
Impact of Wood-Boring Larvae of Xylotrechus arvicola (Coleoptera: Cerambycidae) on Mechanical Properties of Vitis vinifera Plants

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material (Vineyards)
2.2. Grapevine Wood Samples and Experimental Conditions before Mechanical Tests Were Performed
2.3. Mechanical Strength of Grapevine Wood Samples
2.3.1. Experiment 1: Compressive Strength (CS) of Grapevine Wood Trunks in Relation to the Total Damaged Surface Area (TDSA) of Samples
2.3.2. Experiment 2: Flexural Strength (FS) of Grapevine Wood Branches in Relation to the TDSA of Samples
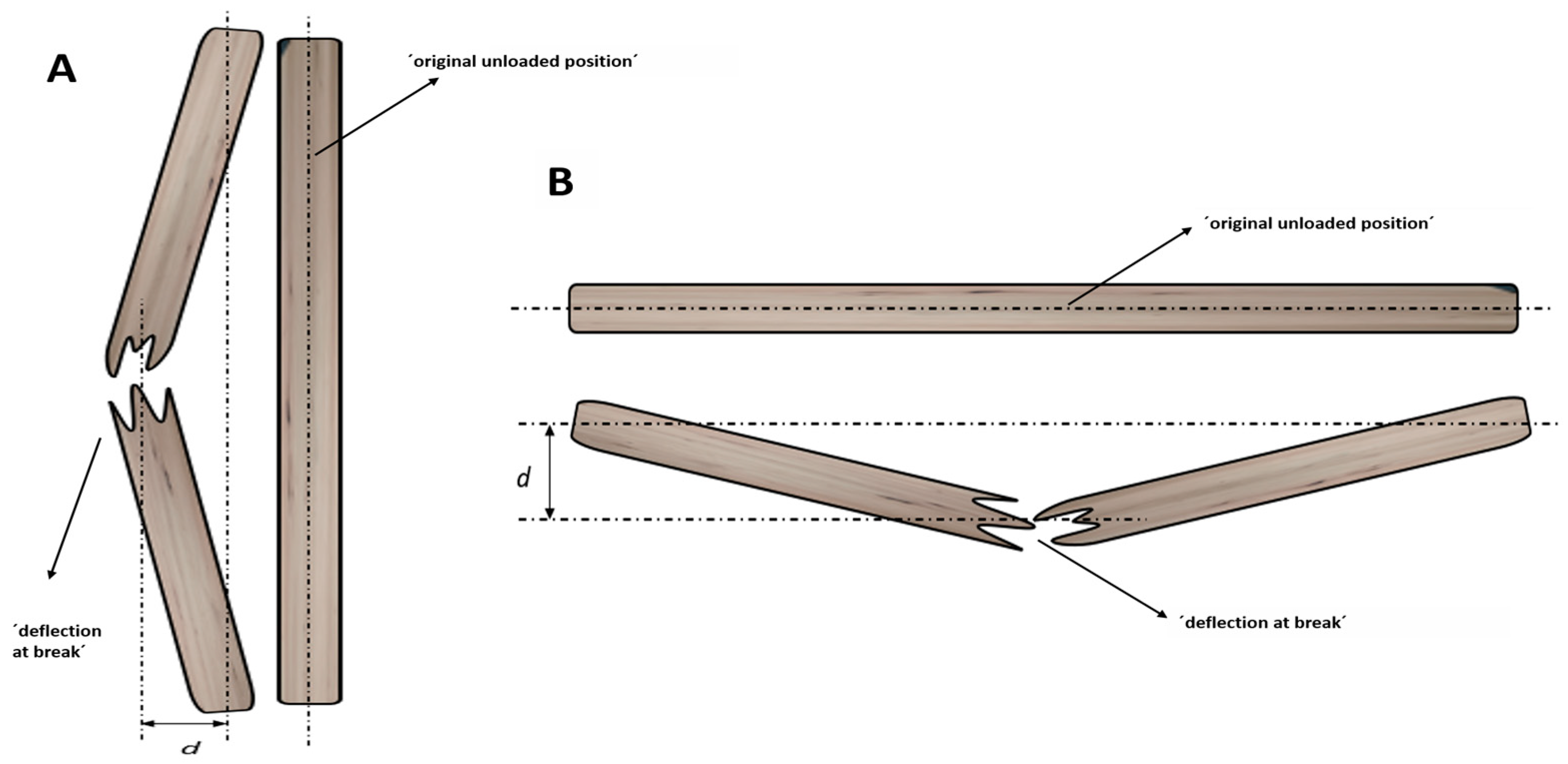
2.4. Experiment 3: Deflection at Break of Grapevine Wood Samples
2.5. Experiment 4: Grapewine Wood Samples with Fungal Growth
2.6. Statistical Analysis
3. Results
3.1. Experiment 1: Mechanical Strength of Grapevine Wood Trunks (CS in Relation to TDSA)
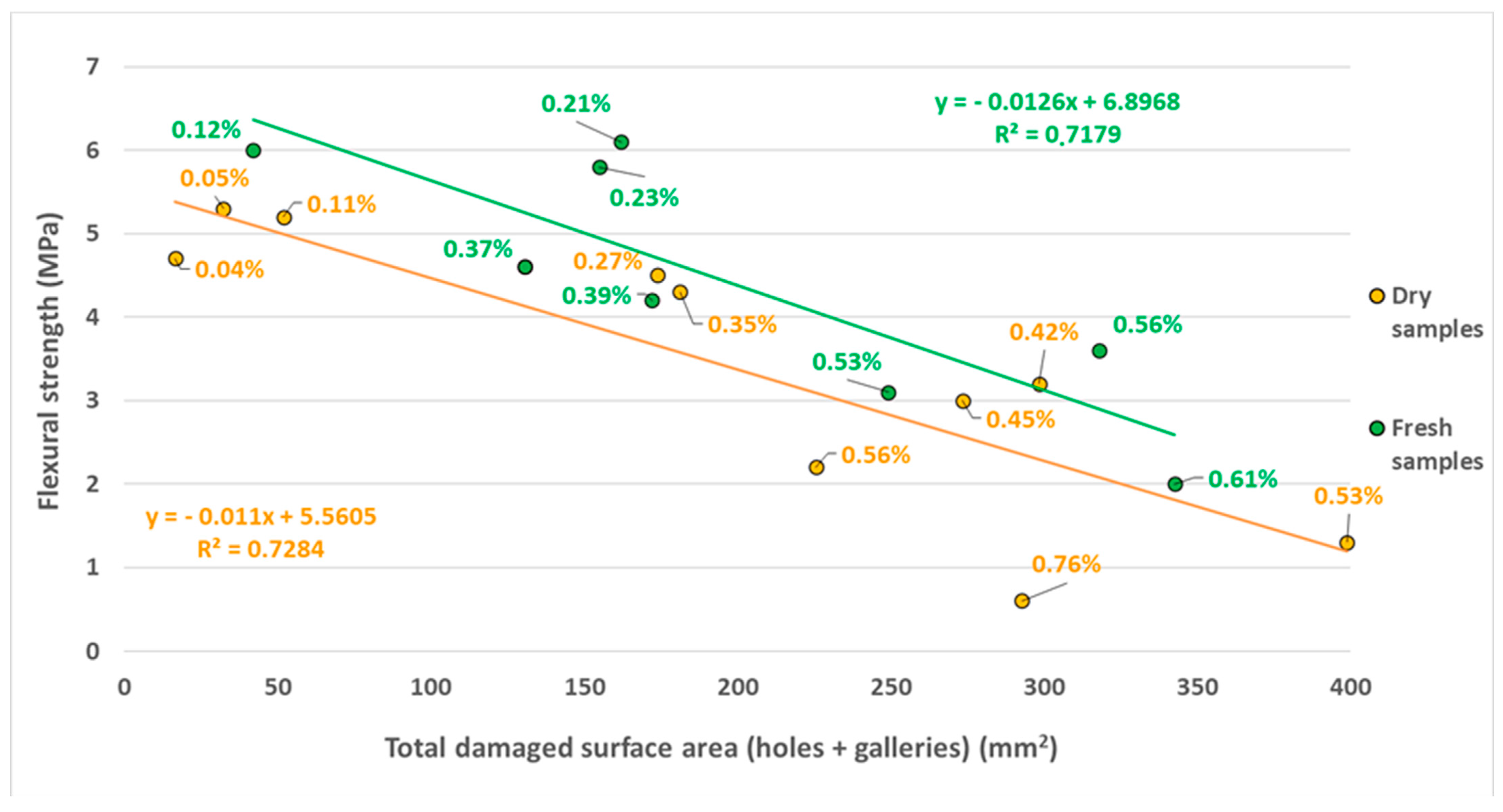
3.2. Experiment 2: Mechanical Strength of Grapevine Wood Branches (FS in Relation to TDSA)
3.3. Experiment 3: Deflection at Break of Grapevine Wood Samples
3.4. Experiment 4: Deflection at Break of Grapevine Wood Samples
4. Discussion
5. Conclusions
- Both CS and FS declined as the TDSA of the wood samples increased, which varied from 0.31% to 0.73% in trunks and from 0.04 to 0.76% in branches, irrespective of the moisture content (fresh or dry). Significant differences between affected grapevine wood samples (fresh and dry) were also confirmed by examining the linear regression coefficients of the interactions CS × TDSA and FS × TDSA.
- Regarding unaffected trunk wood, deflection at break for dry wood samples was lower than that for fresh wood samples (65.00 and 97.85 mm, respectively). With regard to branch wood, deflection at break for affected fresh samples (164.37 mm) was significantly greater than that for both unaffected fresh samples (72.58 mm) and for affected dry samples (37.50 mm).
- Considering the external damages present in wood samples (holes and galleries), a higher percentage of TDSA meant a lower wood resistance.
- Larvae damages on grapevine wood facilitated the spread of grapevine diseases via the emergence of holes created by adult X. arvicola as they exit the wood, as well as through the larval galleries that are connected to them. The percentage of symptoms of fungal growth for affected wood samples were 66.66% for dry wood trunks, 75.00% for fresh wood branches, and 60.00% for dry wood branches.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Syofuna, A.; Banana, A.; Nakabonge, G. Efficiency of natural wood extractives as wood preservatives against termite attack. Maderas Cienc. Tecnol. 2012, 14, 155–163. [Google Scholar] [CrossRef]
- Sen, S.; Yalçin, M.; Tasçioglu, C.; Özbayram, A.K. Larvicidal activities of some bark and wood extracts against wood-damaging insects. Maderas Cienc. Tecnol. 2017, 19, 273–284. [Google Scholar] [CrossRef]
- Visitpanich, J. The biology and survival rate of the coffee stem borer, Xylotrechus quadripes Chevrolat (Coleoptera: Cerambycidae). Jpn. J. Entomol. 1994, 62, 731–745. [Google Scholar]
- Aukema, J.E.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; McCullough, D.G.; et al. Economic impacts of non native forest insects in the continental. United States. PLoS ONE 2011, 6, e24587. [Google Scholar] [CrossRef] [PubMed]
- Svacha, P.; Danilevsky, M.L. Cerambycoid larvae of Europe and Soviet Union (Coleoptera, Cerambycoidea). Part II. Acta Univ. Carol. 1988, 31, 129–279. Available online: https://www.zin.ru/animalia/coleoptera/eng/svadanla.htm (accessed on 10 April 2024).
- Sama, G. Northern, Western, central and Eastern Europe, British Isles and Continental Europe from France (excluding Corsica) to Scandinavia and Urals. In Atlas of the Cerambycidae of Europe and the Mediterranean Area; Nakladatelstvi Kabourek: Zlin, Czech Republic, 2002; Volume 1, 173p. [Google Scholar]
- Bahillo, P. Cerambícidos (Coleoptera, Cerambycidae) del País Vasco. Cuad. Investig. Biológica 1996, 19, 274. [Google Scholar]
- Vives, E. Coleoptera: Cerambycidae. In Fauna Ibérica; Ramos, M.A., Ed.; Museo Nacional de Ciencias Naturales; Consejo Superior de Investigaciones Científicas (CSIC): Madrid, Spain, 2000; Volume 12. [Google Scholar]
- Moreno, C.M. Xylotrechus arvicola (Olivier 1795) (Coleóptera: Cerambycidae): Descripción Morfológica, Ciclo Biológico, Incidencia y Daños en el Cultivo de la Vid. Ph.D. Thesis, Publicaciones del Instituto Tecnológico Agrario de Castilla y León (ITACYL), Valladolid, Spain, 2005. [Google Scholar]
- Biurrun, R.; Yanguas, R.; Garnica, I.; Benito, A. Xylotrechus arvicola. El taladro del endrino. Navar. Agrar. 2007, 164, 47–51. [Google Scholar]
- Ocete, R.; Del Tío, R. Presencia del perforador Xylotrechus arvicola (Olivier) (Coleoptera: Cerambycidae) en viñedos de la Rioja Alta. Bol. Sanid. Veg. Plagas 1996, 22, 199–202. [Google Scholar]
- Rodríguez, M.; Ocaña, P. Presencia del perforador Xylotrechus arvicola (Olivier) en viñas de la provincia de Ciudad Real-1996. In Proceedings of the XXII Reunión Del Grupo de Trabajo de Los Problemas Fitosanitarios de La Vid, Ciudad Real, Spain, 18–20 February 1997. [Google Scholar]
- Ocete, R.; López, M.A. Principales insectos xilófagos de los viñedos de la Rioja Alta y Alavesa. Vitic. Enol. Prof. 1999, 62, 24–30. [Google Scholar]
- Peláez, H.; Maraña, J.R.; Urbez, J.R.; Barrigón, J.M. Xylotrechus arvicola (Olivier, 1795) (Coleoptera: Cerambycidae). Presencia en los viñedos de Castilla y León. IV Congreso Ibérico de Ciencias Hortícolas; (Extremadura, Spain: Caceres). Actas Hortic. 2001, 30, 1326–1332. [Google Scholar]
- Ocete, R.; López, M.; Prendes, C.; Lorenzo, C.; González-Andújar, J.; Lara, M. Xylotrechus arvicola (Olivier) (Coleoptera, Cerambycidae), a new impacting pest on Spanish vineyards. Vitis 2002, 41, 211–212. [Google Scholar] [CrossRef]
- Moreno, C.M.; Martín, C.M.; Urbez, J.R.; Maraña, R.; Moro, S.; García, D.; Peláez, H. Descripción de dos coleópteros que afectan al viñedo en Castilla y León. Phytoma 2003, 147, 34–42. [Google Scholar]
- Peláez, H.; Hernández, J.M.; Martín, M.C.; Moreno, C.M.; Santiago, Y. Determinación De Las Características Del Huevo de Xylotrechus arvicola (Coleoptera: Cerambycidae, Olivier, 1795). In Libro de Actas del X Congreso Ibérico de Entomología; de Diputación, Z., Ed.; Castilla y León: Zamora, Spain, 2002; p. 52. [Google Scholar]
- Rodríguez-González, A.; Peláez, H.J.; González-López, O.; Mayo, S.; Casquero, P.A. Reproductive patterns of Xylotrechus arvicola (Coleoptera: Cerambycidae), an emerging pest of grapevines, under laboratory conditions. J. Econ. Entomol. 2016, 109, 1226–1230. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-González, A.; Mayo, S.; González-López, O.; Peláez, H.J.; Casquero, P.A. Biological parameters of Xylotrechus arvicola females, an insect pest in Iberian Peninsula vineyards. Oeno One 2017, 51, 373–379. [Google Scholar] [CrossRef]
- Rodríguez-González, A.; Peláez, H.J.; Mayo, S.; González-López, O.; Casquero, P.A. Egg development and toxicity of insecticides to eggs, neonate larvae and adults of Xylotrechus arvicola, a pest in Iberian grapevines. Vitis 2016, 55, 83–93. [Google Scholar] [CrossRef]
- García-Ruiz, E. Contribución al Manejo de Plagas en Vid: Xylotrechus arvicola Olivier (Coleoptera: Cerambycidae) y Lobesia botrana Denis & Schiffermüller (Lepidoptera: Tortricidae). Ph.D. Thesis, University of La Rioja, Logroño, Spain, 2009. [Google Scholar]
- Rodríguez-González, A.; Mayo, S.; González-López, O.; Reinoso, B.; Gutiérrez, S.; Casquero, P.A. Inhibitory activity of Beauveria bassiana and Trichoderma spp. on the insect pests Xylotrechus arvicola (Coleoptera: Cerambycidae) and Acanthoscelides obtectus (Coleoptera: Chrisomelidae: Bruchinae). Environ. Monit. Assess. 2017, 189, 12. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-González, A.; Malvar, R.A.; Guerra, M.; Sánchez-Maíllo, E.; Peláez, H.J.; Carro-Huerga, G.; Casquero, P.A. Xylotrechus arvicola (Coleoptera: Cerambycidae) capture in vineyards in relation to climatic factors. Pest Manag. Sci. 2022, 78, 3030–3038. [Google Scholar] [CrossRef]
- García-Benavides, P.; Martin-Zamorano, P.; Ocete-Pérez, C.A.; Maistrello, L.; Ocete, R. Biodiversity of pathogenic wood fungi isolated from Xylotrechus arvicola (Olivier) galleries in vine shoots. J. Int. Sci. Vigne Vin 2013, 47, 73–81. [Google Scholar] [CrossRef]
- Ocete, R.; López-Martínez, M.A.; Prendes, C.; Lorenzo, C.D.; González-Andújar, J.L. Relación entre la infestación de Xylotrechus arvicola (Coleoptera: Cerambycidae) (Olivier) y la presencia de hongos patógenos en un viñedo de la Denominación de Origen “La Mancha”. Bol. San. Veg. Plagas 2002, 28, 97–102. [Google Scholar]
- Peláez, H.; Moreno, C.; Santiago, Y.; Maraña, R.; Urbez, J.R.; Lambert, S.M.; María, C.M.; Evan, E.; Barrigón, J.; Prada, P.V. Xylotrechus arvicola: Un cerambícido en el cultivo de la vid. Terralia 2006, 55, 50–56. [Google Scholar]
- Ocete, R.; López, M.; Gallardo, A.; Pérez, M.; Rubio, I. Efecto de la infestación de Xylotrechus arvicola (Olivier) (Coleoptera: Cerambycidae) sobre la floración de la variedad Tempranillo en La Rioja. Bol. San. Veg. Plagas 2004, 30, 311–316. Available online: https://www.miteco.gob.es/ministerio/pags/biblioteca/plagas/BSVP-30-02-311-316.pdf (accessed on 10 April 2024).
- Rodríguez-González, A.; Peláez, H.J.; Mayo, S.; González-López, O.; Casquero, P.A. Biometric traits of Xylotrechus arvicola adults from laboratory and grape field. Vitis 2016, 55, 73–78. [Google Scholar] [CrossRef]
- Rodríguez-González, A.; Casquero, P.A.; García-González, J.; Rodríguez-Robles, D.; Morán-Del Pozo, J.M.; Juan-Valdés, A. Analysis of the mechanical properties of wood attacked by Xylotrechus arvicola larvae, and its influence on the structural properties of the plant. Vitis 2019, 58, 105–112. [Google Scholar] [CrossRef]
- Rodríguez-González, A.; Casquero, P.A.; Carro-Huerga, G.; García-González, J.; Álvarez-García, S.; Juan-Valdés, A. Failure under stress of grapevine wood: The effects of the cerambycid Xylotrechus arvicola on the biomechanics properties of Vitis vinifera. Maderas—Cienc. Tecnol. 2020, 22, 167–178. [Google Scholar] [CrossRef]
- AENOR—Spanish Association for Standardization and Certification. Website of Spanish Association for Standardization and Certification. 2024. Available online: http://www.aenor.es/ (accessed on 15 January 2024).
- Gere, J.M.; Timoshenko, S.P. Mechanics of Materials, 2nd ed.; PWS Publishers Co.: Worcester, UK, 1984; ISBN 0-534-03099-8. [Google Scholar]
- Gramaje, D.; Urbez-Torres, J.R.; Sosnowski, M.R. Managing grapevine trunk diseases with respect to etiology and epidemiology: Current strategies and future prospects. Plant Dis. 2018, 102, 12–39. [Google Scholar] [CrossRef] [PubMed]
- Di Iorio, O.R. Torneutini of Argentina: New records, host plants, and comparison of their larval and adult biologies with those of Prioninae and Trachyderni (Coleoptera: Cerambycidae). G. Ital. Entomol. 2006, 11, 183–234. [Google Scholar]
- Walczynska, A.; Danko, M.; Kozlowski, J. The considerable adult size variability in wood feeders is optimal. Ecol. Entomol. 2010, 35, 16–24. [Google Scholar] [CrossRef]
- Walczynska, A. Is wood safe for its inhabitants? Bull. Entomol. Res. 2010, 100, 461–465. [Google Scholar] [CrossRef]
- Hanks, L.M. Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annu. Rev. Entomol. 1999, 44, 483–505. [Google Scholar] [CrossRef]
- Ferrero, M.E.; Coirini, R.O.; Díaz, M.P. The effect of wood-boring beetles on the radial growth of Prosopis flexuosa DC. In the arid Chaco of Argentina. J. Arid Environ. 2013, 88, 141–146. [Google Scholar] [CrossRef]
- Moreno, C.M.; Martín, Y.; Santiago, Y.; De Evan, E.; Hernández, J.M.; Peláez, H. Presencia de Xylotrechus arvicola (Olivier, 1795) (Coleoptera: Cerambycidae) en viñedos de la zona centro de Castilla y León. Bol. San. Veg. Plagas. 2004, 30, 475–486. [Google Scholar]
- Soltis, N.E.; Gomez, S.; Leisk, G.G.; Sherwood, P.; Preisser, E.L.; Bonello, P.; Orians, C.M. Failure under stress: The effect of the exotic herbivore Adelges tsugae on biomechanics of Tsuga canadensis. Ann. Bot. 2014, 113, 721–730. [Google Scholar] [CrossRef] [PubMed]
- McClure, M.S. Density-dependent feedback and population cycles in Adelges tsugae (Homoptera: Adelgidae) on Tsuga canadensis. Environ. Entomol. 1991, 20, 258–264. [Google Scholar] [CrossRef]
- Stadler, B.; Müller, T.; Orwig, D.; Cobb, R. Hemlock woolly adelgid in New England forests: Canopy impacts transforming ecosystem processes and landscapes. Ecosystems 2005, 8, 233–247. [Google Scholar] [CrossRef]
- Jankowiak, R.; Rossa, R. Filamentous fungi associated with Monochamus galloprovincialis and Acanthocinus aedilis (Coleoptera: Cerambycidae) in scots pine. Pol. Bot. J. 2007, 52, 143–149. [Google Scholar]
- Spatz, H.C.; Bruechert, F. Basic biomechanics of self-supporting plants: Wind loads and gravitational loads on a Norway spruce tree. For. Ecol. Manag. 2000, 135, 33–44. [Google Scholar] [CrossRef]
- Solomon, J.D. Guide to insect borers of North American broadleaf trees and shrubs. In Forest Service Agriculture Handbook AH-706; United States Department of Agricluture: Washington, DC, USA, 1995. [Google Scholar]
- Stephen, F.M.; Salisbury, V.B.; Oliveria, F.L. Red oak borer, Enaphalodes rufulus (Coleoptera: Cerambycidae), in the Ozark Mountains of Arkansas, USA: An unexpected and remarkable forest disturbance. Integr. Pest Manag. Rev. 2003, 6, 247–252. [Google Scholar] [CrossRef]
- Buchner, P. Endosymbiosis of Animals with Plant Microorganisms; John Wiley & Sons: New York, NY, USA, 1965. [Google Scholar]
- Dominik, J.; Starzyk, J.R. Owady Niszczące Drewno; Państwowe Wydawnictwo Rolnicze i Leśne: Warsaw, Poland, 1989. [Google Scholar]
- Jones, K.G.; Dowd, P.F.; Blackwell, M. Polyphyletic origins of yeast-like endocytobionts from anobiid and cerambycid beetles. Mycol. Res. 1999, 103, 542–546. [Google Scholar] [CrossRef]
- Kurkela, T.; Aalto, T.; Varama, M.; Jalkanen, R. Defoliation by the common pine sawfly (Diprion pini) and subsequent growth reduction in Scots pine: A retrospective approach. Silva Fenn. 2005, 39, 467–480. [Google Scholar] [CrossRef]
- Drenkhan, R.; Kurkela, T.; Hanso, M. The relationship between the needle age and the growth rate in Scots pine (Pinus sylvestris): A retrospective analysis by Needle Trace Method (NTM). Eur. J. For. Res. 2006, 125, 397–405. [Google Scholar] [CrossRef]
- Hansen, E.M.; Stone, J.K.; Capitano, B.R.; Rosso, P.; Sutton, W.; Winton, L. Incidence and impact of Swiss needle cast in forest plantations of Douglas fir in coastal Oregon. Plant Dis. 2000, 84, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Krause, S.C.; Raffa, K.F. Comparison of insect, fungal, and mechanically induced defoliation of larch: Effects on plant productivity and subsequent host susceptibility. Oecologia 1992, 90, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Hauer, R.; Wing, J.W.; Dawson, J.O. Ice storm damage to urban trees. J. Arboric. 1993, 19, 187–193. [Google Scholar] [CrossRef]
- James, K.; Kane, B. Precision digital instruments to measure dynamic wind loads on trees during storms. Agric. For. Meteorol. 2008, 148, 1055–1061. [Google Scholar] [CrossRef]
- Detters, A.; Cowell, C.; McKeown, L.; Howard, P. Evaluation of Current Rigging and Dismantling Practices Used in Arboriculture; Report submitted to the Health and Safety Executive and the Forestry Commission; Health and Safety Executive (HSE): Bootle, UK, 2008; 355p.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grapevine Trunks | ESA of Sample (mm2) | ESA of Damages (mm2) | TDSA (Holes + Galleries) (mm2) | TDSA (%) | Class | ||
|---|---|---|---|---|---|---|---|
| Holes (n) * | Galleries (v) ** | ||||||
| Fresh samples | 1 | 99,665.75 | - | - | - | - | - |
| 2 | 171,378.63 | - | - | - | - | - | |
| 3 | 100,192.53 | - | - | - | - | - | |
| 4 | 104,709.14 | - | - | - | - | - | |
| 5 | 101,257.14 | - | - | - | - | - | |
| 6 | 98,208.14 | - | - | - | - | - | |
| 7 | 99,592.39 | - | - | - | - | - | |
| 1 | 114,633.97 | 17.60 (1) | 570.25 (2) | 587.85 | 0.51 | 3 | |
| 2 | 94,995.80 | 19.79 (1) | 272.80 (2) | 292.59 | 0.31 | 1 | |
| 3 | 135,591.07 | - (0) | 558.84 (14) | 558.84 | 0.41 | 2 | |
| 4 | 169,054.48 | 144.72 (14) | 793.45 (9) | 938.17 | 0.55 | 3 | |
| Dry samples | 1 | 92,907.60 | - | - | - | - | - |
| 2 | 70,285.55 | - | - | - | - | - | |
| 3 | 40,898.55 | - | - | - | - | - | |
| 4 | 38,466.37 | - | - | - | - | - | |
| 5 | 32,664.39 | - | - | - | - | - | |
| 6 | 50,357.97 | - | - | - | - | - | |
| 7 | 88,423.09 | - | - | - | - | - | |
| 8 | 64,874.71 | - | - | - | - | - | |
| 9 | 67,044.42 | - | - | - | - | - | |
| 10 | 145,707.45 | - | - | - | - | - | |
| 11 | 159,414.31 | - | - | - | - | - | |
| 12 | 145,493.83 | - | - | - | - | - | |
| 1 | 98,835.75 | 29.19 (2) | 439.49 (4) | 468.68 | 0.47 | 2 | |
| 2 | 37,335.64 | 34.73 (2) | 167.97 (5) | 204.70 | 0.54 | 3 | |
| 3 | 68,825.39 | 61.12 (4) | 168.70 (4) | 229.82 | 0.33 | 1 | |
| 4 | 77,075.59 | 25.53 (2) | 428.37 (3) | 453.90 | 0.58 | 3 | |
| 5 | 120,266.59 | 59.02 (3) | 820.20 (6) | 879.22 | 0.73 | 3 | |
| 6 | 28,099.99 | - (0) | 117.53 (3) | 117.53 | 0.41 | 2 | |
| 7 | 97,994.55 | 57.68 (3) | 594.51 (4) | 652.19 | 0.66 | 3 | |
| 8 | 61,750.37 | 78.31 (3) | 315.31 (2) | 393.62 | 0.63 | 3 | |
| 9 | 206,440.81 | 71.88 (4) | 1131.92 (2) | 1203.80 | 0.58 | 3 | |
| 10 | 160,910.17 | 34.11 (2) | 814.95 (1) | 849.06 | 0.53 | 3 | |
| 11 | 136,689.25 | - (0) | 716.92 (4) | 716.92 | 0.52 | 3 | |
| 12 | 122,840.49 | - (0) | 713.05 (2) | 713.05 | 0.58 | 3 | |
| Grapevine Branches | ESA of Sample (mm2) | ESA of Damages (mm2) | TDSA (Holes + Galleries) (mm2) | TDSA (%) | Class | ||
|---|---|---|---|---|---|---|---|
| Holes (n) * | Galleries (v) ** | ||||||
| Fresh samples | 1 | 66,380.61 | - | - | - | - | - |
| 2 | 59,697.05 | - | - | - | - | - | |
| 3 | 77,196.38 | - | - | - | - | - | |
| 4 | 46,292.06 | - | - | - | - | - | |
| 5 | 41,982.93 | - | - | - | - | - | |
| 6 | 54,320.83 | - | - | - | - | - | |
| 7 | 53,935.75 | - | - | - | - | - | |
| 8 | 73,918.05 | - | - | - | - | - | |
| 9 | 37,208.91 | - | - | - | - | - | |
| 10 | 45,797.81 | - | - | - | - | - | |
| 11 | 66,177.12 | - | - | - | - | - | |
| 12 | 45,024.32 | - | - | - | - | - | |
| 1 | 34,765.54 | 36.17 (2) | 94.48 (2) | 130.65 | 0.37 | 1 | |
| 2 | 43,831.52 | 53.67 (3) | 118.41 (3) | 172.08 | 0.39 | 1 | |
| 3 | 64,609.07 | 56.34 (3) | 98.61 (1) | 154.95 | 0.23 | 1 | |
| 4 | 76,528.70 | 53.42 (3) | 108.60 (1) | 162.02 | 0.21 | 1 | |
| 5 | 56,372.96 | 31.82 (2) | 310.79 (4) | 342.61 | 0.61 | 3 | |
| 6 | 33,846.14 | 42.01 (3) | - (0) | 42.01 | 0.12 | 1 | |
| 7 | 46,355.11 | 70.20 (5) | 178.93 (3) | 249.13 | 0.53 | 3 | |
| 8 | 56,433.62 | 43.65 (4) | 274.46 (3) | 318.11 | 0.56 | 3 | |
| Dry samples | 1 | 46,231.24 | - | - | - | - | - |
| 2 | 64,228.78 | - | - | - | - | - | |
| 3 | 67,981.04 | - | - | - | - | - | |
| 4 | 67,059.61 | - | - | - | - | - | |
| 5 | 68,540.18 | - | - | - | - | - | |
| 6 | 54,579.34 | - | - | - | - | - | |
| 7 | 53,807.26 | - | - | - | - | - | |
| 8 | 46,149.63 | - | - | - | - | - | |
| 9 | 83,990.55 | - | - | - | - | - | |
| 10 | 72,089.69 | - | - | - | - | - | |
| 11 | 72,451.35 | - | - | - | - | - | |
| 12 | 84,694.19 | - | - | - | - | - | |
| 1 | 59,525.32 | 19.01 (1) | 254.40 (2) | 273.41 | 0.45 | 2 | |
| 2 | 38,104.89 | 33.70 (2) | 259.06 (5) | 292.76 | 0.76 | 3 | |
| 3 | 57,272.18 | - (0) | 325.56 (2) | 225.56 | 0.56 | 3 | |
| 4 | 51,040.10 | - (0) | 181.19 (2) | 181.19 | 0.35 | 1 | |
| 5 | 62,042.15 | 32.20 (2) | - (0) | 32.20 | 0.05 | 1 | |
| 6 | 63,107.76 | 18.70 (1) | 155.02 (1) | 173.72 | 0.27 | 1 | |
| 7 | 74,550.08 | 19.24 (1) | 379.51 (2) | 398.75 | 0.53 | 3 | |
| 8 | 70,947.80 | 18.62 (1) | 279.71 (4) | 298.33 | 0.42 | 2 | |
| 9 | 44,752.36 | 16.54 (1) | - (0) | 16.54 | 0.04 | 1 | |
| 10 | 45,376.31 | - (0) | 51.59 (1) | 51.95 | 0.11 | 1 | |
| Compressive Strength | Flexural Strength | ||||||
|---|---|---|---|---|---|---|---|
| Unaffected Wood (n) (s) | Affected Wood (n) (s) | Unaffected Wood (n) (s) | Affected Wood (n) (s) | ||||
| Fresh samples | 97.85 ± 7.47 aA (7 samples) (7.3 MPa) | 83.75 ± 21.92 aA (4 samples) (3.0 Mpa) | F = 0.562 df = 1.9 p = 0.473 | Fresh samples | 72.58 ± 11.31 bA (12 samples) (72.33 MPa) | 164.37 ± 48.71 aA (8 samples) (76.52 MPa) | F = 4.859 df = 1.18 p = 0.041 |
| Dry samples | 65.00 ± 8.18 aB (12 samples) (9.8 MPa) | 86.67 ± 7.16 aA (12 samples) (5.4 MPa) | F = 3.969 df = 1.22 p = 0.059 | Dry samples | 77.92 ± 5.69 aA (12 samples) (43.65 MPa) | 37.50 ± 8.37 bB (10 samples) (30.45 MPa) | F = 16.835 df = 1.20 p ≤ 0.001 |
| F= | 7.249 | 0.029 | F= | 0.177 | 8.221 | ||
| df= | 1.17 | 1.14 | df= | 1.22 | 1.16 | ||
| p= | 0.015 | 0.868 | p= | 0.678 | 0.011 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Lozano, D.; Antolín-Rodríguez, A.; Carro-Huerga, G.; Zanfaño, L.; Casquero, P.A.; Guerra, M.; Juan-Valdés, A.; Rodríguez-González, Á. Impact of Wood-Boring Larvae of Xylotrechus arvicola (Coleoptera: Cerambycidae) on Mechanical Properties of Vitis vinifera Plants. Horticulturae 2024, 10, 431. https://doi.org/10.3390/horticulturae10050431
Ramírez-Lozano D, Antolín-Rodríguez A, Carro-Huerga G, Zanfaño L, Casquero PA, Guerra M, Juan-Valdés A, Rodríguez-González Á. Impact of Wood-Boring Larvae of Xylotrechus arvicola (Coleoptera: Cerambycidae) on Mechanical Properties of Vitis vinifera Plants. Horticulturae. 2024; 10(5):431. https://doi.org/10.3390/horticulturae10050431
Chicago/Turabian StyleRamírez-Lozano, Daniela, Andrea Antolín-Rodríguez, Guzmán Carro-Huerga, Laura Zanfaño, Pedro A. Casquero, Marcos Guerra, Andrés Juan-Valdés, and Álvaro Rodríguez-González. 2024. "Impact of Wood-Boring Larvae of Xylotrechus arvicola (Coleoptera: Cerambycidae) on Mechanical Properties of Vitis vinifera Plants" Horticulturae 10, no. 5: 431. https://doi.org/10.3390/horticulturae10050431