Abstract
As a consequence of climate change, temperature and rainfall regimes are being modified, threatening the survival of the current gardening concept in many areas of the world. This situation highlights the need to investigate the potential of other species, especially in more sensitive areas such as the Mediterranean Basin. The aim of this research is to study the resilience of adapted species to promote sustainable gardening. To achieve this, seven species belonging to three families (Amaranthaceae, Lamiaceae and Asteraceae) used or potentially used as ornamentals were selected to compare their tolerance at the germinative stage to different levels of temperature (10 °C to 35 °C) and water stress created by PEG 6000 (−0.10 MPa to −2.5 MPa). Germination percentage, mean germination time, base temperature, thermal time, base water potential and hydrotime were calculated. The results showed a good response to high temperature and low osmotic potential in most of the species, and germination even increased under certain stress levels. Salsola oppositifolia presented by far the best results in terms of germination under high-water-stress conditions and the second best at high temperatures. The extraordinary response of Celosia argentea, an alien species, highlighted the risk of it becoming an invasive species.
1. Introduction
Human activities, principally through emissions of greenhouse gases, are unequivocally causing global warming, with the global surface temperature reaching 1.1 °C above the 1850–1900 average in 2011–2020 [1]. These effects are projected to be especially enhanced in the Mediterranean basin, where an average temperature increase of between 2 °C and 3 °C is expected by 2050, compared to between 1.3 °C and 1.8 °C, which is expected globally [2,3]. The climate change associated involves other negative impacts in this area such as water scarcity, extreme events (heat waves, drought periods, etc.), displacement of the rainy season, and disturbances to environmental integrity (biodiversity loss, desertification, etc.) [4].
The distribution of plants in natural ecosystems is closely related to environmental conditions since they have specific temperature and humidity requirements to survive and propagate. Therefore, their distribution range is changing depending on the environment due to climate change [5,6]. These changes also affect agronomic species and those species that are traditionally used in gardening.
Urban gardens can play an important role in mitigating climate change in cities by reducing temperatures and creating thermally healthy spaces [7]. Traditionally, the criteria for selecting species used in gardening have been influenced by social more than by ecological factors, remaining the dominant model inherited from the traditional “garden à la française” [8]. Saurí et al. [9] identified in the metropolitan area of Barcelona that the average water consumption in detached houses was consistently larger than in higher-density residential buildings due to the water consumption of gardening and other recreational activities. Furthermore, in recent years, the increase in temperatures and the lower water availability have been compensated for with supplementary irrigation. Hence, adapting gardening practices and species for future sustainability purposes is a challenge for gardening today. The selection of species resistant to high temperatures, with lower maintenance needs and that also fulfil other ecosystem services, is essential in a climate change scenario.
In this sense, xeriscaping is a valuable method that emerged for developing water-wise landscapes and encourages efficient landscape maintenance, reducing or eliminating supplemental water for irrigation through different strategies such as drought-tolerant plant selection [10]. This approach is based on thoughtful choices that balance aesthetic appeal with water conservation and offer a sustainable alternative to traditional landscaping practices, particularly in regions facing water scarcity challenges [11]. Therefore, the environmental benefits of switching from traditional gardening to xeriscaping are diverse, such as reduced water consumption, improved biodiversity, protection of soil health, reduced use of chemicals, and promoting resilience to climate change, but also lower maintenance costs [12,13,14,15].
Germination is one of the most sensitive stages of a plant’s life cycle and one of the most susceptible to various types of stress. A plant resistant at this stage is more likely to overcome the rest of the phases of its life cycle [16,17]. For this reason, the aim of this research is to study the resilience of ornamental and potentially ornamental species to conditions arising from climate change at the germination stage in order to promote sustainable gardening. Mediterranean flora, especially plants living in the most arid environments, are adapted to the characteristics of the Mediterranean climate and should therefore be potentially more suitable for use in sustainable and environmentally responsible gardening in harmony with current and future environmental conditions. To achieve this objective, seven species belonging to three families (Amaranthaceae, Lamiaceae and Asteraceae) were selected, three of them already used in gardening and four belonging to the native flora. Their tolerance to different levels of temperature and water stress was compared to select those species that are more resistant and adaptable to high temperatures and lower water supply. Therefore, this study represents a valuable approach to the selection of tolerant plants for xeriscaping.
2. Materials and Methods
Seven species belonging to three families (Amaranthaceae: Salsola oppositifolia Desf., Celosia argentea L.; Lamiaceae: Phlomis purpurea L., Thymbra capitata (L.) Cav., Salvia sclarea L. and Asteraceae: Pallenis maritima (L.) Greuter, Leucanthemum vulgare Lam.) (Figure 1), used or with potential use as ornamentals, were selected to compare their tolerance to different levels of temperature and water stress. The assays were carried out in 2023 in the Germplasm bank of the Valencian Botanical Garden (Universitat de València, Spain). Most of the seeds were provided by two plant nurseries: Semillas Silvestres S.L. (Cordoba, Spain) and Verdecora (Valencia, Spain), except for S. oppositifolia. Seeds of S. oppositifolia were collected at the time of natural dispersal from the wild population (Alicante, Spain), cleaned up and stored in paper bags at 20 °C and 40–50% RH, until used in the germination tests. All plant debris was removed from these non-nursery seeds.

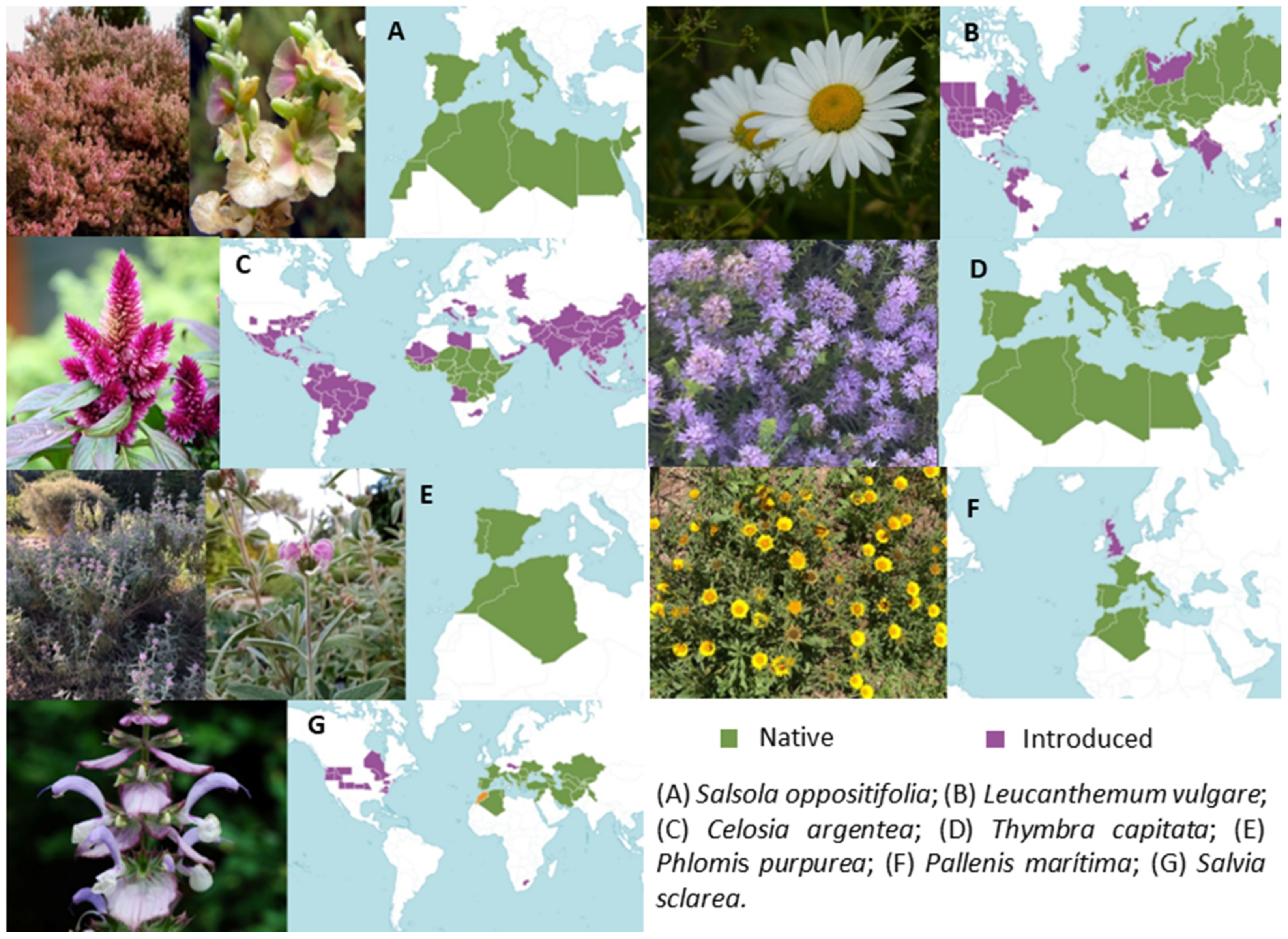
Figure 1.
Distribution maps of the species studied. (A) Salsola oppositifolia; (B) Leucanthemum vulgare (by Jamain, CC); (C) Celosia argentea (modified from Hedwig Storch, CC); (D) Thymbra capitata; (E) Phlomis purpurea; (F) Pallenis maritima; (G) Salvia sclarea (by Pancrat, CC). Green shading on the maps indicates the native range of the species; purple shading indicates the range where they have been introduced. All the maps were extracted from Plants of the World Online (Kew, Royal Botanic Gardens; https://powo.science.kew.org, accessed on 4 April 2023).
2.1. Germination Assays
Seed germination assays were performed using four replications of 25 seeds per treatment and species for different conditions of temperature and water stress. Tests were conducted on 55 mm diameter Petri dishes in 0.6% agar (Scharlau Chemie S.A., Barcelona, Spain) kept in temperature-controlled cabinets. Illumination was provided by daylight fluorescent tubes with a 12 h photoperiod and a mean irradiance of 100 μmol m−2s−1. The germination process was evaluated for 30 days. Germinated seeds were counted daily. None of the species in this study showed physiological dormancy, so no pre-treatment was necessary. A temperature screening was carried out including six constant temperatures: 10, 15, 20, 25, 30 and 35 °C (MIR-154, Sanyo Electric Co. Ltd., Tokyo, Japan; Grow-600, Climaslab S.L., Barcelona, Spain). As a result of the screening, the optimal condition, 20 °C, was selected to perform the water stress experiments.
The effect of water stress on germination was studied by creating a low-osmotic-potential environment with increasing concentrations of polyethylene glycol-6000 (PEG-6000; Scharlau Chemie S.A., Barcelona, Spain). The osmotic potentials selected were −0.1, −0.2, −0.3, −0.4, −0.6, −1, −1.6, −2 and −2.5 MPa. The concentrations of PEG were calculated following [18] Michel and Kaufmann to obtain the specific osmotic potentials (Ψ) for tests at 20 °C. The seeds were sown with two 4.6 mm diameter filter papers. To each plate, 2 mL of the corresponding solution was added. The plates were stored in zippered plastic bags to prevent evaporation.
In order to know the influence of temperature and lack of water, germination percentage and mean germination time (MTG) were calculated. MTG was determined according to the formula MGT = ∑ Dn/∑n, where D represents the number of days from the beginning of the germination test and n is the number of seeds newly germinated on day D [19]. To better compare the behaviour of the different species regarding the influence of water stress in germination, “change in germination percentage” was calculated as follows: CGP = −[1 − (N° of germinated seeds in the salt treatment/N° of germinated seeds in the control)] × 100. CGP is based on the reduction in germination percentage (RGP) proposed by El-Madidi et al. [20], which only considers a negative effect of treatment.
2.2. Germination Modelling
To predict the germination rate under fluctuating environmental conditions, base temperature (Tb), thermal time (S), base osmotic potential (Ψb) and hydrotime (ƟH) were also calculated. Base temperature (Tb) and thermal time (S) determine the relationship between temperature, time and germination behaviour and compare thermal responses between species [21,22]. The base temperature corresponds to the theoretical temperature in degrees Celsius, below which germination is not possible, and the thermal time reflects the thermal requirements in temperature per time to achieve germination. Both parameters were calculated from linear regression on the graph of the inverse time to germination, expressed as the mean germination time (1/TMG), as a function of temperature. The germination time curves for the different PEG concentrations were analysed by applying the hydrotime model, which determines the relationship between osmotic potential, rate and germination response. The hydrotime model provides an approach to predict seed germination in soil under water deficit conditions, according to the equation of Gummerson [23]. The base water potential (Ψb) was calculated by extrapolating the regression line on the graph of the inverse of radicle emergence time, expressed as the mean germination time (1/MGT), versus osmotic potential. Hydrotime (ƟH) was estimated as the inverse of the slope of these regression lines [24,25]. The base values should be treated with caution because they were obtained from extrapolation beyond the range of experimental conditions. These calculated values were taken as theoretical figures for comparisons between species.
2.3. Statistical Analysis
All germination data for analysis of temperature and water stress effects were statistically assessed using the statistical software SPSS 28.0. The significance of the differences among germination treatments was tested by applying a nonparametric test, Kruskal–Wallis one-way ANOVA with a 95% confidence interval. One-way ANOVA is a very robust method that provides good approximations for small samples when model assumptions are not fully satisfied [26]; therefore, post hoc multiple-comparison tests associated with the ANOVA analysis were conducted to estimate homogeneous groups. Tukey’s test was used for multiple comparisons among temperature groups and Dunnet’s t-test for PEG-treated groups, which were compared with the control.
3. Results
3.1. Temperature Assays
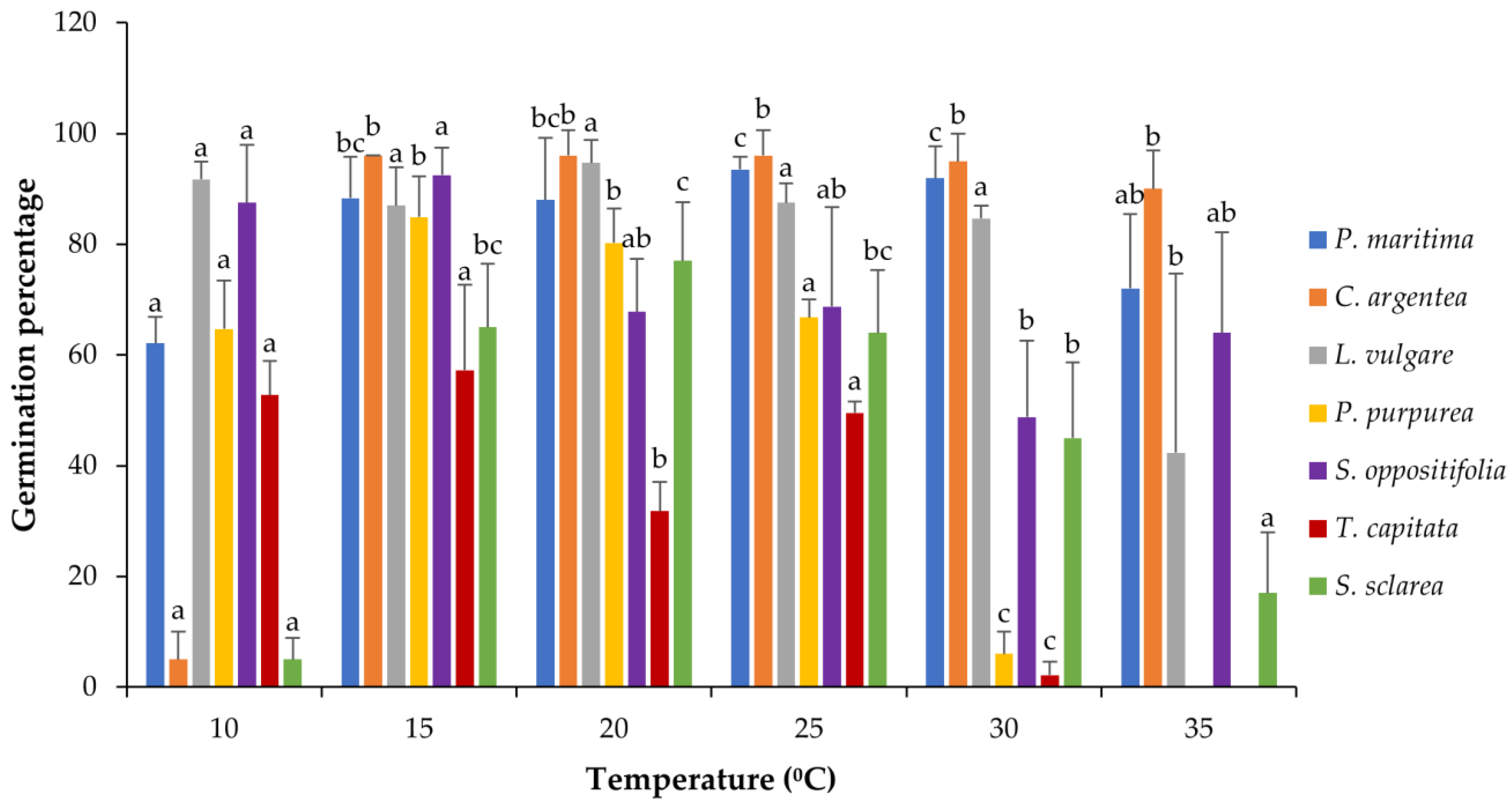
Most of the species revealed a good tolerance to high temperatures and reached high levels of germination percentage even at 35 °C, especially C. argentea, P. maritima and S. oppositifolia (Table S1). Nevertheless, all the species showed significant differences among treatments (Kruskal–Wallis one-way ANOVA), although the germinative response temperature was different depending on the species (Figure 2 and Figure A1). The response in P. maritima was around 90% in the range of 15–30 °C, being lower in the most extreme temperatures tested, 10 and 35 °C, with 62% and 72% of germination, respectively. Salsola oppositifolia yielded a high percentage of germination in all treatments, with the highest percentage around 90% for 10 °C and 15 °C and the lowest 48% for 30 °C. In the case of C. argentea, the germination was nearly 100% in most temperatures except at 10 °C, where only 5% of seeds germinated. The other species that presented such low germination at 10 °C was S. sclarea, for which the optimum was 20 °C (77%), and as temperatures moved away from the optimum, the percentage decreased progressively: around 65% at 15 °C and 25 °C, 45% at 30 °C, and 17% at 35 °C. This pattern was similar for P. purpurea but with less tolerance to high temperatures: the maximum was at 15 °C with almost 85% of germination, the minimum at 30 °C with 6%, and no response to 35 °C. Thymbra capitata was the least tolerant to high temperatures and also failed to germinate at 35 °C. In general, the germination for this species was lower than for the rest, with values around 50% in the best cases (10 °C, 15 °C and 25 °C) and 2% in the worst.
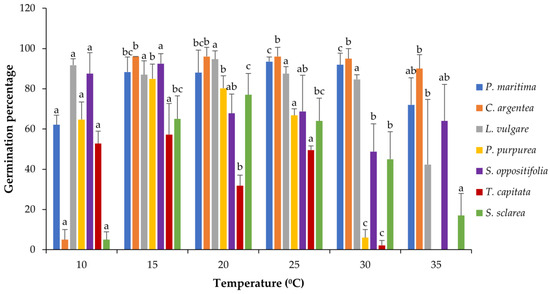
Figure 2.
Germination response to temperature for all the species depicted as mean bars with the standard deviation (+SD). The same letters indicate homogenous intra-species groups (Tukey p < 0.05).
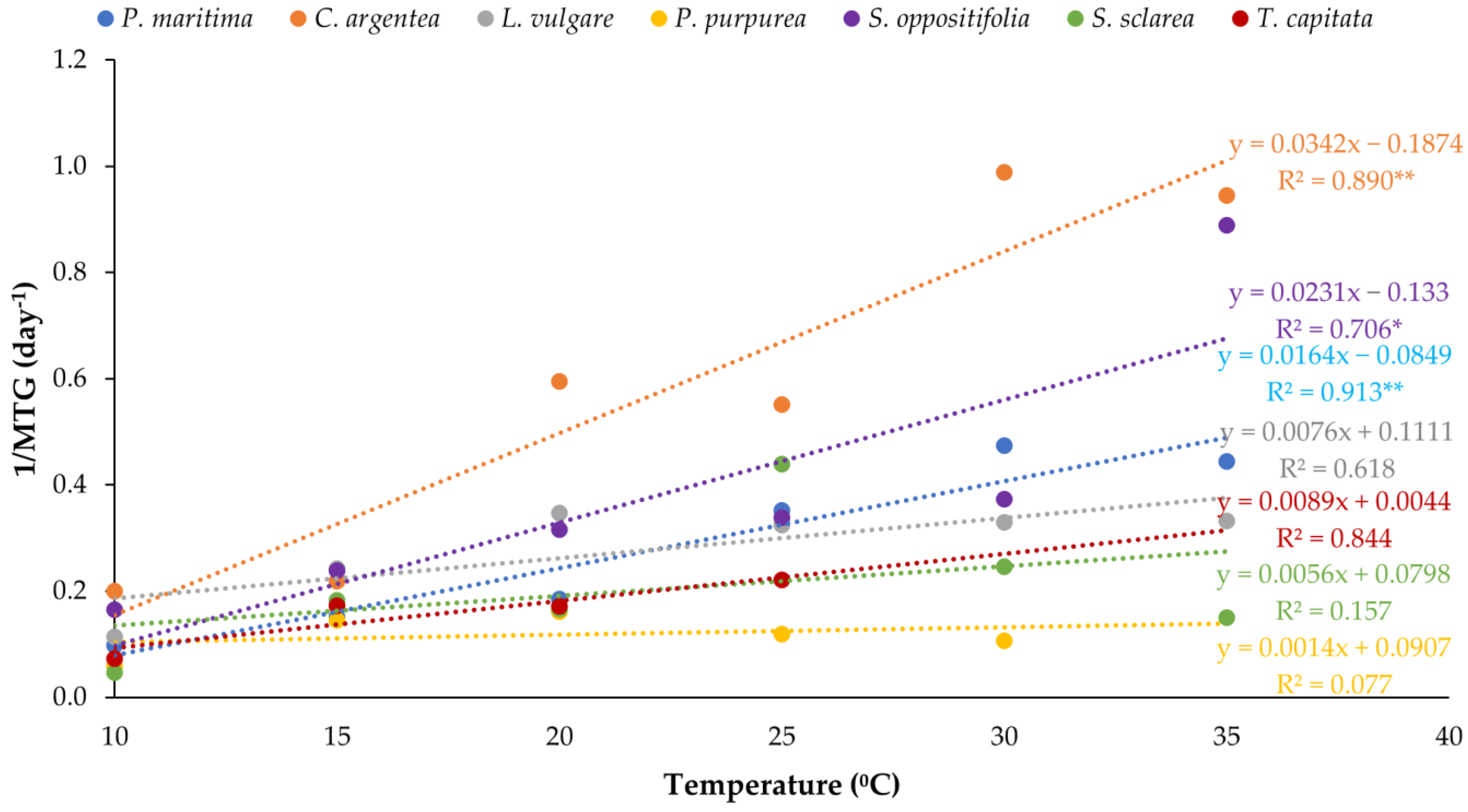
With regard to the velocity of germination, C. argentea, followed by S. oppositifolia, germinated earlier at high temperatures, but all the species also showed significant differences among treatments. Table 1 summarises the MGT for all the species and temperatures tested. As a rule, velocity increased with temperature, although at 35 °C, S. sclarea decreased significantly and P. purpurea and T. capitata were unable to germinate. All the species presented significant differences between the velocity at 10 °C and the rest of the temperatures, but the velocity depended on the species and differences in terms of germination velocity were found. These results are clearly shown in the representation of the inverse of the MGT values. Figure 3 represents the linear regression adjusted for the 1/MGT values for each of the species to fit the linear regression for thermal time constant calculation. The regression line of C. argentea is the upper one, showing that it was the most competitive species in terms of germination velocity. Salsola oppositifolia occupied the second position as a competitor in velocity. Thymbra capitata, S. sclarea and P. purpurea were the least competitive species at the temperatures tested. Base temperature (Tb) and thermal time (S) were obtained from regression lines (Table 2). These data numerically reflect the exposed results, although these calculated values should be considered as theoretical figures. Celosia argentea and S. oppositifolia were shown to require the least accumulation of heat units to germinate with the lowest thermal times and therefore had the highest competitiveness (as shown by the linear regressions).

Table 1.
Mean germination time (MGT) in days under the range of studied constant temperatures (mean ± s.d.) in a 12/12 h photoperiod. The same letters indicate homogeneous groups (Tukey p < 0.05). When germination percentages were lower than 5%, MGT was not calculated. Over 5% germination occurred on only one plate of C. argentea at 10 °C and of P. purpurea at 30 °C; therefore, Tukey’s test cannot be performed for these treatments.
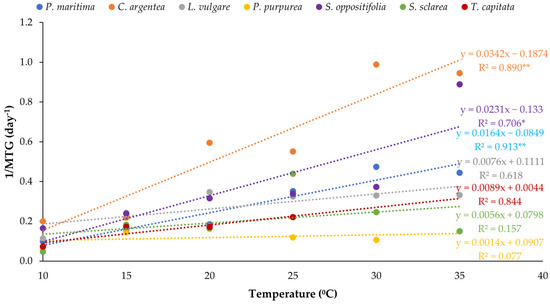
Figure 3.
Relationships between temperature and germination rate (1/MGT) for each of the species. The regression lines are represented for the calculation of the thermal time requirements. Asterisks indicate three levels of significance of R2 (* p < 0.05, ** p < 0.01). The regression lines that are above correspond to the most competitive population in a range of temperatures.
Table 2.
Theoretical base temperature (Tb) and thermal time (S) calculated for each species.
3.2. Water Stress Assays
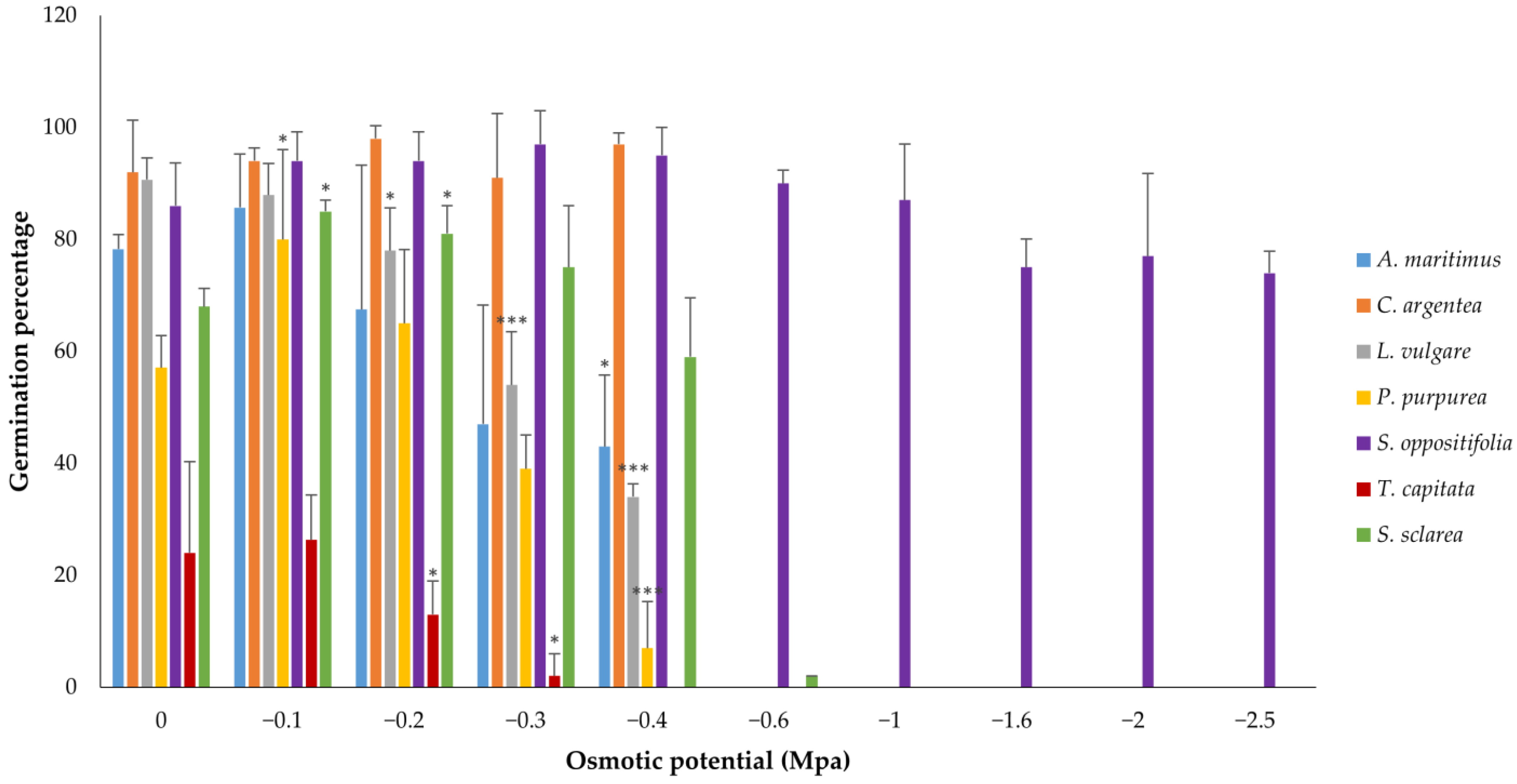
Under the water stress conditions generated by the different concentrations of PEG, S. oppositifolia presented the highest tolerance to hydric stress, reaching high germination percentages at all osmotic potentials (Table S2). Nevertheless, all the species, except C. argentea, showed significant differences among treatments (Kruskal–Wallis one-way ANOVA) and all, except L. vulgare, presented a positive CGP at least at −0.1 Mpa. However, a high variability in the germinative response was found depending on the species (Figure 4 and Figure A2, Table 3). Salsola oppositifolia reached a 74% rate of germination even in the −2.5 Mpa treatment, whereas most species did not germinate beyond −0.4 Mpa. Furthermore, although there were significant differences among all treatments in this species, no significant differences were found between each of the treatments and the control, and the CGP increased until −0.6 MPa. Celosia argentea and S. sclarea presented similar responses when comparing between treatments with CGP increases in some of them, but S. sclarea obtained a significantly higher response at −0.1 and −0.2 Mpa. Phlomis purpurea reached the maximum of germination at −0.1 MPa and, although the following treatments yielded decreases in the germination percentage, the CGP with respect to the control was still positive at −0.2 Mpa. This decreasing pattern was repeated, but from the control, by L. vulgare and T. capitata with significant differences as water stress increased. The former species presented the lowest germination percentages (maximum around 25%) and did not germinate at −0.4 Mpa, although CGP was positive at −0.1 Mpa.
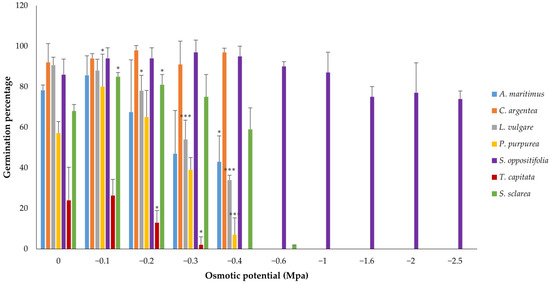
Figure 4.
Germination response to ascending concentrations of PEG for all the species, depicted as mean bars with standard deviations. Asterisks indicate three levels of significance with respect to the control (Dunnett: * p < 0.05, *** p < 0.001). Over 5% germination occurred on only one plate of S. sclarea at −0.6 Mpa; therefore, Dunnett’s t-test cannot be performed.
Table 3.
Change in germination percentage (CGP) under the range of PEG concentrations in a 12/12 h photoperiod with respect to the control mean. Positive values indicate an increase in germination and negative values indicate a decrease.
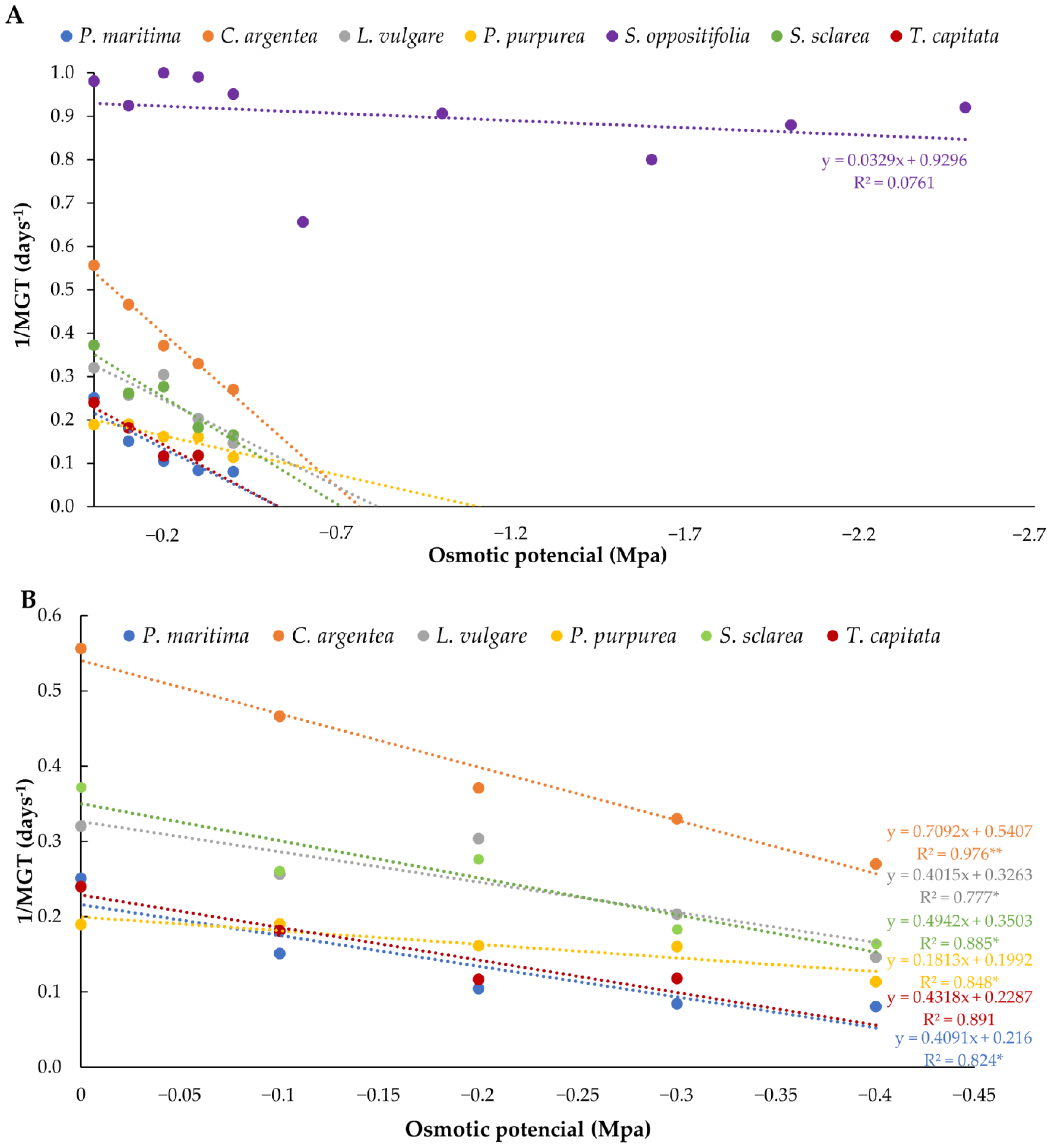
Concerning germination velocity in water stress assays, S. oppositifolia also showed the highest velocities at all osmotic potentials tested. The rest of the species revealed significant differences among treatments and velocity decreased as water potential decreased (Table 4). Pallenis maritima was the species that showed significant differences in velocity compared to the control at higher water potential (−0.1 Mpa), followed by T. capitata and C. argentea (−0.2 Mpa), then by S. sclarea (−0.3 Mpa), and L. vulgare and P. purpurea (−0.4 Mpa). Using the velocity data, the hydrotime model was applied to predict the germination of species under reduced water potentials and to determine the more drought-tolerant of them. Figure 5 shows the inverse of the MTG values obtained at the different osmotic potentials tested. The purpose of this figure is to fit the linear regression for the calculation of the hydrotime, constant for each species. In this sense, the most competitive species in drought conditions was S. oppositifolia, followed by C. argentea, which demonstrated a decreasing pattern of germination. Pallenis maritima and T. capitata were the least competitive, although P. purpurea took longer to germinate in control conditions. Base water potential (Ψb) and hydrotime (ƟH) were obtained from regression lines (Table 5) and numerically reflect these results: S. oppositifolia was by far the most tolerant to water stress conditions with the lowest theoretical base water potential (−28.26 MPa) and hydrotime (30.40 MPa × day). It was followed by P. purpurea, with a base water potential of −1.10 MPa and a hydrotime of 5.52 MPa × day. On the other hand, T. capitata showed less competitive values (Ψb = −0.53 MPa; θ = 2.32 MPa).
Table 4.
Mean germination time (MGT) in days under the range of PEG concentrations in a 12/12 h photoperiod. Asterisks indicate three levels of significance with respect to the control (Dunnett: * p < 0.05, ** p < 0.01, *** p < 0.001). When germination percentages were lower than 5%, the MGT was not calculated. Over 5% germination occurred on only one plate of T. capitata at 0.3 MPa; therefore, Dunnett’s test could not be performed for these treatments.
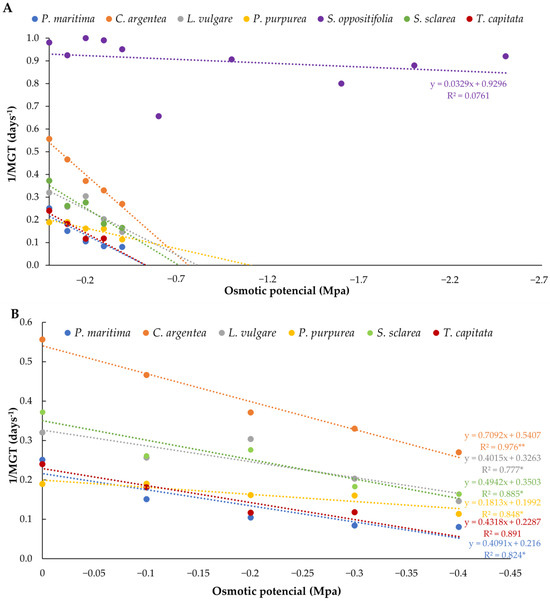
Figure 5.
Relationships between PEG concentration and germination rate (1/MGT) for each of the species: (A) all the species; (B) all the species except S. oppositifolia. The regression lines are represented for the calculation of the hydrotime requirements. Asterisks indicate three levels of significance of R2 (* p < 0.05, ** p < 0.01). The regression lines that are above correspond to the most competitive population in a range of osmotic potentials.
Table 5.
Base water potential (Ψb) and hydrotime (ƟH) calculated for each population through the linear regressions.
4. Discussion
Changes in environmental conditions have especially affected the Mediterranean basin in recent decades as one of the most vulnerable territories, characterised by a long, hot and dry summer period. This area is expected to remain among the most affected regions, with a significant increase in temperature and the duration of heat waves, as well as a decrease in the amount of precipitation and a change in rainfall distribution patterns [27,28,29]. It is not only wild plants that are affected by these changing environmental conditions but also the design of gardening and urban landscapes, and challenges are posed to plants in adapting to them [30]. This aspect occurs simultaneously with a trend towards increasing green areas in cities, which would imply a growth in water needs for irrigation [31].
This situation requires, in Mediterranean climate zones, the choice of plants resistant to high temperatures and water stress. In this sense, xeriscaping, based on the use of plants with greater resistance and better efficiency in the use of water resources, is a good option due to its environmental and economic benefits. Native plants from arid and saline areas of the Mediterranean basin represent an important source of resources for research, since they can potentially be used as ornamentals due to their ability to overcome adverse environmental conditions through the development of diverse adaptation mechanisms including morphological, physiological and biochemical modification patterns [13,14,15].
This study focuses on the proposal of alternative species to those traditionally used as ornamentals, based on their germination response. Germination is a crucial stage in the life of a plant since the survival of the new seedlings depends on their precision and adaptation to environmental conditions. Furthermore, in many cases, the germination response is related to the behaviour during the juvenile and adult stages of the plant [16,17], being a good predictor for selecting plants at the first level. This study evidences considerable differences in germination response under thermal and water stress among the seven studied species.
Temperature and water potential are key factors in controlling germination and seedling success [32,33] in unpredictable environments such as Mediterranean territories. Germination depends directly on temperature because it controls the water absorption of seeds and the biochemical pathways involved in the metabolism of the germination process [34]. The analysis of temperature tolerance showed some differences between the species studied, with the Amarantaceae being the most tolerant and Lamiaceae the least. In this sense, T. capitata was the most sensitive species since it barely germinated above 25 °C. This species was also studied for its potential use in xeriscaping [35], finding high germination percentages (80%) in laboratory experiments, but only 20% in greenhouse conditions and null results in hydroseeding experiments. On the contrary, Celosia argentea is a not frost-tolerant warm-season species [36], affected by low temperatures [37], especially in the germination period. However, this is the most tolerant species, reaching higher percentages than the rest of the species at the highest temperatures experienced and overcoming the germination percentages obtained by other authors in some of the conditions tested [34,38]. Generally, almost all species reached germination percentages above 50% in a wide range of temperatures, and some of them even reached values that exceeded 80% at the lowest temperatures tested. These results agree with those of other wild and ornamental species from arid zones, proving that temperature is not a limiting factor for germination but rather water availability [39,40,41]. The ability to germinate in a wide range of temperatures is important for plants with potential ornamental use because it allows germination and seedling establishment at any time of the year, as long as water is available [42] and helps them succeed in environments with fluctuating temperatures [43]. The capacity to germinate at low temperatures was also demonstrated in Hypericum ericoides, a wild plant with ornamental potential growing in Mediterranean areas. It has been postulated as an adaptive mechanism to avoid the adverse conditions of hot summers and germinate in autumn or spring when soil moisture conditions are most suitable for seedling survival [44].
When comparing competitiveness according to germination rate, the most competitive species at almost all the temperatures tested was C. argentea followed by S. oppositifolia at 15 °C and above. The species with the lowest Tb, P. purpurea, is the most sensitive to an increase in temperature due to global warming. As evidenced for other species, an inverse correlation between Tb and thermal time means that adaptation to a higher Tb corresponds to lower thermal time and more efficient germination [43]. As these authors highlighted when comparing different groups of species, the lowest thermal times were recorded for ornamental and other species living in harsh habitats.
Concerning drought tolerance, when seeds were subjected to decreasing values of water potential, most of the plants in this study were equally or more sensitive to water stress at the germination stage than other ornamental and invasive species [41,44,45]. None of the species, except S. oppositifolia, were able to germinate at osmotic potentials lower than −0.4 MPa. This species germinates at the lowest potentials tested, with results that equal or exceed those obtained in the control, above the percentages of some species of the same genus [46] and even those of some of the most tolerant halophytes [47]. These results were evidenced in other species of the same genus living in desert areas under saline soils that also germinate at −2.4 Mpa, although with percentages lower than 15% [48]. This capacity was also confirmed by the lower Ψb values of S. oppositifolia, as proof of a high resistance to drought in the germination phase that allows adaptation to harsh environments [49]. Furthermore, when competitiveness, based on germination speed, was evaluated, this species was by far the most competitive of all the species, while the most sensitive and the least competitive at low potentials were P. maritima and T. capitata.
The CGP values demonstrate that most of the species studied improve germination when subjected to low levels of stress. This is of interest not only because it allows its use in sustainable gardens, reducing water consumption, but also in plant propagation in nurseries for ornamental purposes.
In light of the results achieved, C. argentea and S. oppositifolia are the two species with the best results to be proposed as ornamental, as the most competitive species concerning temperature and water stress tolerance.
Celosia argentea, the species most tolerant to an increase in temperature, is the second most competitive at the highest osmotic potentials, but this competitiveness decreases when the osmotic potential values are low. This behaviour, typical of a species with a tropical distribution, makes it unsuitable for cultivation in water-scarce areas. Likewise, the use of a non-native species tolerant to heat and drought conditions and the rapid germination observed in this species are predictors of invasiveness that make it problematic because of its invasive potential and ability to alter natural environments [50,51,52].
The germination percentage of S. oppositifolia did not show differences between the control and the lowest potentials tested. Native species tolerant to water and thermal stress, such as the one mentioned above, constitute an important resource for use in sustainable gardening projects [53]. Furthermore, the selection of species tolerant to desiccation in climates such as the Mediterranean, with significant summer aridity, reduces irrigation needs during a significant part of the year [31]. This allows us to respond to the current challenge of the limited water availability for irrigating gardens during periods of drought. In any case, once its tolerance in the germination stage has been demonstrated as a first screening level, further studies confirming these results in the growth phase are being carried out in order to endorse its suitability.
5. Conclusions
Salsola oppositifolia proved to be the best candidate for sustainable xeriscaping in the face of climate change in the Mediterranean area due to its high resilience to water stress and high temperatures. C. argentea also demonstrated an excellent response, but it is a non-native organism in the Mediterranean area, making it a potential invasive species if not sufficiently controlled. However, P. maritima, S. sclarea and L. vulgare may also be used in sustainable gardening, where water stress is maintained at low to medium levels. Finally, the use of P. purpurea and T. capitata in xeriscaping should be considered on a case-by-case basis due to their limited germination at high temperatures. Nevertheless, it is important to note that these findings are drawn from the results of the germination phase and, although this stage is crucial in the life cycle of the plant, further experiments in the growth phase are underway to confirm them.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae10060620/s1, Table S1: Raw germination percentages for the temperature assays of all the species; Table S2: Raw germination percentages for the water stress assays of all the species.
Author Contributions
Conceptualization, P.S. and M.I.M.-N.; methodology, E.E., P.S. and M.I.M.-N.; validation, E.E., P.S. and M.I.M.-N.; formal analysis, R.M. and M.I.M.-N.; investigation, R.M. and P.S.; resources, P.S. and E.E.; data curation, P.S. and M.I.M.-N.; writing—original draft preparation, P.S., M.I.M.-N. and R.M.; writing—review and editing, E.E., P.S. and M.I.M.-N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in the study are included in the Supplementary Material (Tables S1 and S2 contain the original germination percentages of the study) further inquiries can be directed to the corresponding author/s.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
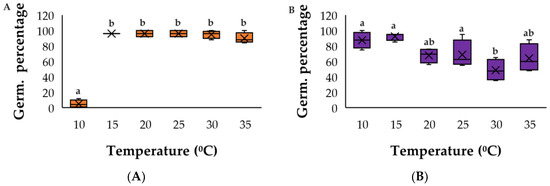
Figure A1.
Germination response to temperature for (A) C. argentea and (B) S. oppositifolia as the most competitive species. The same letters indicate homogenous groups (Tukey p < 0.05).
Figure A1.
Germination response to temperature for (A) C. argentea and (B) S. oppositifolia as the most competitive species. The same letters indicate homogenous groups (Tukey p < 0.05).
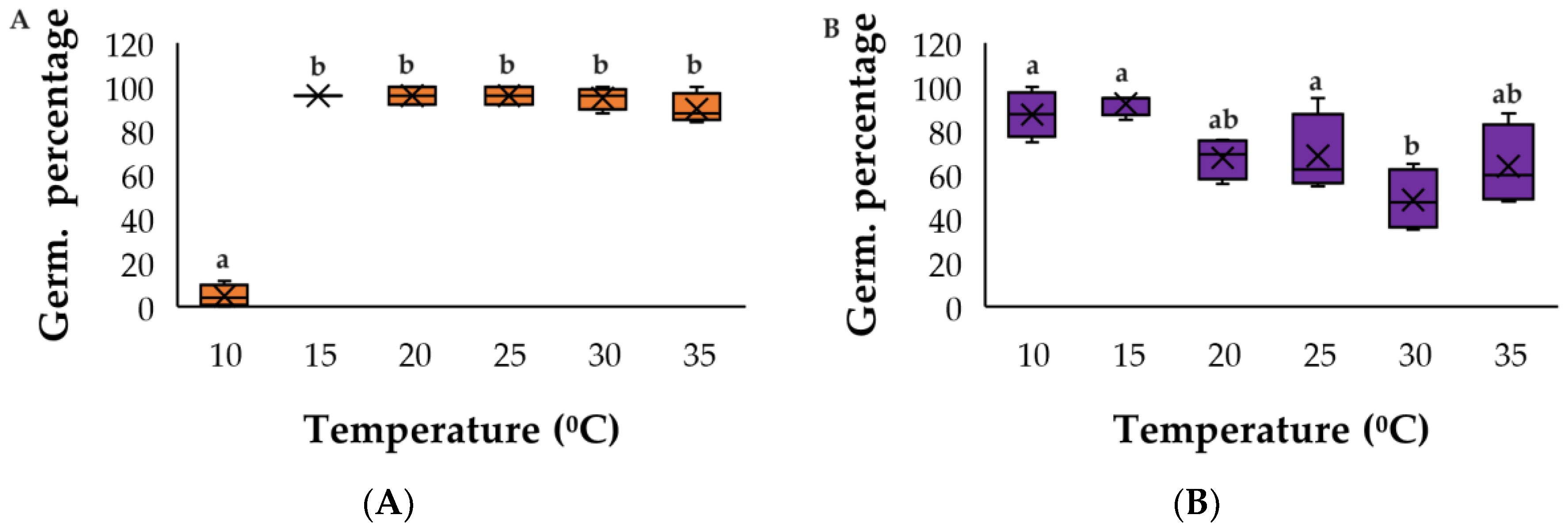
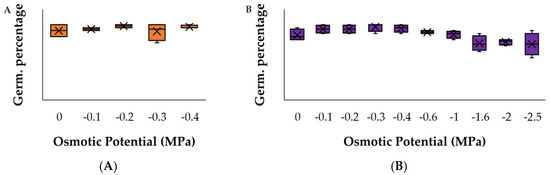
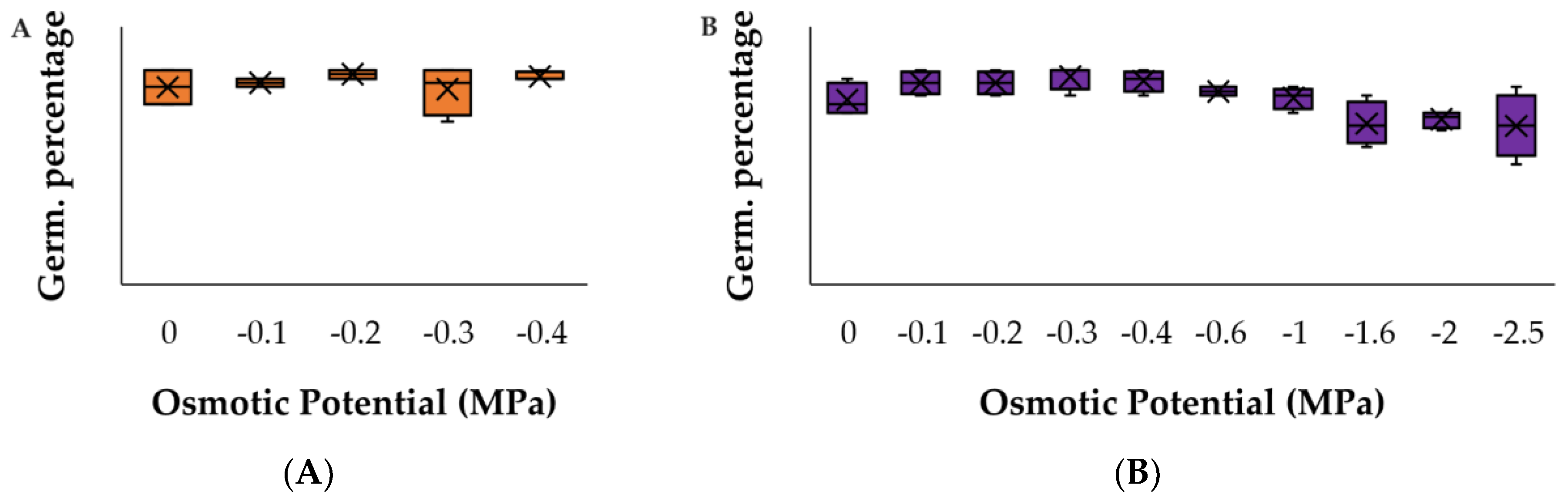
Figure A2.
Germination response to PEG for (A) C. argentea and (B) S. oppositifolia as the most competitive species. There were no significant differences with respect to the control (p > 0.05).
Figure A2.
Germination response to PEG for (A) C. argentea and (B) S. oppositifolia as the most competitive species. There were no significant differences with respect to the control (p > 0.05).
References
- IPCC. Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Lee, H., Romero, J., Eds.; IPCC: Paris, France, 2023; Available online: https://www.ipcc.ch/report/ar6/syr/ (accessed on 14 March 2024).
- Christensen, J.H.; Hewitson, B.; Busuioc, A.; Chen, A.; Gao, X.; Held, I.; Jones, R.; Kolli, R.K.; Kwon, W.T.; Laprise, R.; et al. Chapter 11: Regional Climate Projections. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Foruth Assessment Report of the Intergovernmental Panel on climate Change Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007. [Google Scholar]
- Petit, R.; Hampe, A.; Cheddadi, R. Climate changes and tree phylogeography in the Mediterranean. Taxon 2005, 54, 877–885. [Google Scholar] [CrossRef]
- MedECC. First Mediterranean Assessment Report (MAR1). 2020. Available online: https://climate.copernicus.eu/notes-editors (accessed on 15 March 2024).
- Egerer, M.H.; Lin, B.B.; Threlfall, C.G.; Kendal, D. Temperature variability influences urban garden plant richness and gardener water use behavior, but not planting decisions. Sci. Total Environ. 2019, 646, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.; Naudiyal, N.; Wang, J.; Gaire, N.P.; Wu, Y.; Wei, Y.; He, J.; Wang, C. Assessing the impact of climate change on potential distribution of Meconopsis punicea and its influence on ecosystem services supply in the southeastern margin of Qinghai-Tibet Plateau. Front. Plant Sci. 2022, 12, 3338. [Google Scholar] [CrossRef]
- Sari, D.; Karaşah, B. Future adaptability of urban trees due to the effects of climate change: The case of Artvin, Turkey. J. Environ. Sci. Manag. 2020, 23, 60–70. [Google Scholar] [CrossRef]
- Marco, A.; Barthelemy, C.; Dutoit, T.; Bertaudière-Montes, V. Bridging human and natural sciences for a better understanding of urban floral patterns: The role of planting practices in Mediterranean gardens. Ecol. Soc. 2010, 15, 1–18. [Google Scholar] [CrossRef]
- Saurí, D. Lights and shadows of urban water demand management: The case of the metropolitan region of Barcelona. Eur. Plan. Stud. 2003, 11, 229–243. [Google Scholar] [CrossRef]
- Wade, G.L.; Midcap, J.T.; Coder, K.D.; Landry, G.W.; Tyson, A.W.; Weatherly, N., Jr. Xeriscape: A Guide to Developing a Water-wise Landscape; University of Georgia: Athens, GA, USA, 2010; Available online: https://esploro.libs.uga.edu/esploro/outputs/report/Xeriscape-a-guide-to-developing-a/9949316545202959#file-0 (accessed on 10 March 2024).
- Kattamanchi, K.; Thaneshwari, T.; Kumari, P. Xeriscapping–water efficient gardening. Int. J. All Res. Educ. Sci. Methods 2020, 8, 830–836. [Google Scholar]
- Çetin, N.; Mansuroğlu, S.; Önaç, A.K. Xeriscaping feasibility as an urban adaptation method for global warming: A case study from Turkey. Pol. J. Environ. Stud. 2018, 27, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Toscano, S.; Ferrante, A.; Romano, D. Response of Mediterranean ornamental plants to drought stress. Horticulturae 2019, 5, 6. [Google Scholar] [CrossRef]
- Juan-Vicedo, J.; Lumbreras, E.L.; Ruiz, S.R.; Casas, J. Ornamental potential of the coastal plant Lapiedra martinezii Lag. (Amaryllidaceae): The role of its revalorization in xero-gardening and ex-situ conservation. Nereis Rev. Iberoam. Interdiscip. Métodos Model. Simul. 2021, 13, 211–226. [Google Scholar] [CrossRef]
- Leotta, L.; Toscano, S.; Ferrante, A.; Romano, D.; Francini, A. New strategies to increase the abiotic stress tolerance in woody ornamental plants in Mediterranean climate. Plants 2023, 12, 2022. [Google Scholar] [CrossRef] [PubMed]
- Soltani, E.; Soltani, A. Meta-analysis of seed priming effects on seed germination, seedling emergence and crop yield: Iranian studies. Int. J. Plant Prod. 2015, 9, 413–432. [Google Scholar]
- Donohue, K.; Rubio de Casas, R.; Burghardt, L.; Kovach, K.; Willis, C.G. Postgermination, Adaptation, and Species Ecological Ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Michel, B.E.; Kaufmann, M.R. The osmotic potential of polyethylene glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef]
- Ellis, R.A.; Roberts, E.H. The quantification of aging and survival in orthodox seeds. Seed Sci. Technol. 1981, 9, 373–409. [Google Scholar]
- El-Madidi, S.A.I.D.; El-Baroudi, B.R.A.H.I.M.; Aameur, F.B. Effects of salinity on germination and early growth of barley (Hordeum vulgare L.) cultivars. Int. J. Agric. Biol. 2004, 6, 767–770. [Google Scholar]
- García-Huidobro, J.; Monteith, J.L.; Squire, G.R. Time, temperature and germination of pearl millet. J. Exp. Bot. 1982, 33, 288–296. [Google Scholar] [CrossRef]
- Trudgill, D.L. Why do tropical poikilothermic organisms tend to have higher threshold temperatures for development than temperate ones? Funct. Ecol. 1995, 9, 136–137. [Google Scholar]
- Gummerson, R.J. The effect of constant temperatures and osmotic potentials on the germination of sugar beet. J. Exp. Bot. 1986, 37, 729–741. [Google Scholar] [CrossRef]
- Kebreab, E.; Murdoch, A.J. Modelling the effects of water stress and temperature on germination rate of Orobanche aegyptiaca seeds. J. Exp. Bot. 1999, 50, 655–664. [Google Scholar] [CrossRef]
- Bradford, K.J. A water relations analysis of seed germination rates. Plant Physiol. 1990, 94, 840–849. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Rayner, G.D. Robustness to non-normality of common tests for the many-sample location problem. J. Appl. Math. Decis. Sci. 2003, 7, 187–206. [Google Scholar] [CrossRef]
- Alpert, P.T.; Ben-Gai, A.; Baharad, Y.; Benjamini, D.; Yekutieli, M.; Colacino, L.; Diodato, C.; Ramis Homar, V.; Romero, R.; Michaelides, S.; et al. The paradoxical increase of Mediterranean extreme daily rainfall in spite of decrease in total values. Geophys. Res. Lett. 2002, 29, 31-1–31-4. [Google Scholar] [CrossRef]
- Kuglitsch, F.G.; Toreti, A.; Xoplaki, E.; Della-Marta, P.M.; Zerefos, C.S.; Türkeş, M.; Luterbacher, J. Heat wave changes in the eastern Mediterranean since 1960. Geophys. Res. Lett. 2010, 37, L04802. [Google Scholar] [CrossRef]
- Paz, S.; Negev, M.; Clermont, A.; Green, M.S. Health aspects of climate change in cities with Mediterranean climate, and local adaptation plans. Int. J. Environ. Res. Public Health 2016, 13, 438. [Google Scholar] [CrossRef] [PubMed]
- Darras, A.I. Implementation of sustainable practices to ornamental plant cultivation worldwide: A critical review. Agronomy 2020, 10, 1570. [Google Scholar] [CrossRef]
- Ruíz-Pérez, M.R.; Alba-Rodríguez, M.D.; Marrero, M. The water footprint of city naturalisation. Evaluation of the water balance of city gardens. Ecol. Model. 2020, 424, 109031. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Maleki, K.; Soltani, E.; Arabhosseini, A.; Aghili Lakeh, M. A quantitative analysis of primary dormancy and dormancy changes during burial in seeds of Brassica napus. Nord. J. Bot. 2021, 39, e03281. [Google Scholar] [CrossRef]
- Mason, J.; Hopkins, A.; Hopper, J. The best environmental conditions for the germination of Celosia argentea L. CCAMLR Sci. 2019, 26, 416–423. [Google Scholar]
- Gómez, F.; Giménez, E.; Delgado, I.; de Haro, S.; del Moral, F. Estimación de los rangos de tolerancia a los factores de diversas especies mediterráneas de interés ecológico-forestal. Lazaroa 2009, 30, 145–159. [Google Scholar]
- Collins, A.; Stock, M.; Lewis, M.A.; Hansen, S.M. Celosia Cut Flower Production; USU Extension Publications, Utah State University: Logan, UT, USA, 2022. [Google Scholar]
- Okusanya, O.T. Germination and growth of Celosia cristata L., under various light and temperature regimes. Am. J. Bot. 1980, 67, 854–858. [Google Scholar] [CrossRef]
- Constantin, M.F.; Dobrin, A.; Constantin, C.G.; Toma, F. Research on the behavior of ornamental species in saline soils conditions. Sci. Pap. Ser. B Hortic. 2023, 67, 356–366. [Google Scholar]
- Moruno, F.; Soriano, P.; Vicente, O.; Boscaiu, M.; Estrelles, E. Opportunistic germination behaviour of Gypsophila (Caryophyllaceae) in two priority habitats from semi-arid Mediterranean steppes. Not. Bot. Horti Agrobot. 2011, 39, 18–23. [Google Scholar] [CrossRef]
- Estrelles, E.; Biondi, E.; Galiè, M.; Mainardi, F.; Hurtado, A.; Soriano, P. Aridity level, rainfall pattern and soil features as key factors in germination strategies in salt-affected plant communities. J. Arid. Environ. 2015, 117, 1–9. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Tribulato, A.; Patanè, C. Effects of drought stress on seed germination of ornamental sunflowers. Acta Physiol. Plant. 2017, 39, 1–12. [Google Scholar] [CrossRef]
- Gairola, S.; Hameed, A.; Rasheed, A.; AlKetbi, A.; Aljasmi, M.; El-Keblawy, A. Seed germination and salinity tolerance of habitat-indifferent halophytes as associated with geographical distribution. Seed Sci. Technol. 2022, 50, 125–140. [Google Scholar] [CrossRef]
- Maleki, K.; Soltani, E.; Seal, C.E.; Colville, L.; Pritchard, H.W.; Lamichhane, J.R. The seed germination spectrum of 486 plant species: A global meta-regression and phylogenetic pattern in relation to temperature and water potential. Agric. For. Meteorol. 2024, 346, 109865. [Google Scholar] [CrossRef]
- Vicente, M.J.; Martínez-Díaz, E.; Martínez-Sánchez, J.J.; Franco, J.A.; Bañón, S.; Conesa, E. Effect of light, temperature, and salinity and drought stresses on seed germination of Hypericum ericoides, a wild plant with ornamental potential. Sci. Hortic. 2020, 270, 109433. [Google Scholar] [CrossRef]
- Ahmad, S.; Ahmad, R.; Ashraf, M.Y.; Ashraf, M.; Waraich, E.A. Sunflower (Helianthus annuus L.) response to drought stress at germination and seedling growth stages. Pak. J. Bot. 2009, 41, 647–654. [Google Scholar]
- Alhaddad, F.A.; Abu-Dieyeh, M.H.; ElAzazi, E.S.M.; Ahmed, T.A. Salt tolerance of selected halophytes at the two initial growth stages for future management options. Sci. Rep. 2021, 11, 10194. [Google Scholar] [CrossRef]
- Khan, M.A.; Gul, B. Halophyte seed germination. In Ecophysiology of High Salinity Tolerant Plants; Springer: Dordrecht, The Netherlands, 2006; pp. 11–30. [Google Scholar]
- Zaman, S.; Padmesh, S.; Tawfiq, H. Seed germination and viability of Salsola imbricata Forssk. Int. J. Biodivers. Conserv. 2010, 2, 388–394. [Google Scholar]
- Ramírez-Tobías, H.; Peña-Valdivia, C.; Trejo, C.; Aguirre, J.; Vaquera, H. Seed germination of Agave species as influenced by substrate water potential. Biol. Res. 2014, 47, 11. [Google Scholar] [CrossRef] [PubMed]
- Kotzen, B. Plant use in desert climates-looking forward to sustainable planting in the Negev and other world deserts. Acta Hortic. 2004, 643, 39–49. [Google Scholar] [CrossRef]
- Mircea, D.M.; Calone, R.; Estrelles, E.; Soriano, P.; Sestras, R.E.; Boscaiu, M.; Sestras, A.F.; Vicente, O. Responses of different invasive and non-invasive ornamental plants to water stress during seed germination and vegetative growth. Sci. Rep. 2023, 13, 13281. [Google Scholar] [CrossRef] [PubMed]
- Süle, G.; Miholcsa, Z.; Molnár, C.; Kovács-Hostyánszki, A.; Fenesi, A.; Bauer, N.; Szigeti, V. Escape from the garden: Spreading, effects and traits of a new risky invasive ornamental plant (Gaillardia aristata Pursh). NeoBiota 2023, 83, 43–69. [Google Scholar] [CrossRef]
- Franco, J.A.; Martínez-Sánchez, J.J.; Fernández, J.A.; Bañón, S. Selection and nursery production of ornamental plants for landscaping and xerogardening in semi-arid environments. J. Hortic. Sci. Biotechnol. 2006, 81, 3–17. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).