds in the upper canopy was much lower than that of seeds in the lower canopy; at this time, the total rainfall (only 10.9 mm) was seriously reduced by more than 90% compared to that in August (Supplemental Table S1). These results may have occurred because fruits in the upper canopy position were more sensitive to water deficit than those in the lower canopy position. Overall, the above results showed that uneven fruit development was related to the nonuniform distribution of light intensity, and the change in oil content in the upper canopy layer was more sensitive to environmental changes. Based on the results of this study of the relationship between the canopy microclimate and fruit development, we further carried out canopy manipulations and mulch application, which might have helped to ensure the uniformity of the canopy light distribution and alleviated water deficit during the oil accumulation stage.
4.3. High-Light Efficiency Pruning Regulated the Canopy Microclimate
Pruning is a widely used technique in fruit crops to improve the tree structure, optimize sunlight utilization, and maximize the economic yield and fruit quality [44,45,46]. Pruning mainly changes the distribution of light in the canopy and further affects the temperature and relative humidity [41,47]. In this study, we found that the light intensity and temperature were greater while the relative humidity was lower in the canopies of pruned than in that of unpruned trees. The light intensity inside the canopy showed the greatest increase. This finding is also in accordance with earlier observations, which showed that an open-center canopy resulted in more light at every canopy position than a round-headed canopy [35]. A comparison of these findings with those of a previous study confirmed that the canopy of pruned fruit trees had higher relative humidity and lower temperature [29]. A possible explanation for this difference might be the sparser leaf distribution and greater light interception of trees subjected to pruning, which eliminated overcrowded and crossed branches from inside the canopy.
4.4. High-Light Efficiency Pruning of C. oleifera Trees Improved Fruit Quality and Maturity Uniformity at Harvest
Pruning resulted in differences in light intensity, temperature, and relative humidity. These differences may further affect fruit development, quality, and substance metabolism [44,46]. In this study, the fruit fresh weight and yield were greater in the CPM group than in the M group, except for those in the outside lower canopy position, which was consistent with prior research findings [35,42]. The oil content ratio of seeds from lower canopy fruits was substantially greater in the CPM than in M. These findings are consistent with those of Wen, Zhang, Su, Yang, Ma, Zhang, and Wang [35], who reported a greater oil content in an open-center canopy due to a better light distribution than in a round-headed canopy. There were no differences in fatty acid composition between the CPM and M treatments. These findings indicated the strong impact of light intensity on the oil yield parameters [30]. We found a markedly greater soluble starch content, as expected for LO, whereas there were no significant differences in the seed water content or soluble sugar content in the CPM group than the M group. A previous study on Castanea henryi showed that under medium and heavy pruning, light distribution was negatively correlated with the fruit water content and positively correlated with the soluble sugar and starch contents [46]. A possible explanation for this might be that light pruning slightly affected the water supply of the tree because the water deficit of the tree was satisfied after mulching. In our study on ‘Huaxin’, there was a greater fruit cracking rate during the CPM treatment compared to the M treatment, especially in UI and LO, due to the greater amount of lighting radiation, which indicated a significant effect of the light distribution in the tree canopy on maturation. This finding is consistent with the results of olive trees in this area linking the canopy microclimate and maturation indices [42]. Importantly, the difference in the fruit cracking rate among the four canopy positions in CPM treatment was not significant, indicating that the fruit matured more uniformly at harvest. Our results demonstrated that an open center is the ideal design for C. oleifera production because of the more equal distribution of the microclimatic variables and more consistent fruit quality parameters [35].
The results showed the importance of pruning and mulching for resisting drought, fruit maturation, and fruit quality in adult camellia forests. We are also aware of the limitations of this study. For example, does the interaction between the light environment and water status affect fruit quality and maturation? What are the light intensity and soil water content thresholds for fruit development? The effects of the canopy microclimate on fatty acid metabolism, the key genes involved in fatty acid synthesis, and the regulatory elements that drive differences in fatty acid metabolism difference will be reported in the next study.
5. Conclusions
The faster oil accumulation from the outside canopy, which had a higher light intensity, suggested earlier maturation compared with fruit harvested from inside the lower canopy. The crucial stage of fatty acid synthesis and accumulation differences occurred in mid-September due to the uneven distribution of the canopy microclimate. The high-light efficiency pruning of C. oleifera trees significantly increased the fruit fresh weight, yield, soluble starch content, and oil content in the kernel from the inside lower canopy position due to increased light radiation compared with unpruned trees. Uneven maturation, high yield, and high seed quality were achieved by pruning and mulching with camellia shells and weedproof film. These findings provide new insights for improving the harvesting efficiency.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/horticulturae10040343/s1, Table S1: Climate change in experimental field in 2021–2022.
Author Contributions
Conceptualization, S.S. and Q.B.; methodology, Y.S. (Yongjiang Sun); validation, S.S.; formal analysis, Q.B.; resources, S.S.; data curation, Y.L. and Y.S. (Yuanyuan Si); writing—original draft preparation, Y.S. (Yuanyuan Si); writing—review and editing, Y.S. (Yuanyuan Si); project administration, S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Jiangxi Forestry of Science and Technology Innovation project ‘Study on Drought-resistant Cultivation Techniques of Camellia oleifera in Jiangxi’ [Innovation project 2022, No. 37].
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
The authors are grateful to Taishan Chen of the private farm for the assistance provided during the in-field trial. The authors are grateful to Zhuzhou Meteorological Administration, Hunan, China, for the assistance provided with the local meteorological data.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhu, G.; Liu, H.; Xie, Y.; Liao, Q.; Lin, Y.; Liu, Y.; Liu, Y.; Xiao, H.; Gao, Z.; Hu, S. Postharvest processing and storage methods for Camellia oleifera seeds. Food Rev. Int. 2019, 36, 319–339. [Google Scholar] [CrossRef]
- Cheng, X.X.; Yang, T.; Wang, Y.H.; Zhou, B.Q.; Yan, L.; Teng, L.Z.; Wang, F.B.; Chen, L.L.; He, Y.; Guo, K.P.; et al. New method for effective identification of adulterated Camellia oil basing on Camellia oleifera-specific DNA. Arab. J. Chem. 2018, 11, 815–826. [Google Scholar] [CrossRef]
- Luan, F.; Zeng, J.; Yang, Y.; He, X.; Wang, B.; Gao, Y.; Zeng, N. Recent advances in Camellia oleifera Abel: A review of nutritional constituents, biofunctional properties, and potential industrial applications. J. Funct. Foods 2020, 75, 104242. [Google Scholar] [CrossRef]
- Ma, J.L.; Ye, H.; Rui, Y.K.; Chen, G.C.; Zhang, N.Y. Fatty acid composition of Camellia oleifera oil. J. Für Verbraucherschutz Und Leb. 2010, 6, 9–12. [Google Scholar] [CrossRef]
- Zhou, D.Y.; Liu, Y.X.; Xu, Z.L.; Yin, F.W.; Song, L.; Wan, X.L.; Song, Y.K.; Zhu, B.W. Effects of long-term intake of Antarctic krill oils on artery blood pressure in spontaneously hypertensive rats. J. Sci. Food Agric. 2017, 97, 1143–1148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.K.; Hu, M.J.; Yun, Z.; Wang, J.B.; Feng, G.; Gao, Z.Y.; Shi, X.Q.; Jiang, Y.M. Effect of tea seed oil treatment on browning of litchi fruit in relation to energy status and metabolism. Postharvest Biol. Technol. 2017, 132, 97–104. [Google Scholar] [CrossRef]
- Chou, T.Y.; Lu, Y.F.; Inbaraj, B.S.; Chen, B.H. Camelia oil and soybean-camelia oil blend enhance antioxidant activity and cardiovascular protection in hamsters. Nutrition 2018, 51–52, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Guo, Y.; Wu, P.; Lu, F.; Zhu, J.; Ma, H.; Chen, Y.; Zhang, T. Camellia oil lowering blood pressure in spontaneously hypertension rats. J. Funct. Foods 2020, 70, 103915. [Google Scholar] [CrossRef]
- Cao, Y.Q.; Yao, X.H.; Ren, H.D.; Wang, K.L. Changes in contents of endogenous hormones and main mineral elements in oil-tea camellia fruit during maturation. J. Beijing For. Univ. 2015, 37, 76–81. [Google Scholar] [CrossRef]
- Song, Q.; Ji, K.; Mo, W.; Wang, L.; Chen, L.; Gao, L.; Gong, W.; Yuan, D. Dynamics of sugars, endogenous hormones, and oil content during the development of Camellia oleifera fruit. Botany 2021, 99, 515–529. [Google Scholar] [CrossRef]
- Zhang, F.; Li, Z.; Zhou, J.; Gu, Y.; Tan, X. Comparative study on fruit development and oil synthesis in two cultivars of Camellia oleifera. BMC Plant Biol. 2021, 21, 348. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.P.; Lei, T.W.; Mao, L.L.; Liu, H.; Wu, Y. Soil surface evaporation processes under mulches of different sized gravel. Catena 2009, 78, 117–121. [Google Scholar] [CrossRef]
- Xie, Z.K.; Wang, Y.J.; Jiang, W.L.; Wei, X.H. Evaporation and evapotranspiration in a watermelon field mulched with gravel of different sizes in northwest China. Agric. Water Manag. 2006, 81, 173–184. [Google Scholar] [CrossRef]
- Ye, H.L.; Chen, Z.G.; Jia, T.T.; Su, Q.W.; Su, S.C. Response of different organic mulch treatments on yield and quality of Camellia oleifera. Agric. Water Manag. 2021, 245, 106654. [Google Scholar] [CrossRef]
- Bai, H.S.; Blumfield, T.J.; Reverchon, F. The impact of mulch type on soil organic carbon and nitrogen pools in a sloping site. Biol. Fertil. Soils 2013, 50, 37–44. [Google Scholar] [CrossRef]
- Luo, J.; Xu, M.; Liu, C.; Wei, S.; Tang, H. Effects comparation of different mulching methods on soil in pitaya orchards. Int. Agrophysics 2021, 35, 267–276. [Google Scholar] [CrossRef]
- Waheed, A.; Li, C.; Muhammad, M.; Ahmad, M.; Khan, K.A.; Ghramh, H.A.; Wang, Z.; Zhang, D. Sustainable potato growth under straw mulching practices. Sustainability 2023, 15, 442. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, R.Y.; Ma, B.L.; Xiong, Y.C.; Qiang, S.C.; Wang, C.L.; Liu, C.A.; Li, F.M. Ridge-furrow with full plastic film mulching improves water use efficiency and tuber yields of potato in a semiarid rainfed ecosystem. Field Crops Res. 2014, 161, 137–148. [Google Scholar] [CrossRef]
- Li, X.B. The Effect of Mulching on Soil Temperature, Winter potato (Solanum Tuberosum L.) growth and yield in field experiment, South China. Appl. Ecol. Environ. Res. 2018, 16, 913–929. [Google Scholar] [CrossRef]
- Goel, L.; Shenker, V.; Sharma, R.K. Effect of organic mulches on agronomic parameters—A case study of tomato crop (Mill.). Int. J. Recycl. Org. 2020, 9, 297–307. [Google Scholar] [CrossRef]
- López-Urrea, R.; Sánchez, J.M.; Montoro, A.; Mañas, F.; Intrigliolo, D.S. Effect of using pruning waste as an organic mulching on a drip-irrigated vineyard evapotranspiration under a semi-arid climate. Agric. For. Meteorol. 2020, 291, 108064. [Google Scholar] [CrossRef]
- Chan, K.Y.; Fahey, D.J.; Newell, M.; Barchia, I. Using composted mulch in vineyards—Effects on grape yield and quality. Int. J. Fruit Sci. 2010, 10, 441–453. [Google Scholar] [CrossRef]
- Chen, H.X.; Liu, J.J.; Zhang, A.F.; Chen, J.; Cheng, G.; Sun, B.H.; Pi, X.M.; Dyck, M.; Si, B.C.; Zhao, Y.; et al. Effects of straw and plastic film mulching on greenhouse gas emissions in Loess Plateau, China: A field study of 2 consecutive wheat-maize rotation cycles. Sci. Total Environ. 2017, 579, 814–824. [Google Scholar] [CrossRef] [PubMed]
- Kader, M.A.; Nakamura, K.; Senge, M.; Mojid, M.A.; Kawashima, S. Soil hydro-thermal regimes and water use efficiency of rain-fed soybean (Glycine max) as affected by organic mulches. Agric. Water Manag. 2019, 223, 105707. [Google Scholar] [CrossRef]
- Adiga, D.J.; Veena, G.L.; Thondaiman, V.; Babli, M. An overview of canopy management in cashew (Anacardium occidentale L.). J. Hortic. Sci.-India 2020, 15, 127–135. [Google Scholar] [CrossRef]
- Hipps, N.A.; Davies, M.J.; Dunn, J.M.; Griffiths, H.; Atkinson, C.J. Effects of two contrasting canopy manipulations on growth and water use of London plane (Platanus × acerifolia) trees. Plant Soil 2014, 382, 61–74. [Google Scholar] [CrossRef]
- Rutter, N.; Essery, R.; Baxter, R.; Hancock, S.; Horton, M.; Huntley, B.; Reid, T.; Woodward, J. Canopy structure and air temperature inversions impact simulation of sub-canopy longwave radiation in snow-covered boreal forests. J. Geophys. Res. Atmos. 2023, 128, e2022JD037980. [Google Scholar] [CrossRef]
- Song, Q.F.; Chu, C.C.; Parry, M.A.; Zhu, X.G. Genetics-based dynamic systems model of canopy photosynthesis: The key to improve light and resource use efficiencies for crops. Food Energy Secur. 2016, 5, 18–25. [Google Scholar] [CrossRef]
- Sharma, R.R.; Singh, R. Pruning intensity modifies canopy microclimate, and influences sex ratio, malformation incidence and development of fruited panicles in ‘Amrapali’ mango (Mangifera indica L.). Sci. Hortic. 2006, 109, 118–122. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Wen, Y.; Bai, Q.; Ma, Z.; Ye, H.L.; Su, S.C. Spatio-temporal effects of canopy microclimate on fruit yield and quality of Sapindus mukorossi Gaertn. Sci. Hortic. 2019, 251, 136–149. [Google Scholar] [CrossRef]
- Wen, Y.; Su, S.C.; Ma, L.Y.; Yang, S.Y.; Wang, Y.W.; Wang, X.N. Effects of canopy microclimate on fruit yield and quality of Camellia oleifera. Sci. Hortic. 2018, 235, 132–141. [Google Scholar] [CrossRef]
- Ou, L.J.; Zhang, Y.; Zhang, Z.S.; Chen, Y.X.; Wang, K.X.; Wen, Y.; Ao, Y. The relationship between canopy microclimate, fruit and seed yield, and quality in Xanthoceras sorbifolium. J. Plant Physiol. 2023, 284, 153975. [Google Scholar] [CrossRef] [PubMed]
- Li, K.T.; Lakso, A.N.; Piccioni, R.; Robinson, T. Summer pruning reduces whole-canopy carbon fixation and transpiration in apple trees. J. Hortic. Sci. Biotechnol. 2003, 78, 749–754. [Google Scholar] [CrossRef]
- Lodolini, E.M.; Polverigiani, S.; Giorgi, V.; Famiani, F.; Neri, D. Time and type of pruning affect tree growth and yield in high-density olive orchards. Sci. Hortic. 2023, 311, 111831. [Google Scholar] [CrossRef]
- Wen, Y.; Zhang, Y.; Su, S.; Yang, S.; Ma, L.; Zhang, L.; Wang, X. Effects of tree shape on the microclimate and fruit quality parameters of Camellia oleifera Abel. Forests 2019, 10, 563. [Google Scholar] [CrossRef]
- Ye, C.Q.; Li, Y.; Liu, Y.; Li, C.; Qin, C.; Su, S.C. Effects of shortcut strength on the growth of fruiting and developing branches of Chinese chestnut (Castanea mollissima). Non-Wood For. Res. 2020, 38, 184–191. [Google Scholar] [CrossRef]
- Lu, Y.; Si, Y.; Zhang, L.; Sun, Y.; Su, S. Effects of canopy position and microclimate on fruit development and quality of Camellia oleifera. Agronomy 2022, 12, 2158. [Google Scholar] [CrossRef]
- Zhang, D.P. Basic concepts and implications in the study of canopy microclimate and its regulatory biology. Grape Cultiv. Winemak. 1993, 1–4. [Google Scholar] [CrossRef]
- Cronje, P.J.R.; Barry, G.H.; Huysamer, M. Fruiting position during development of ‘Nules Clementine’ mandarin affects the concentration of K, Mg and Ca in the flavedo. Sci. Hortic. 2011, 130, 829–837. [Google Scholar] [CrossRef]
- Zhao, X.; Rong, G.; Wang, Y.; Duan, J.; Ma, L. Relationships between branch growth and leaf canopy microclimate of Xanthoceras sorbifolium Bunge. J. Northwest A F Univ. (Nat. Sci. Ed.) 2020, 48, 44–56. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, W.; Jia, H.; Li, S.; Yang, L.; Wang, H. Effects of light distribution in canopy of peach trees with different tree shapes on fruit yield and quality. North. Hortic. 2024. [Google Scholar] [CrossRef]
- Benelli, G.; Caruso, G.; Giunti, G.; Cuzzola, A.; Saba, A.; Raffaelli, A.; Gucci, R. Changes in olive oil volatile organic compounds induced by water status and light environment in canopies of Olea europaea L. trees. J. Sci. Food Agric. 2015, 95, 2473–2481. [Google Scholar] [CrossRef] [PubMed]
- Hamrounia, I.; Salahb, H.B.; Marzouk, B. Effects of water-deficit on lipids of safflower aerial parts. Phytochemistry 2001, 58, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Li, K.T.; Lakso, A.N.; Piccioni, R.; Robinson, T. Summer pruning effects on fruit size, fruit quality, return bloom and fine root survival in apple trees. J. Hortic. Sci. Biotechnol. 2003, 78, 755–761. [Google Scholar] [CrossRef]
- Wang, X.; De Bei, R.; Fuentes, S.; Collins, C. Influence of canopy management practices on canopy architecture and reproductive performance of semillon and shiraz grapevines in a hot climate. Am. J. Enol. Vitic. 2019, 70, 360–372. [Google Scholar] [CrossRef]
- Wang, G.; Yuan, D.Y.; Zou, F.; Xiong, H.; Zhu, Z.J.; Liu, Z.Q.; Ouyang, F. Effect of different pruning intensity on the canopy light distribution and yield and quality in Castanea henryi. J. Fruit Sci. 2017, 34, 329–336. [Google Scholar] [CrossRef]
- Gao, Y.; Jia, L.M.; Gao, S.L.; Su, S.C.; Duan, J.; Weng, Z. Reasonable canopy light intensity and high light efficiency regulation of Sapindus mukorossi. Sci. Silvae Sin. 2016, 52, 29–38. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).