

Combined Pretreatment with Bioequivalent Doses of Plant Growth Regulators Alleviates Dehydration Stress in Lactuca sativa
 , ,
, ,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Growth Conditions and Plant Material
2.1.1. Hydroponic Experiment with 12% PEG
2.1.2. Soil Drought Experiment (“Proof-of-Concept”)
2.2. Biochemical Analyses
2.2.1. Stress Markers and Non-Enzyme Antioxidants
2.2.2. Antioxidant Enzyme Activities
2.3. Immunoblot Analyses of Dehdyrins
2.4. RT-qPCR Analysis of SOD, CAT and Peroxidase-Coding Genes
2.5. Statistical Analyses
3. Results and Discussion
3.1. The Combined Pretreatment with Bioequivalent Amounts of ABA, ACC, NAA and KIN Ameliorates the Negative Effect of Dehydration Stress Provoked by 12% PEG in Hydroponically Grown Lactuca sativa
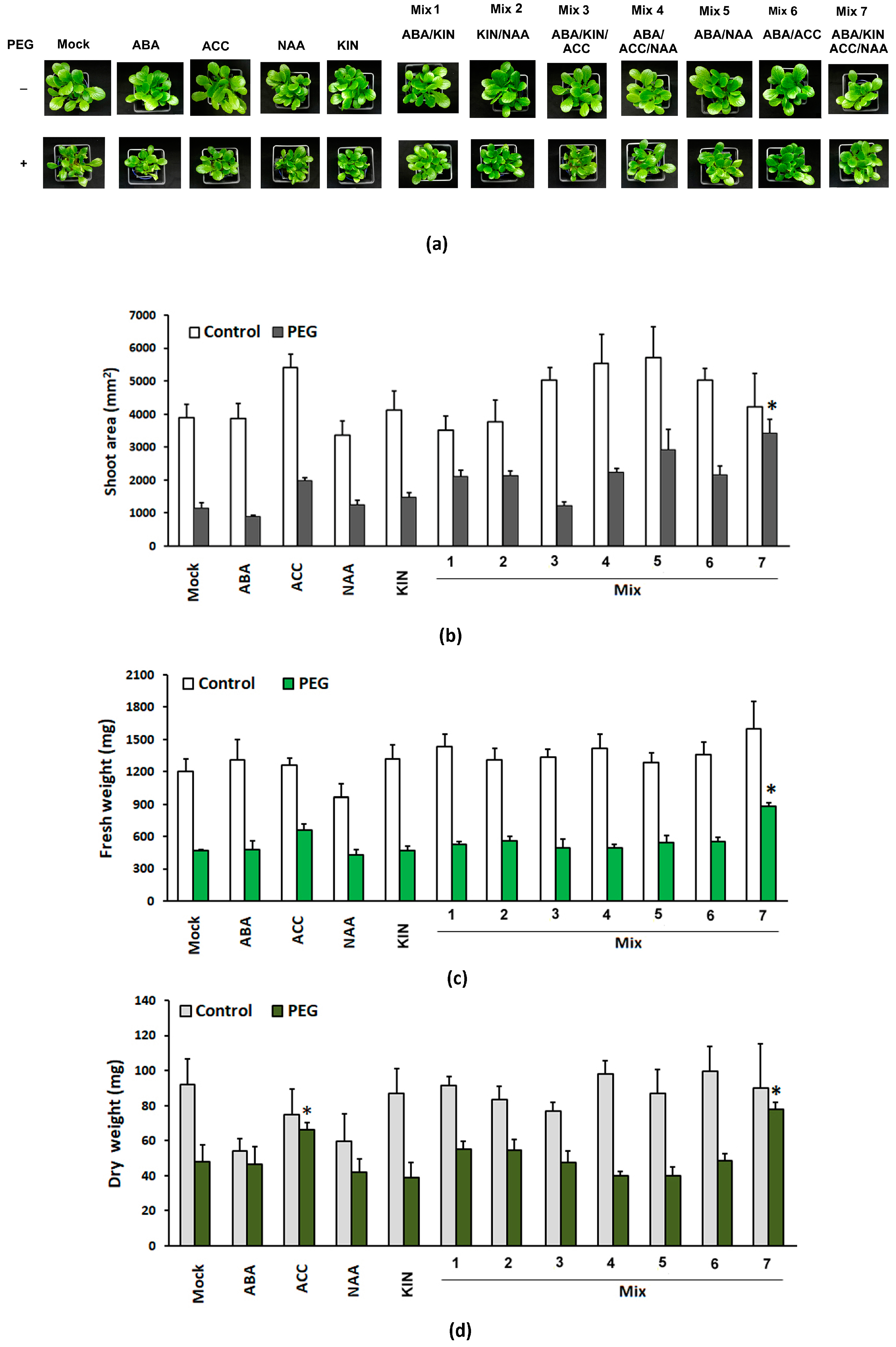
3.1.1. Growth Parameters
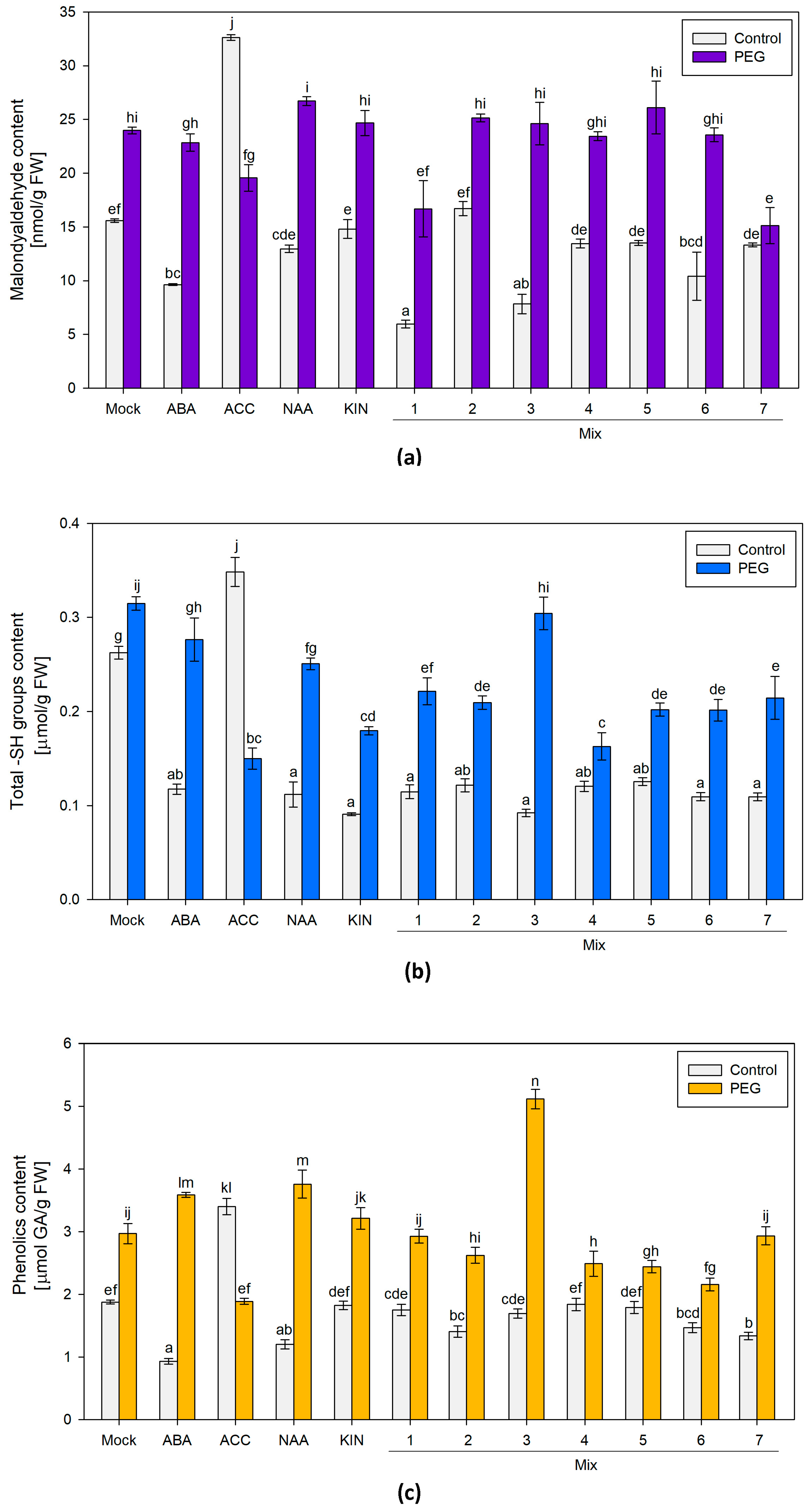
3.1.2. Malondialdehyde (MDA), Free Sulfhydryl Groups (SH-Groups) and Total Phenolic Compounds
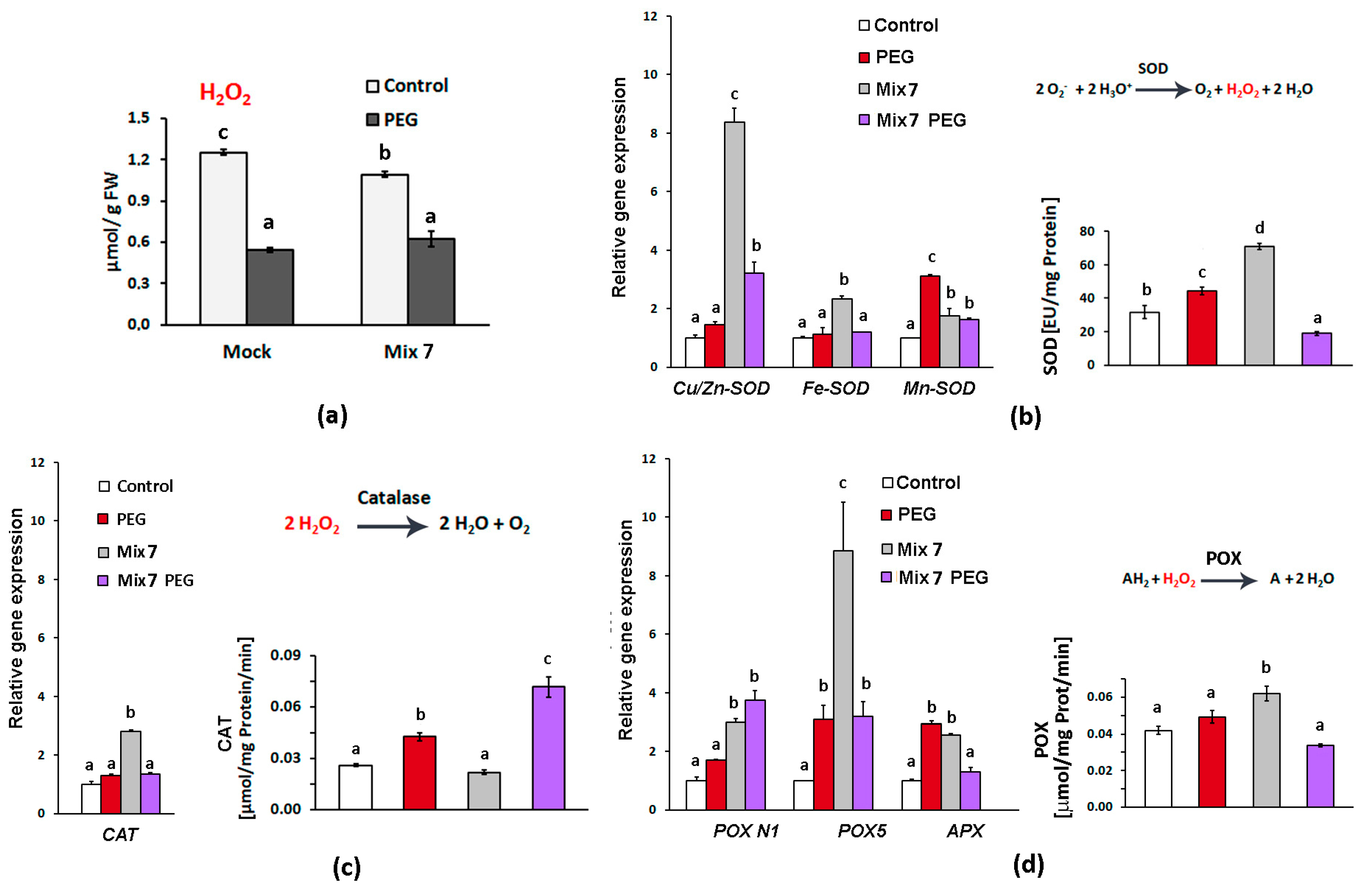
3.2. Combined Pretreatment with ABA, ACC, NAA and KIN Activates Enzymatic Antioxidant Defense
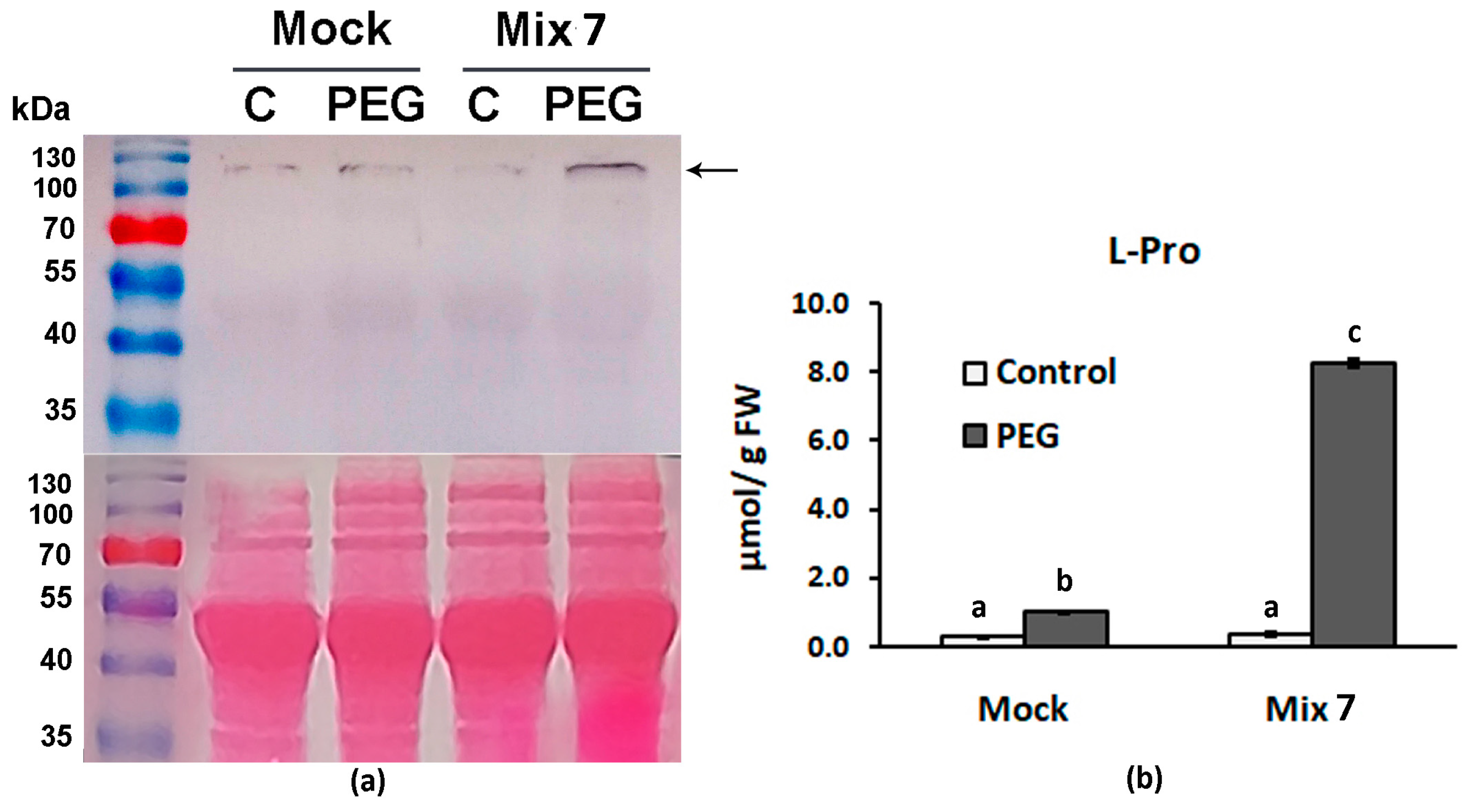
3.3. Combined Pretreatment with ABA, ACC, NAA and KIN Positively Affects the Accumulation of Free Proline upon Dehydration
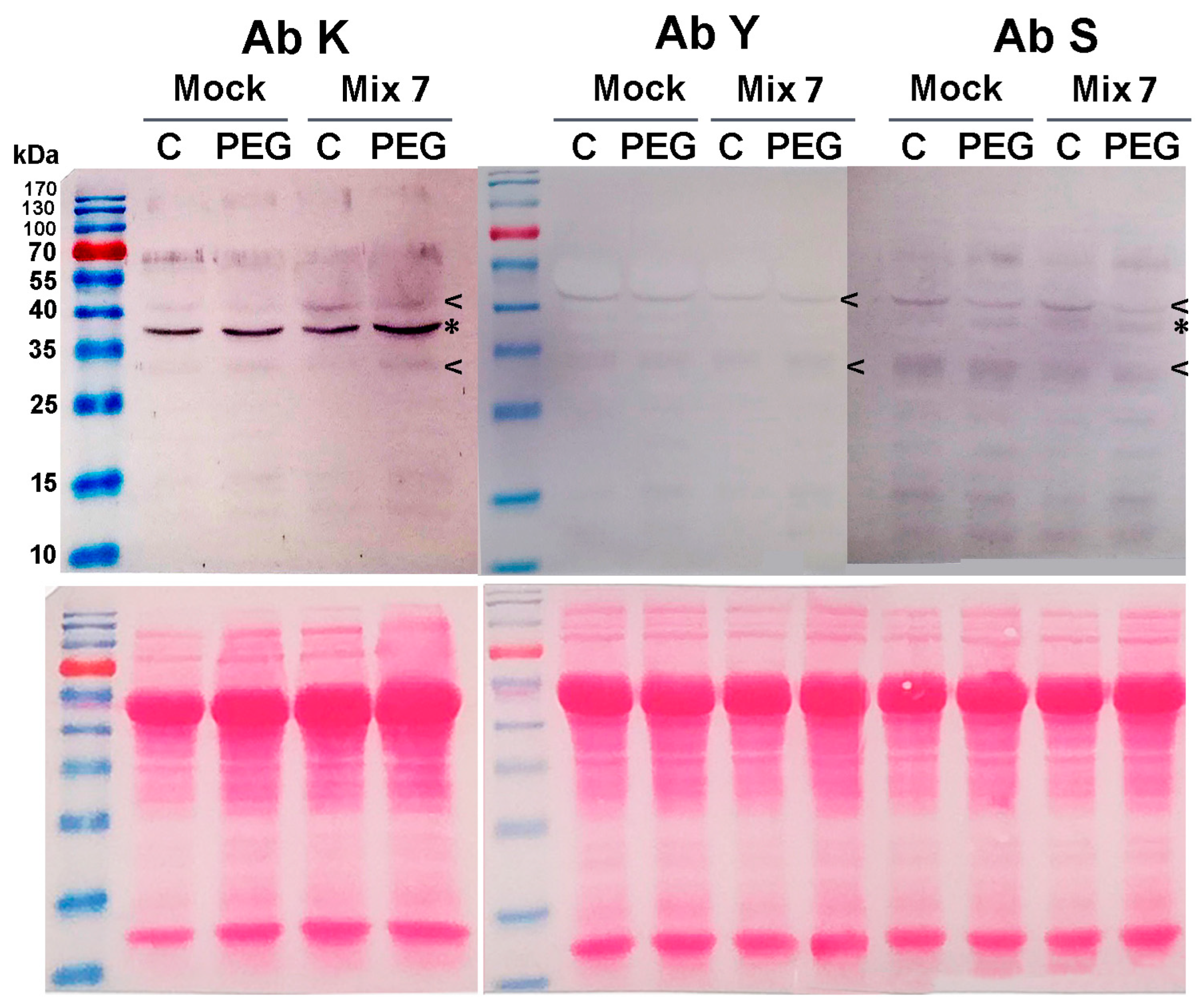
3.4. Combined Pretreatment with ABA, ACC, NAA and KIN Stimulates the Accumulation of Certain Dehydrin Types
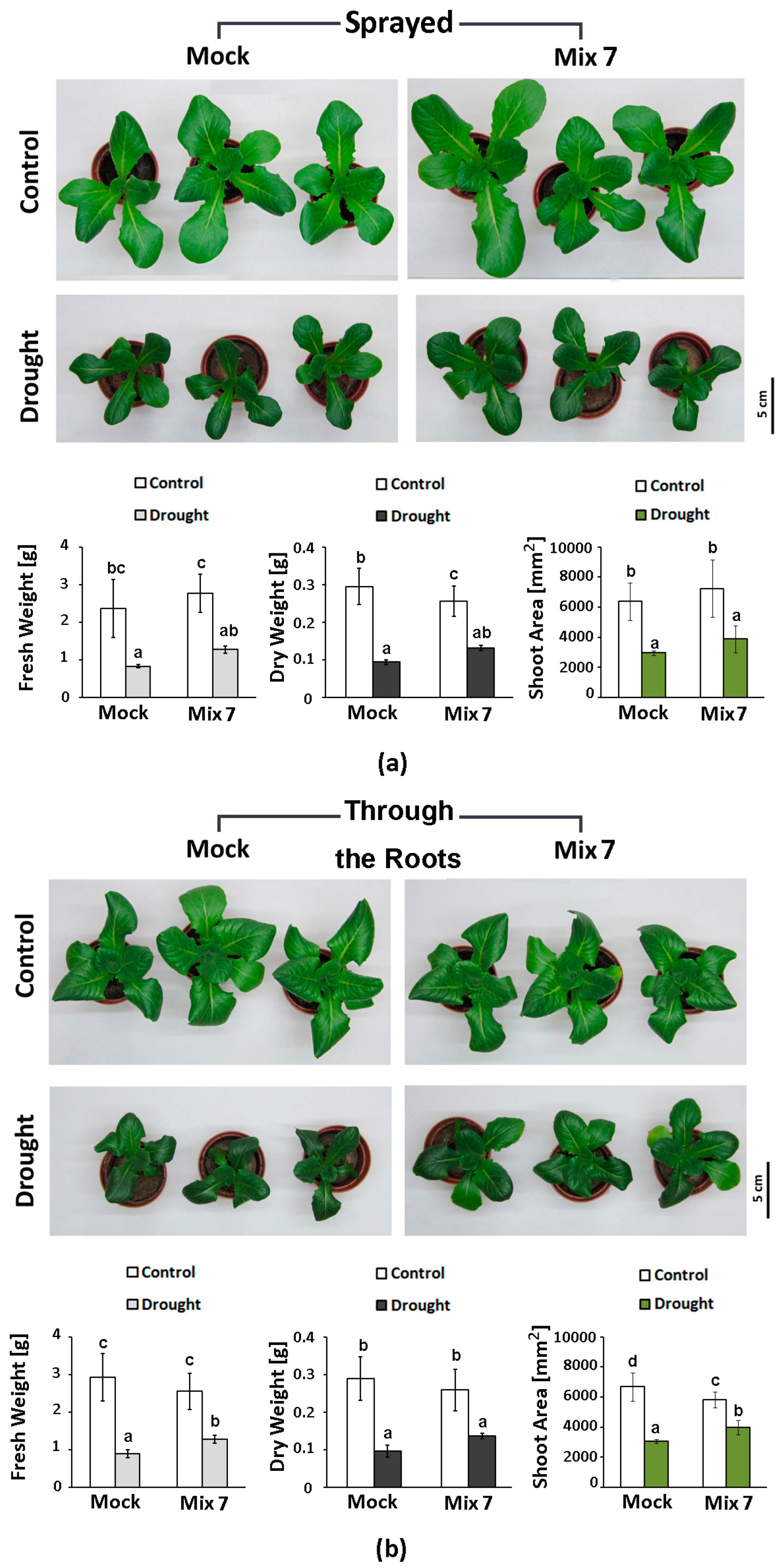
3.5. Testing of the Stress-Mitigating Potential of Mix 7 on Soil-Grown L. sativa Plants Subjected to Moderate Drought
3.6. Comprehensive Analysis of the Observed Drought-Mitigating Property of the Applied PGR Combination
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Commission. The European Green Deal. Available online: https://commission.europa.eu/strategy-and-policy/priorities-2019-2024/european-green-deal_en (accessed on 31 March 2024).
- European Commission. Farm to Fork Strategy Action Plan. Available online: https://food.ec.europa.eu/horizontal-topics/farm-fork-strategy_en (accessed on 31 March 2024).
- De Vasconcelos, A.C.F.; Chaves, L.H.G. Biostimulants and their role in improving plant growth under abiotic stresses. Biostimulants Plant Science. Intech. Open 2020, 88829, 1–14. [Google Scholar]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed]
- Cabello-Conejo, M.I.; Prieto-Fernández, Á.; Kidd, P.S. Exogenous treatments with phytohormones can improve growth and nickel yield of hyperaccumulating plants. Sci. Total Environ. 2014, 494–495, 1–8. [Google Scholar] [CrossRef]
- Jiang, K.; Asami, T. Chemical regulators of plant hormones and their applications in basic research and agriculture. Biosci. Biotechnol. Biochem. 2018, 82, 1265–1300. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 28, 259. [Google Scholar] [CrossRef]
- Khalid, M.F.; Huda, S.; Yong, M.; Li, L.; Li, L.; Chen, Z.-H.; Ahmed, T. Alleviation of drought and salt stress in vegetables: Crop responses and mitigation strategies. Plant Growth Regul. 2023, 99, 177–194. [Google Scholar] [CrossRef]
- Kaldate, R.; Singh, S.K.; Guleria, G.; Soni, A.; Naikwad, D.; Kumar, N.; Meshram, S.; Rana, M. Current approaches in horticultural crops to mitigate the effect of drought stress. In Stress Tolerance in Horticultural Crops; Rai, A.C., Rai, A., Rai, K.K., Rai, V.P., Kumar, A., Eds.; Woodhead Publishing: Sawston, UK; Elsevier Inc.: Amsterdam, The Netherlands, 2021; pp. 213–240. [Google Scholar]
- Movahedi, A.; Dzinyela, R.; Aghaei-Dargiri, S.; Alhassan, A.R.; Yang, L.; Xu, C. Advanced study of drought-responsive protein pathways in plants. Agronomy 2023, 13, 849. [Google Scholar] [CrossRef]
- Hoekstra, F.A.; Golovina, E.A.; Buintink, J. Mechanisms of plant desiccation tolerance. Trends Plant Sci. 2001, 6, 431–438. [Google Scholar] [CrossRef]
- Close, T.J. Dehydrins: A Commonalty in the response of plants to dehydration and low temperature. Physiol. Plant 1997, 100, 291–296. [Google Scholar] [CrossRef]
- González-Morales, S.; Solís-Gaona, S.; Valdés-Caballero, M.V.; Juárez-Maldonado, A.; Loredo-Treviño, A.; Benavides-Mendoza, A. Transcriptomics of biostimulation of plants under abiotic stress. Front. Genet. 2021, 12, 583888. [Google Scholar] [CrossRef] [PubMed]
- Vaseva, I.I.; Simova-Stoilova, L.; Kostadinova, A.; Yuperlieva-Mateeva, B.; Karakicheva, T.; Vassileva, V. Heat-stress-mitigating effects of a protein-hydrolysate-based biostimulant are linked to changes in Protease, DHN, and HSP gene expression in maize. Agronomy 2022, 12, 1127. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, L.; Chang, X.; Li, Q.; Abbasi, A.M. Influence of plant growth regulators on key-coding genes expression associated with phytochemicals biosynthesis and antioxidant activity in soybean (Glycine max (L.) Merr) sprouts. Int. J. Food. Sci. Technol. 2019, 54, 771–779. [Google Scholar] [CrossRef]
- Guerrero, F.; Mullet, J.E. Increased abscisic acid biosynthesis during plant dehydration requires transcription. Plant Physiol. 1986, 80, 588–591. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.A.; Santos, I.S.; Torres, M.E.L.; Cardon, C.H.; Caldeira, C.F.; Lima, R.R.; Chalfun-Junior, A. Drought and re-watering modify ethylene production and sensitivity, and are associated with coffee anthesis. Environ. Exp. Bot. 2021, 181, 104289. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Hassan, M.J.; Li, Z.; Peng, Y. Indole-3-acetic acid improves drought tolerance of white clover via activating auxin, abscisic acid and jasmonic acid related genes and inhibiting senescence genes. BMC Plant Biol. 2020, 20, 150. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.Y.; Azhar, N.; Hussain, M. Indole acetic acid (IAA) induced changes in growth, relative water contents and gas exchange attributes of barley (Hordeum vulgare L.) grown under water stress conditions. Plant Growth Regul. 2006, 50, 85–90. [Google Scholar] [CrossRef]
- Negi, S.; Ivanchenko, M.G.; Muday, G.K. Ethylene regulates lateral root formation and auxin transport in Arabidopsis thaliana. Plant J. 2008, 55, 175–187. [Google Scholar] [CrossRef]
- Ivanchenko, M.G.; Muday, G.K.; Dubrovsky, J.G. Ethylene–auxin interactions regulate lateral root initiation and emergence in Arabidopsis thaliana. Plant J. 2008, 55, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Ramireddy, E.; Hosseini, S.A.; Eggert, K.; Gillandt, S.; Gnad, H.; von Wirén, N.; Schmülling, T. Root engineering in barley: Increasing cytokinin degradation produces a larger root system, mineral enrichment in the shoot and improved drought tolerance. Plant Physiol. 2018, 177, 1078–1095. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.; Jervis, G.; Topping, J.F.; Chen, C.; Liu, J.; Lindsey, K. A predictive model for ethylene-mediated auxin and cytokinin patterning in the Arabidopsis root. Plant Commun. 2024, 100886. [Google Scholar] [CrossRef] [PubMed]
- De Vylder, J.; Vandenbussche, F.; Hu, Y.; Philips, W.; Van Der Straeten, D. Rosette tracker: An open source image analysis tool for automatic quantification of genotype effects. Plant Physiol. 2012, 160, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Boyes, D.C.; Zayed, A.M.; Ascenzi, R.; McCaskill, A.J.; Hoffman, N.E.; Davis, K.R.; Görlach, J. Growth stage-based phenotypic analysis of Arabidopsis: A model for high throughput functional genomics in plants. Plant Cell 2001, 13, 1499–1510. [Google Scholar] [CrossRef]
- Kramer, G.; Norman, H.; Krizek, D.; Mirecki, R. Influence of UV-B radiation on polyamines, lipid peroxidation and membrane lipids in cucumber. Phytochemistry 1991, 30, 2101–2108. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Bates, L.; Waldren, R.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Swain, T.; Goldstein, L. Methods in Polyphenol Chemistry; Pridham, J.B., Ed.; Pergamon Press: Oxford, UK, 1964; pp. 131–146. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–75. [Google Scholar] [CrossRef]
- Aebi, M. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Dias, I.; Costa, M. Effect of low salt concentration on nitrate reductase and peroxidase of sugar beet leaves. J. Exp. Bot. 1983, 34, 537–543. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase. Improved assay and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Analyt. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Vaseva, I.; Akiscan, Y.; Simova-Stoilova, L.; Kostadinova, A.; Nenkova, R.; Anders, I.; Feller, U.; Demirevska, K. Antioxidant response to drought in red and white clover. Acta Physiol. Plant. 2012, 34, 1689–1699. [Google Scholar] [CrossRef]
- Vaseva, I.; Anders, I.; Feller, U. Identification and expression of different dehydrin subclasses involved in the drought response of Trifolium repens. J. Plant Physiol. 2014, 171, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhashi, W.; Feller, U. Effects of light and external solutes on the catabolism of nuclear-encoded stromal proteins in intact chloroplasts isolated from pea leaves. Plant Physiol. 1992, 100, 2100–2105. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCq method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, X.; Dai, M. Improving crop drought resistance with plant growth regulators and rhizobacteria: Mechanisms, applications, and perspectives. Plant Commun. 2022, 3, 100228. [Google Scholar] [CrossRef] [PubMed]
- Brenya, E.; Dutta, E.; Herron, B.; Walden, L.H.; Roberts, D.M.; Binder, B.M. Ethylene-mediated metabolic priming increases photosynthesis and metabolism to enhance plant growth and stress tolerance. PNAS Nexus 2023, 2, pgad216. [Google Scholar] [CrossRef]
- Růzicka, K.; Ljung, K.; Vanneste, S.; Podhorská, R.; Beeckman, T.; Friml, J.; Benková, E. Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution. Plant Cell 2007, 19, 2197–2212. [Google Scholar] [CrossRef]
- Zdarska, M.; Cuyacot, A.R.; Tarr, P.T.; Yamoune, A.; Szmitkowska, A.; Hrdinova’, V.; Gelova’, Z.; Meyerowitz, E.M.; Hejatko, J. ETR1 integrates response to ethylene and cytokinins into a single multistep phosphorelay pathway to control root growth. Mol. Plant 2019, 12, 1338–1352. [Google Scholar] [CrossRef]
- Most, P.; Papenbrock, J. Possible roles of plant sulfurtransferases in detoxification of cyanide, reactive oxygen species, selected heavy metals and arsenate. Molecules 2015, 20, 1410–1423. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Gill, S.S.; Corpas, F.J.; Ortega-Villasante, C.; Hernandez, L.E.; Tuteja, N.; Sofo, A.; Hasanuzzaman, M.; Fujita, M. Editorial: Recent insights into the double role of hydrogen peroxide in plants. Front. Plant Sci. 2022, 13, 843274. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Kurepa, J.; Smalle, J.A. Auxin/cytokinin antagonistic control of the shoot/root growth ratio and its relevance for adaptation to drought and nutrient deficiency stresses. Int. J. Mol. Sci. 2022, 23, 1933. [Google Scholar] [CrossRef]
- Chaves, M.M.; Oliveira, M.M. Mechanisms underlying plant resilience to water deficits: Prospects for water-saving agriculture. J. Exp. Bot. 2004, 55, 2365e2384. [Google Scholar] [CrossRef]
- Hu, C.A.; Delauney, A.J.; Verma, D.P. A bifunctional enzyme (delta 1-pyrroline-5-carboxylate synthetase) catalyzes the first two steps in proline biosynthesis in plants. Proc. Natl. Acad. Sci. USA 1992, 89, 9354–9358. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.E.; Savouré, A.; László Szabados, L. Proline metabolism as regulatory hub. Trends Plant Sci. 2022, 27, 39–55. [Google Scholar] [CrossRef]
- Shrestha, A.; Cudjoe, D.K.; Kamruzzaman, M.; Siddique, S.; Fiorani, F.; Léon, J.; Naz, A.A. Abscisic acid-responsive element binding transcription factors contribute to proline synthesis and stress adaptation in Arabidopsis. J. Plant Physiol. 2021, 261, 153414. [Google Scholar] [CrossRef]
- Vaseva, I.I.; Simova-Stoilova, L.; Kirova, E.; Mishev, K.; Depaepe, T.; Van Der Straeten, D.; Vassileva, V. Ethylene signaling in salt-stressed Arabidopsis thaliana ein2-1 and ctr1-1 mutants—A dissection of molecular mechanisms involved in acclimation. Plant Physiol. Biochem. 2021, 167, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Ozfidan, C.; Turkan, I.; Sekmen, A.H.; Seckin, B. Abscisic acid-regulated responses of aba2-1 under osmotic stress: The abscisic acid-inducible antioxidant defence system and reactive oxygen species production. Plant Biol. 2012, 14, 337–346. [Google Scholar] [CrossRef]
- Xie, Y.; Sun, G.; Wang, L.; Tang, Y. Effects of spraying abscisic acid on growth and antioxidant enzyme of lettuce seedlings under salt stress. IOP Conf. Ser. Earth Environ. Sci. 2018, 199, 032015. [Google Scholar] [CrossRef]
- Tiwari, P.; Chakrabarty, D. Dehydrin in the past four decades: From chaperones to transcription co-regulators in regulating abiotic stress response. Curr. Res. Biotech. 2021, 3, 249–259. [Google Scholar] [CrossRef]
- Dalal, M.; Tayal, D.; Chinnusamy, V.; Bansal, K.C. Abiotic stress and ABA-inducible Group 4 LEA from Brassica napus plays a key role in salt and drought tolerance. J Biotech. 2009, 139, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhu, H.; Zhu, H.; Tao, Y.; Liu, C.; Liu, J.; Yang, F.; Li, M. Exogenous ABA enhances the antioxidant defense system of maize by regulating the AsA-GSH cycle under drought stress. Sustainability 2022, 14, 3071. [Google Scholar] [CrossRef]
- Sehar, Z.; Gautam, H.; Masood, A.; Khan, N.A. Ethylene- and proline-dependent regulation of antioxidant enzymes to mitigate heat stress and boost photosynthetic efficacy in wheat plants. J. Plant Growth Regul. 2023, 42, 2683–2697. [Google Scholar] [CrossRef]
- Hu, Z.; Fan, J.; Chen, K.; Amombo, E.; Chen, L.; Fu, J. Effects of ethylene on photosystem II and antioxidant enzyme activity in Bermuda grass under low temperature. Photosynth. Res. 2016, 128, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Leckie, C.P.; McAinsh, M.R.; Allen, G.J.; Sanders, D.; Hetherington, A.M. Abscisic acid-induced stomatal closure mediated by cyclic ADP-ribose. Proc. Natl. Acad. Sci. USA 1998, 95, 15837–15842. [Google Scholar] [CrossRef]
- Zarembinski, T.I.; Theologis, A. Ethylene biosynthesis and action: A case of conservation. In Signals and Signal Transduction Pathways in Plants; Palme, K., Ed.; Springer: Dordrecht, The Netherlands, 1994; pp. 343–361. [Google Scholar]
- Lakehal, A.; Bellini, C. Control of adventitious root formation: Insights into synergistic and antagonistic hormonal interactions. Physiol. Plant. 2019, 165, 90–100. [Google Scholar] [CrossRef]
- Lee, Z.H.; Hirakawa, T.; Yamaguchi, N.; Ito, T. The roles of plant hormones and their interactions with regulatory genes in determining meristem activity. Int. J. Mol. Sci. 2019, 20, 4065. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Mix 1 | Mix 2 | Mix 3 | Mix 4 | Mix 5 | Mix 6 | Mix 7 |
|---|---|---|---|---|---|---|---|
| ABA | + | - | + | + | + | + | + |
| KIN | + | + | + | − | − | − | + |
| NAA | − | + | − | + | + | − | + |
| ACC | − | - | + | + | − | + | + |
| Gene Name | Locus | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|---|
| Ls Mn-SOD | LOC111900556 | CACCCCAGTATTTGGATGGCT | CTCCCTCCCCCTATGTGCTA |
| Ls Fe-SOD | LOC111884404 | GGGAATCCATGCAACCAGGA | AAAACAAGCCAAACCCAGCC |
| Ls Cu/Zn-SOD | LOC111882980 | CACTCTTACAGACGCTTTGCG | ATGGTGCCACTAACACCCTC |
| Ls Catalase | LOC111878432 | GCCATGCTGAACAGTACCCT | TCTCTCTCCTGGCTGCTTGA |
| Ls APX2 | LOC111882573 | GACATCGGCGATCTTCTGGT | TCTCGAAGCTTCCTCTTCGC |
| Ls POX N1 | LOC111896420 | CTATGGTTGATATTGGCGTCGT | ACAAAGTCGGCCATTGGAGAT |
| Ls POX5 | LOC111879417 | GGTCGCTAAAGCCTACTCCC | ACTTGGGTTGTTTGCTGGTG |
| Ls18S RNA | AH001680 | CGGGTGACGGAGAATTAGGG | TACCTCCCCGTGTCAGGATT |
| Ls Actin-7 | LOC111882438 | CTGGTGATGGTGTCTCCCAC | GGCGAGCTTCTCCTTCATGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaseva, I.I.; Sergiev, I.; Todorova, D.; Urbutis, M.; Samuolienė, G.; Simova-Stoilova, L. Combined Pretreatment with Bioequivalent Doses of Plant Growth Regulators Alleviates Dehydration Stress in Lactuca sativa. Horticulturae 2024, 10, 544. https://doi.org/10.3390/horticulturae10060544
Vaseva II, Sergiev I, Todorova D, Urbutis M, Samuolienė G, Simova-Stoilova L. Combined Pretreatment with Bioequivalent Doses of Plant Growth Regulators Alleviates Dehydration Stress in Lactuca sativa. Horticulturae. 2024; 10(6):544. https://doi.org/10.3390/horticulturae10060544
Chicago/Turabian StyleVaseva, Irina I., Iskren Sergiev, Dessislava Todorova, Martynas Urbutis, Giedrė Samuolienė, and Lyudmila Simova-Stoilova. 2024. "Combined Pretreatment with Bioequivalent Doses of Plant Growth Regulators Alleviates Dehydration Stress in Lactuca sativa" Horticulturae 10, no. 6: 544. https://doi.org/10.3390/horticulturae10060544