
Re-Sequencing the Mitochondrial Genome Unveils a Novel Isomeric Form of NWB CMS Line in Radish and Functional Verification of Its Candidate Sterile Gene

 and
and
Abstract
:1. Introduction
2. Results
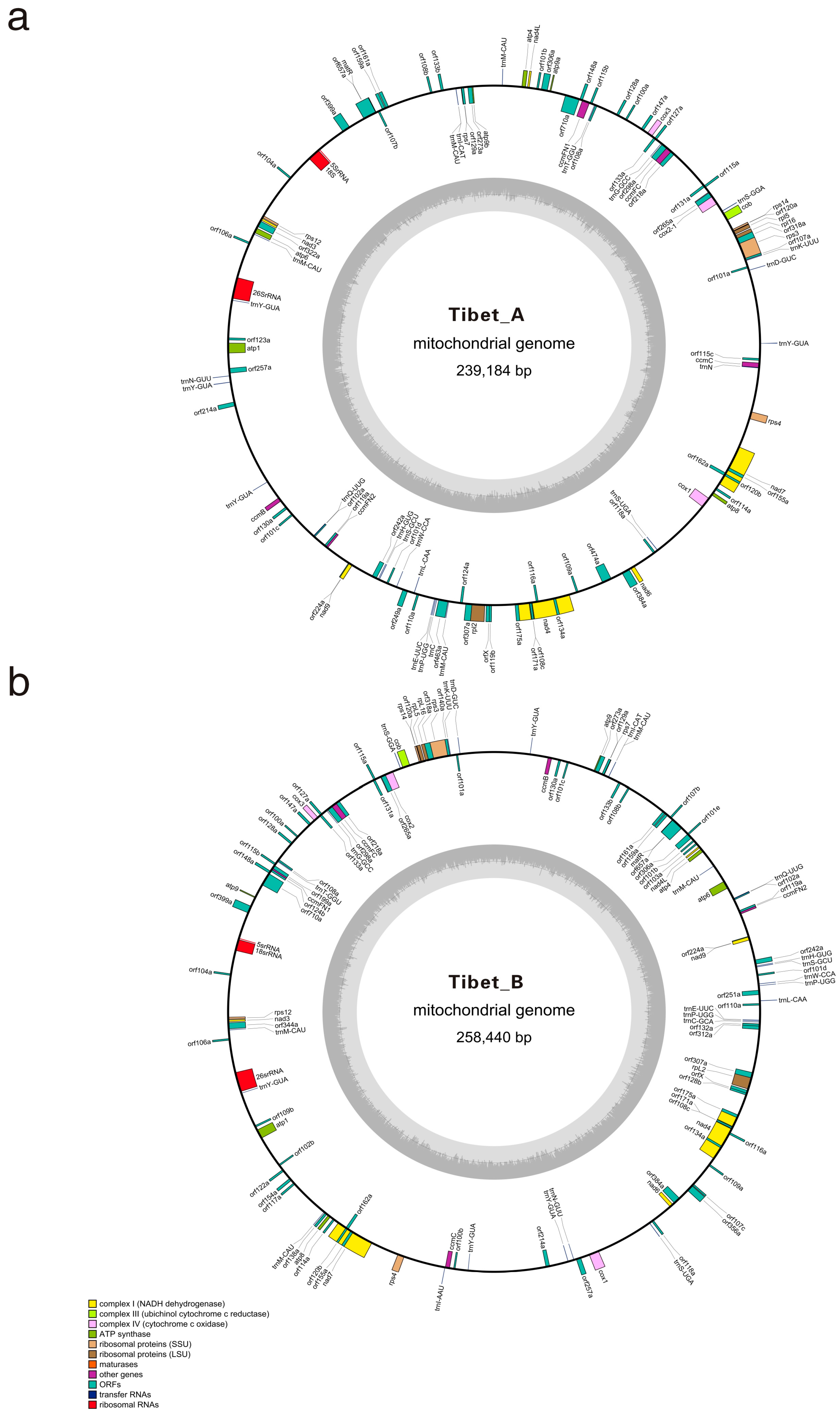
2.1. Mitochondrial Genomes Composition of Tibet A and Tibet B
2.2. Screening for Genes Responsible for Male Sterility in Tibet A Mitogenome
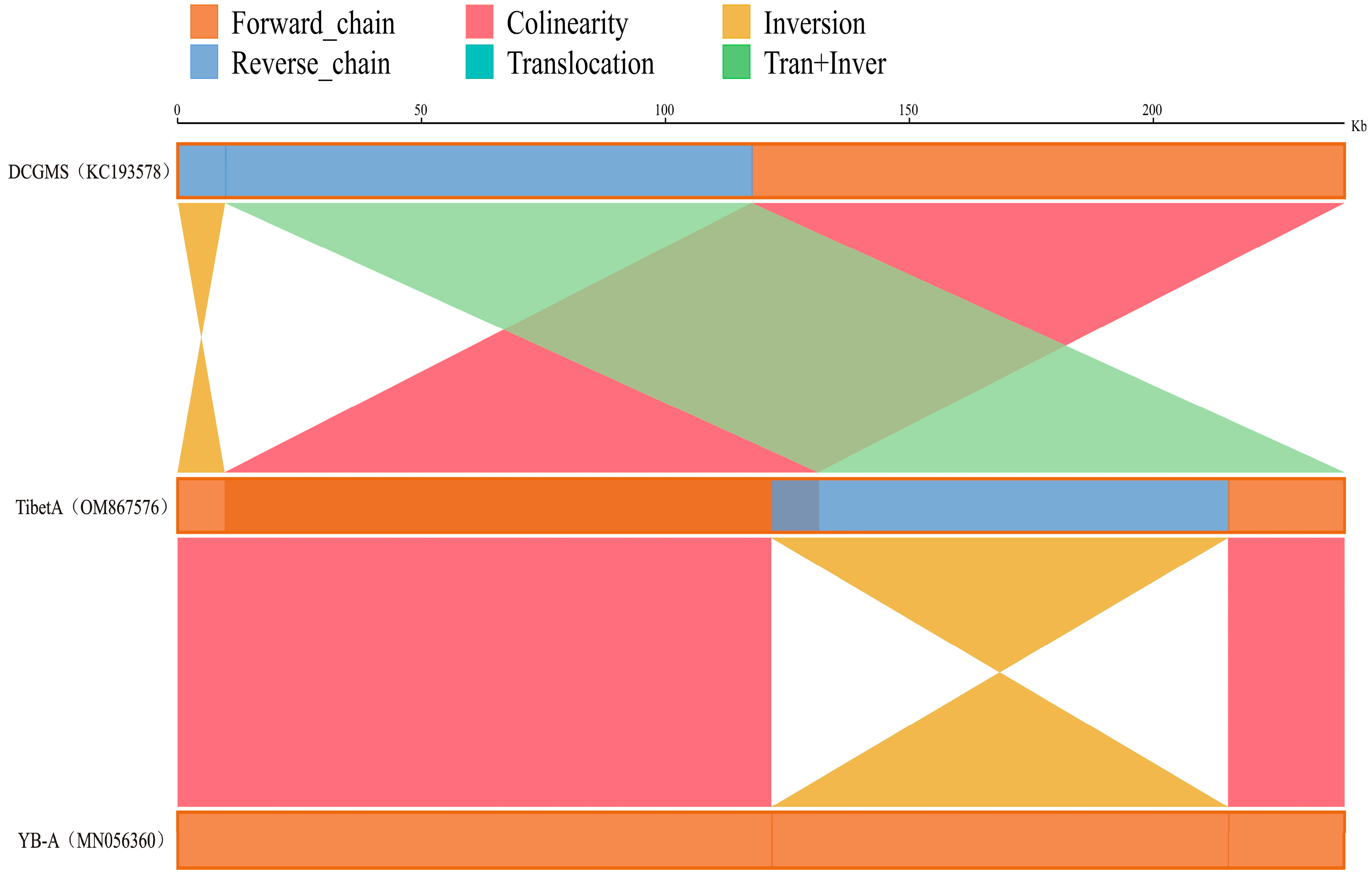
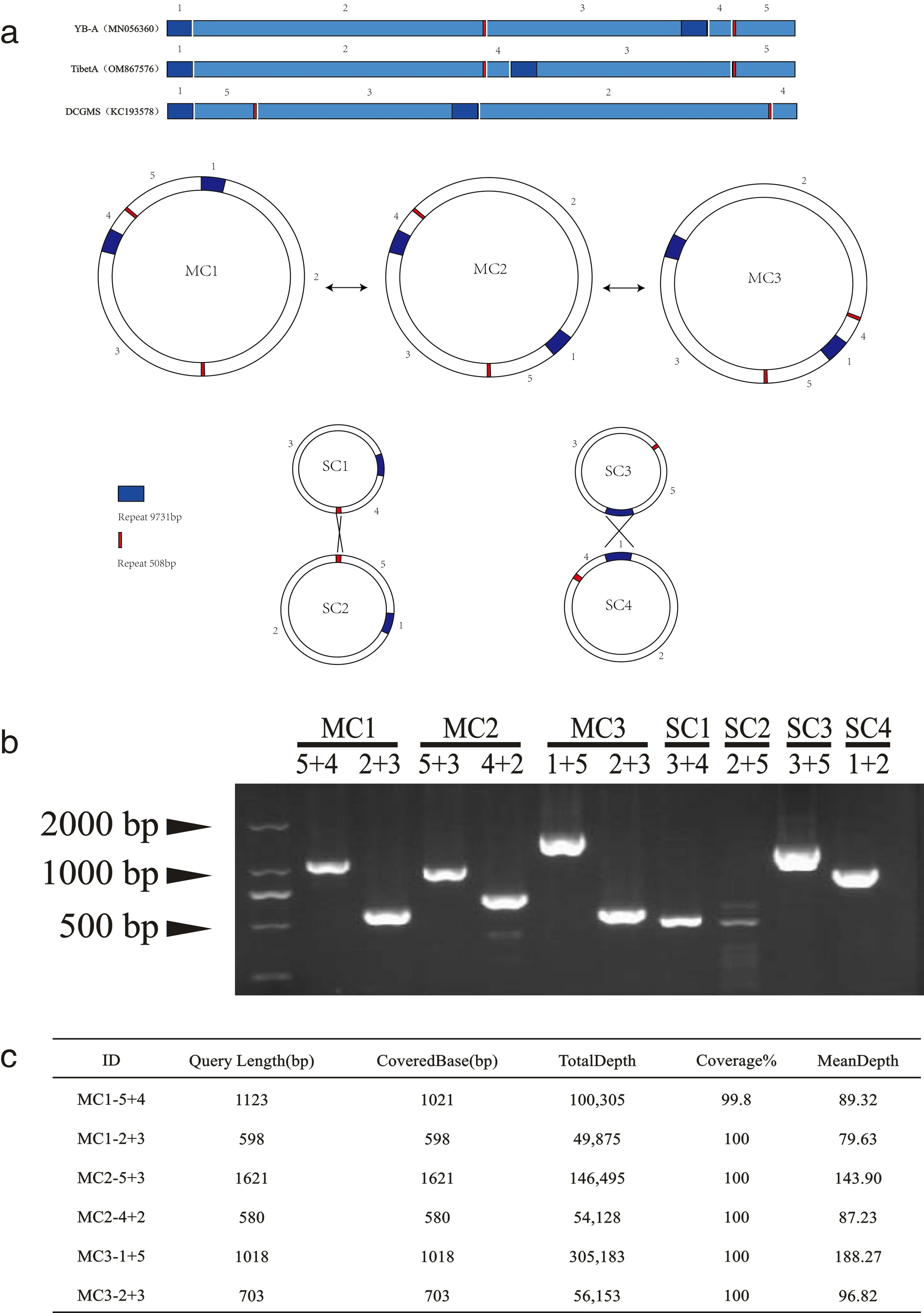
2.3. Explores the Relationship among the Three Mitogenomes of Tibet A, NWB YB-A, and DCGMS
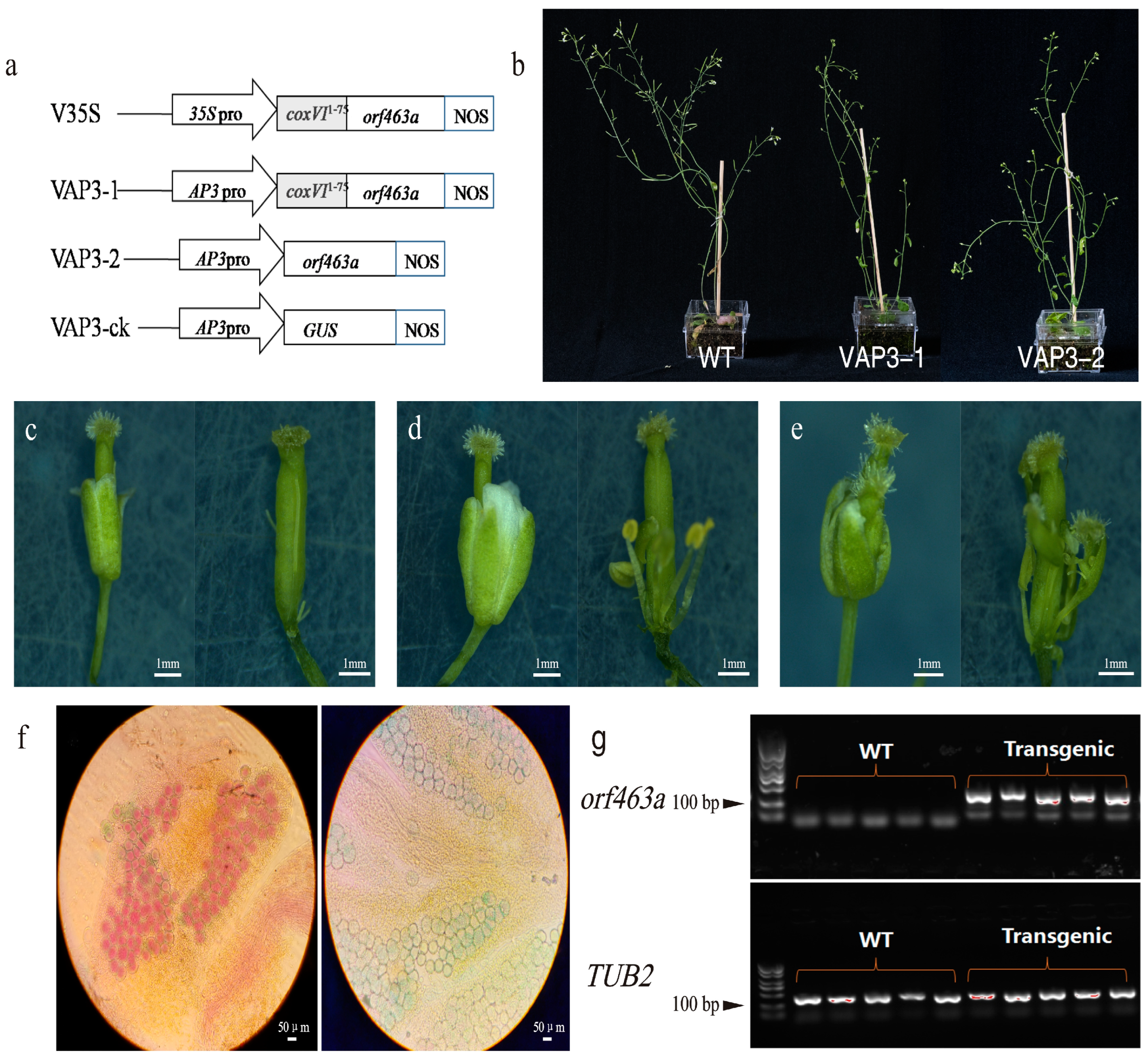
2.4. Expression of orf463a in Arabidopsis Severely Affects the Development of Floral Organs
3. Discussion
3.1. The Mitochondrial Genomes of Tibet A and NWB CMS YB-A and DCGMS Are Coexisting Isoforms of the Same Mitotype
3.2. Genetic Transformation Confirms That orf463a Is the Functional Gene Controlling Pollen Abortion in NWB CMS
4. Materials and Methods
4.1. Plant Materials
4.2. Mitochondrial DNA Extraction, Sequence, and Genome Assembly
4.3. Gene Annotation
4.4. Prediction of Transmembrane Domains and Comparative Analysis of Radish Mitochondrial Genomes
4.5. Detection of Mitochondrial Genome Isoforms
4.6. Vector Construction and Genetic Transformation of Arabidopsis thaliana
4.7. RNA Extraction and RT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Laser, K.D.; Lersten, N.R. Anatomy and cytology of microsporogenesis in cytoplasmic male sterile angiosperms. Bot. Rev. 1972, 38, 425–454. [Google Scholar] [CrossRef]
- Levings, C.S.; Pring, D.R. Restriction endonuclease analysis of mitochondrial DNA from normal and Texas cytoplasmic male-sterile maize. Science 1976, 193, 158–160. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.R.; Bentolila, S. Interactions of mitochondrial and nuclear genes that affect male gametophyte development. Plant Cell 2004, 16, S154–S169. [Google Scholar] [CrossRef] [PubMed]
- Horn, R.; Gupta, K.J.; Colombo, N. Mitochondrion role in molecular basis of cytoplasmic male sterility. Mitochondrion 2014, 19, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Bohra, A.; Jha, U.C.; Adhimoolam, P.; Bisht, P.; Singh, N.P. Cytoplasmic male sterility (CMS) in hybrid breeding in field crops. Plant Cell Rep. 2016, 35, 967–993. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.W.; Xie, Y.Y.; Liu, Y.G.; Chen, L.T. Advances in understanding the molecular mechanisms of cytoplasmic male sterility and restoration in rice. Plant Reprod. 2017, 30, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Zhang, D.B. Molecular control of male fertility for hybrid breeding. Trends Plant Sci. 2018, 23, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Dey, S.S.; Bhatia, R.; Kumar, R.; Behera, T.K. Current understanding of male sterility systems in vegetable Brassicas and their exploitation in hybrid breeding. Plant Reprod. 2019, 32, 231–256. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.Y.; Yang, X.D.; Zhao, N.; Hu, Z.Y.; Mackenzie, S.A.; Zhang, M.Y.; Yang, J.H. Exploiting sterility and fertility variation in cytoplasmic male sterile vegetable crops. Hortic. Res. 2022, 9, uhab039. [Google Scholar] [CrossRef]
- Dewey, R.E.; Timothy, D.H.; Levings, C.S. A mitochondrial protein associated with cytoplasmic male sterility in the T cytoplasm of maize. Proc. Natl. Acad. Sci. USA 1987, 84, 5374–5378. [Google Scholar] [CrossRef]
- Song, J.; Hedgcoth, C. A chimeric gene (orf256) is expressed as protein only in cytoplasmic male-sterile lines of wheat. Plant Mol. Biol. 1994, 26, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.P.; Shinada, H.; Onodera, Y.; Komaki, C.; Mikami, T.; Kubo, T. A male sterility-associated mitochondrial protein in wild beets causes pollen disruption in transgenic plants. Plant J. 2008, 54, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Yi, P.; Wang, L.; Sun, Q.; Zhu, Y.G. Discovery of mitochondrial chimeric-gene associated with cytoplasmic male sterility of HL-rice. Chin. Sci. Bull. 2002, 47, 744–747. [Google Scholar] [CrossRef]
- Igarashi, K.; Kazama, T.; Motomura, K.; Toriyama, K. Whole genomic sequencing of RT98 mitochondria derived from Oryza rufipogon and northern blot analysis to uncover a cytoplasmic male sterility-associated gene. Plant Cell Physiol. 2013, 54, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Brown, G.G. Suppression of cytoplasmic male sterility by nuclear genes alters expression of a novel mitochondrial gene region. Plant Cell 1991, 3, 1349–1362. [Google Scholar] [PubMed]
- Okazaki, M.; Kazama, T.; Murata, H.; Motomura, K.; Toriyama, K. Whole mitochondrial genome sequencing and transcriptional analysis to uncover an RT102-type cytoplasmic male sterility-associated candidate gene derived from Oryza rufipogon. Plant Cell Physiol. 2013, 54, 1560–1568. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.D.; Choi, Y.; Kim, D.H.; Kim, B.D.; Kang, B.C. Extensive structural variations between mitochondrial genomes of CMS and normal peppers (Capsicum annuum L.) revealed by complete nucleotide sequencing. BMC Genom. 2014, 15, 561. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.F.; Mei, D.S.; Liu, J.; Zaman, Q.U.; Zhang, H.Y.; Hao, M.Y.; Fu, L.; Wang, H.; Cheng, H.T.; Hu, Q. Organelle genome composition and candidate gene identification for Nsa cytoplasmic male sterility in Brassica napus. BMC Genom. 2019, 20, 813. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Yang, T.J.; Kim, S. Identification of a gene responsible for cytoplasmic male-sterility in onions (Allium cepa L.) using comparative analysis of mitochondrial genome sequences of two recently diverged cytoplasms. Theor. Appl. Genet. 2019; 132, 313–322. [Google Scholar]
- Zhang, S.; Yin, Z.P.; Wu, X.M.; Li, C.C.; Xie, K.D.; Deng, X.X.; Grosser, J.W.; Guo, W.W. Assembly of Satsuma mandarin mitochondrial genome and identification of cytoplasmic male sterility-specific ORFs in a somatic cybrid of pummelo. Tree Genet. Genomes 2020, 16, 84. [Google Scholar] [CrossRef]
- Arun, V.; Kuriakose, B.; Sridhar, V.V.; Thomas, G. Transformation and analysis of tobacco plant var Petit havana with T-urf13 gene under anther-specific TA29 promoter. Biotech 2011, 1, 73–82. [Google Scholar] [CrossRef]
- Bhatnagar-Mathur, P.; Gupta, R.; Reddy, P.S.; Reddy, B.P.; Reddy, D.S.; Sameerkumar, C.V.; Saxena, R.K.; Sharma, K.K. A novel mitochondrial orf147 causes cytoplasmic male sterility in pigeonpea by modulating aberrant anther dehiscence. Plant Mol. Biol. 2018, 97, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Heng, S.; Gao, J.; Wei, C.; Chen, F.; Li, X.; Wen, J.; Yi, B.; Ma, C.; Tu, J.; Fu, T.; et al. Transcript levels of orf288 are associated with the hau cytoplasmic male sterility system and altered nuclear gene expression in Brassica juncea. J. Exp. Bot. 2018, 69, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Kazama, T.; Okuno, M.; Watari, Y.; Yanase, S.; Koizuka, C.; Tsuruta, Y.; Sugaya, H.; Toyoda, A.; Itoh, T.; Tsutsumi, N.; et al. Curing cytoplasmic male sterility via TALEN-mediated mitochondrial genome editing. Nat. Plants 2019, 5, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, X.; Su, T.; Hu, Z.; Zhang, M. The development of mitochondrial gene editing tools and their possible roles in crop improvement for future agriculture. Adv. Genet. 2021, 3, 2100019. [Google Scholar] [CrossRef] [PubMed]
- Arimura, S.I.; Nakazato, I. Genome editing of plant mitochondrial and chloroplast genomes. Plant Cell Physiol. 2023, 19, pcad162. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.W.; Hatakeyama, K.; Takahata, Y. The use of genome information for intergeneric hybridization breeding. In The Radish Genome; Nishi, T., Kitashiba, H., Eds.; Springer Press: Cham, Switzerland, 2017; pp. 189–198. [Google Scholar]
- Ogura, H. Studies on the new male sterility in Japanese radish, with special reference to the utilization of this sterility towards the practical raising of hybrid seeds. Mem. Fac. Agric. Kagoshima Univ. 1968, 6, 39–78. [Google Scholar]
- Chen, L.; Liu, Y.G. Male sterility and fertility restoration in crops. Annu. Rev. Plant Biol. 2014, 65, 579–606. [Google Scholar] [CrossRef] [PubMed]
- Bonhomme, S.; Budar, F.; Férault, M.; Pelletier, G. A 2.5 kb NcoI fragment of Ogura radish mitochondrial DNA is correlated with cytoplasmic male sterility in Brassica cybrids. Curr. Genet. 1991, 19, 121–127. [Google Scholar] [CrossRef]
- Bonhomme, S.; Budar, F.; Lancelin, D.; Small, I.; Defrance, M.C.; Pelletier, G. Sequence and transcript analysis of the Nco2.5 Ogura-specific fragment correlated with cytoplasmic male sterility in Brassica cybrids. Mol. Gen. Genet. 1992; 235, 340–348. [Google Scholar]
- Grelon, M.; Budar, F.; Bonhomme, S.; Pelletier, G. Ogura cytoplasmic male-sterility (CMS)-associated orf138 is translated into a mitochondrial membrane polypeptide in male-sterile Brassica cybrids. Mol. Gen. Genet. 1994, 243, 540–547. [Google Scholar] [CrossRef]
- Duroc, Y.; Gaillard, C.; Hiard, S.; Defrance, M.C.; Pelletier, G.; Budar, F. Biochemical and functional characterization of ORF138, a mitochondrial protein responsible for Ogura cytoplasmic male sterility in Brassiceae. Biochimie 2005, 87, 1089–1100. [Google Scholar] [CrossRef]
- Tanaka, Y.; Tsuda, M.; Yasumoto, K.; Yamagishi, H.; Terachi, T. A complete mitochondrial genome sequence of Ogura-type male-sterile cytoplasm and its comparative analysis with that of normal cytoplasm in radish (Raphanus sativus L.). BMC Genom. 2012, 13, 352. [Google Scholar] [CrossRef] [PubMed]
- Nahm, S.H.; Lee, H.J.; Lee, S.W.; Joo, G.Y.; Harn, C.H.; Yang, S.G.; Min, B.W. Development of a molecular marker specific to a novel CMS line in radish (Raphanus sativus L.). Theor. Appl. Genet. 2005; 111, 1191–1200. [Google Scholar]
- Lee, Y.P.; Park, S.; Lim, C.; Kim, H.; Lim, H.; Ahn, Y.; Sung, S.K.; Yoon, M.K.; Kim, S. Discovery of a novel cytoplasmic male-sterility and its restorer lines in radish (Raphanus sativus L.). Theor. Appl. Genet. 2008; 117, 905–913. [Google Scholar]
- Park, J.Y.; Lee, Y.P.; Lee, J.; Choi, B.S.; Kim, S.; Yang, T.J. Complete mitochondrial genome sequence and identification of a candidate gene responsible for cytoplasmic male sterility in radish (Raphanus sativus L.) containing DCGMS cytoplasm. Theor. Appl. Genet. 2013; 126, 1763–1774. [Google Scholar]
- Yamagishi, H.; Tanaka, Y.; Shiiba, S.; Hashimoto, A.; Fukunaga, A. Mitochondrial orf463 causing male sterility in radish is possessed by cultivars belonging to the ‘Niger’ group. Euphytica 2019, 215, 109. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Hao, W.; Li, J.; Qi, M.; Zhang, L. Mitochondrial genome sequencing reveals orf463a may induce male sterility in NWB cytoplasm of radish. Genes 2020, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, H.; Hashimoto, A.; Fukunaga, A.; Terachi, T. Appearance of male sterile and black radishes in the progeny of cross between Raphanus raphanistrum and Raphanus sativus. Breed Sci. 2020, 70, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, H.; Terachi, T. Cytoplasmic male sterility and mitochondrial genome variations in radish. In The Radish Genome; Nishi, T., Kitashiba, H., Eds.; Springer Press: Cham, Switzerland, 2017; pp. 151–163. [Google Scholar]
- Yamagishi, H.; Terachi, T. Intra- and inter-specific variations in the mitochondrial gene orf138 of Ogura-type male-sterile cytoplasm from Raphanus sativus and Raphaus raphanistrum. Theor. Appl. Genet. 2001, 103, 725–732. [Google Scholar] [CrossRef]
- Knoop, V. Seed Plant Mitochondrial Genomes. In Complexity Evolving in Genomics of Chloroplasts and Mitochondria; Bock, R., Knoop, V., Eds.; Springer Press: Dordrecht, The Netherlands, 2012; pp. 175–200. [Google Scholar]
- Bellaoui, M.; Martin-Canadell, A.; Pelletier, G.; Budar, F. Low-copy-number molecules are produced by recombination, actively maintained and can be amplified in the mitochondrial genome of Brassicaceae: Relationship to reversion of the male sterile phenotype in some cybrids. Mol. Gen. Genet. 1998, 257, 177–185. [Google Scholar] [CrossRef]
- Arrieta-Montiel, M.; Lyznik, A.; Woloszynska, M.; Janska, H.; Tohme, J.; Mackenzie, S. Tracing evolutionary and developmental implications of mitochondrial stoichiometric shifting in the common bean. Genetics 2001, 158, 851–864. [Google Scholar] [CrossRef]
- Gualberto, J.M.; Newton, K.J. Plant Mitochondrial Genomes: Dynamics and mechanisms of mutation. Annu. Rev. Plant Biol. 2017, 68, 225–252. [Google Scholar] [CrossRef] [PubMed]
- Bentolila, S.; Stefanov, S. A reevaluation of rice mitochondrial evolution based on the complete sequence of male-fertile and male-sterile mitochondrial genomes. Plant Physiol. 2012, 158, 996–1017. [Google Scholar] [CrossRef]
- Hattori, N.; Kitagawa, K.; Takumi, S.; Nakamura, C. Mitochondrial DNA heteroplasmy in wheat, Aegilops and their nucleus-cytoplasm hybrids. Genetics 2002, 160, 1619–1630. [Google Scholar] [CrossRef]
- Chen, J.; Guan, R.; Chang, S.; Du, T.; Zhang, H.; Xing, H. Substoichiometrically different mitotypes coexist in mitochondrial genomes of Brassica napus L. PLoS ONE 2011, 6, e17662. [Google Scholar] [CrossRef]
- Tanaka, Y.; Tsuda, M.; Yasumoto, K.; Terachi, T.; Yamagishi, H. The complete mitochondrial genome sequence of Brassica oleracea and analysis of coexisting mitotypes. Curr. Genet. 2014, 60, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, Y.; Watase, Y.; Nagase, M.; Makita, N.; Yagura, S.; Hirai, A.; Sugiura, M. The complete nucleotide sequence and multipartite organization of the tobacco mitochondrial genome: Comparative analysis of mitochondrial genomes in higher plants. Mol. Gen. Genome 2005, 272, 603–615. [Google Scholar] [CrossRef]
- Larosa, V.; Remacle, C. Transformation of the mitochondrial genome. Int. J. Dev. Biol. 2013, 57, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Jing, B.; Heng, S.P.; Tong, D.; Wan, Z.J.; Fu, T.D.; Tu, J.X.; Ma, C.Z.; Yi, B.; Wen, J.; Shen, J.X. A male sterility-associated cytotoxic protein ORF288 in Brassica juncea causes aborted pollen development. J. Exp. Bot. 2012, 63, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Antipov, D.; Korobeynikov, A.; McLean, J.S.; Pevzner, P.A. hybridSPAdes: An algorithm for hybrid assembly of short and long reads. Bioinformatics 2016, 32, 1009–1015. [Google Scholar] [CrossRef]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq-versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Lowe, T.M.; Eddy, S.R. tRNAscanSE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | Tibet A | Tibet B |
|---|---|---|
| Genome size (bp) | 239,184 | 258,440 |
| GC content (%) | 45.14 | 45.20 |
| Protein coding genes (types/numbers) | 33/34 | 33/34 |
| tRNAs (types/numbers) | 16/25 | 16/26 |
| tRNAs sequences length (bp) and ratio (%) | 1937/0.81 | 1944/0.75 |
| rRNAs | 3 | 3 |
| rRNAs length (bp) and ratio (%) | 4911/2.05 | 4911/1.91 |
| Protein coding genes sequences length (bp) and ratio (%) | 30,603/12.80 | 30,276/11.71 |
| Intergenic regions sequences length (bp) and ratio (%) | 208,581/87.20 | 228,164/88.29 |
| Chloroplast migration sequences length (bp) and ratio (%) | 7490/3.13 | 7486/2.90 |
| Repeated sequences length (bp) and ratio (%) | 27,217/11.38 | 31,564/12.21 |
| Gene | Position from Start Codon (on CDS) | Nucleotide Variation (Tibet A–Tibet B) | Amino Acid Change (Tibet A–Tibet B) |
|---|---|---|---|
| rps3 | 551 | A-G | E-G |
| matR | 488 | T-C | I-T |
| rpl2 | 204 | A-G | N-D |
| 840 | C-T | R-* Premature stop | |
| atp8 | 370 | A-C | I-L |
| 449 | T-C | V-A | |
| nad7 | 685 | A-G | D-G |
| rps4 | 776 | C-T | S-F |
| ccmC | 351 | G-A | Synonymous |
| atp9b | 64 | A-G | I-V |
| ccmFN1 | 362 | T-* | Frame shifted deletion |
| Features of the orf Genes | orf ID |
|---|---|
| In the unique region | orf463a |
| In the non-unique region | orf115C, orf119C, orf124a, orf107a, orf123a, orf249a, orf261a, orf322a, orf381a, orf474a |
| Chimeric structure | orf463a |
| Transmembrane domain | orf119C, orf123a, orf261a, orf322a, orf381a, orf463a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Liang, L.; Ran, J.; Yang, F.; Ran, M.; Yong, X.; Kong, C.; Tang, Y.; Li, H. Re-Sequencing the Mitochondrial Genome Unveils a Novel Isomeric Form of NWB CMS Line in Radish and Functional Verification of Its Candidate Sterile Gene. Horticulturae 2024, 10, 395. https://doi.org/10.3390/horticulturae10040395
Li X, Liang L, Ran J, Yang F, Ran M, Yong X, Kong C, Tang Y, Li H. Re-Sequencing the Mitochondrial Genome Unveils a Novel Isomeric Form of NWB CMS Line in Radish and Functional Verification of Its Candidate Sterile Gene. Horticulturae. 2024; 10(4):395. https://doi.org/10.3390/horticulturae10040395
Chicago/Turabian StyleLi, Xiaomei, Le Liang, Jianzhao Ran, Feng Yang, Maolin Ran, Xiaoping Yong, Chuibao Kong, Yi Tang, and Huanxiu Li. 2024. "Re-Sequencing the Mitochondrial Genome Unveils a Novel Isomeric Form of NWB CMS Line in Radish and Functional Verification of Its Candidate Sterile Gene" Horticulturae 10, no. 4: 395. https://doi.org/10.3390/horticulturae10040395