Abstract
Watermelon (Citrullus lanatus L. Thunb. Matsum. and Nakai) is a vegetable species with high water content that is rich in lycopene and refreshing; therefore, breeding studies are carried out intensively to develop new varieties. The breeding period in these plants can be shortened with the use of the haploidization technique, and determining the ploidy levels of plants in haploidization studies is very important. In this study, it was examined whether morphological and cytological analyses would be sufficient for ploidy determination of haploid plants obtained by ovary culture in watermelon. With this aim, the stomatal, morphological and cytological characteristics were determined, taken from 15 haploid and 19 double haploid plants. The ploidy level of the plants was detected by flow cytometry before these analyses. In this study, “Principal Component Analysis” was performed based on average values to analyze the structure of the relationship between the parameters examined. It is thought that stomatal features can be used as morphological markers in determining the ploidy levels of plants. The differences obtained from the study results may help to develop effective strategies in determining the ploidy levels of plants.
1. Introduction
Watermelon (Citrullus lanatus, 2n = 2x = 22), which is widely produced around the world, is one of the ten most grown species in the Cucurbitaceae family and has high economic importance [1,2,3]. The total production amount of watermelon worldwide is 101 million tons. China, the leading country in watermelon production, produced over 60.6 million tons in 2021, and 79.5% of the world’s watermelon production is in Asia, 7.5% is in Africa, 6.9% is in America and 5.8% is in Europe [4]. The amount of watermelon production in Türkiye was 3.5 million tons in 2023 [5].
Breeding in watermelon has been carried out to produce high-yield, high-quality and disease- and pest-resistant varieties to meet the demands of both producers and consumers, and the varieties released to the market have mostly been hybrids developed with traditional breeding programs, until recently [6]. It is necessary to use new and appropriate biotechnological methods for more efficient and high-quality watermelon cultivation, especially by reducing production costs by combating possible diseases and pests. One of the most important biotechnological developments in plant breeding is the production of double haploid plants using in vitro techniques [7,8]. Producing haploid plants to obtain double haploid (DH) lines is one of the most effective breeding strategies in modern agriculture. Haploid plants are created by culturing haploid gametophytic tissues or organs such as anthers [9,10,11,12,13], microspores [14] and ovaries [8,15,16] and using the irradiated pollen technique [17,18,19,20] in in vitro conditions [21]. Haploid plant production, which is among tissue culture techniques, has an important role in plant breeding [22,23]. Haploid plants of a species have half of the normal number of chromosomes. Haploid production through in vitro techniques allows new lines to be generated more quickly and their resistance to diseases to be screened more efficiently [24]. Using haploidy techniques, 100% homozygous pure lines can be obtained in 1–2 years [6,17]. In summary, haploidization techniques provide advantages in shortening the duration of breeding and obtaining homozygous pure lines to be used in variety development in a shorter time [19]. There is limited information on obtaining haploid plants in watermelon using the ovary culture method, making these plants double haploid and determining their morphological and cytological characteristics. One of the important stages in double haploidy studies is determining the ploidy levels of plants. Methods such as phenotypic observation, chromosome counting, flow cytometry and identification of stomatal characteristics are used to determine the ploidy level. In phenotypic observation, plants are expected to develop and reach anthesis. Chromosome counting is carried out in samples obtained from plants’ healthy and strong fresh growing root tips and is a reliable method [25,26]. However, the applicability of this method is quite difficult and requires technical knowledge. The flow cytometry method is based on the analysis of the light absorbed by the cells whose characteristics will be determined while passing through the fluorescent detector one by one. The expensiveness of the flow cytometry method has led to the search for an alternative method. Since stomatal observations are the most applicable method and do not require infrastructure in determining ploidy levels, they are also widely used in breeding studies in different species such as watermelon [27], cucumber [28], squash [29,30,31] and pepper [32,33,34]. Also, thanks to a new method applied in this study, leaf samples can be stored without being damaged, and stomatal counting can be carried out without depending on time. Since watermelon leaves have a hairy structure, conventional leaf membrane peeling methods are time-consuming and difficult. This method is very practical, as it is used directly without peeling the leaves or removing the membrane. At the same time, it allows for the development and use of technology as it allows measurement with artificial intelligence.
Watermelon is a dicotyledonous and annual plant species [35]. The leaves consist of five parts (lobes) and are connected to the stem with a short or long stipe. Its leaves have a hairier structure than other cucurbits. Its stem is thin, angular, creeping and grows by spreading over the soil surface. Watermelon has small flowers that are less flashy than other cucurbits [36]. Although watermelon flowers are generally monoecious, they also have andromonoic structures and are pollinated by insects [37,38]. In watermelons, the stipes of male flowers are longer and greater in number. The pedicel of female flowers is shorter, thicker and slightly hairy [36].
Since pollination and fertilization are the main factors affecting the fruit set rate, knowing the pollen and other characteristics of species and varieties is of great importance for producers and breeders [39]. The viability and germination rates of pollen show their quality characteristics and vary depending on the genotype [40]. In all plant species, pollen characteristics (amount of pollen produced, germination rate, etc.) have a significant impact on fertilization and fruit set [39]. Climatic conditions such as humidity and temperature directly affect pollen quality [40]. The germination rate of pollen depends on environmental factors as well as the amount of nutrients such as sucrose and plant growth regulators. Following pollination, pollen germinates on the stigma [41]. Climate change affects the generative phase of plants much more than the vegetative phase. Heat stress causes morphological disorders in flower development [42], decrease in pollen viability and germination capacity [43,44], ovarian degeneration [42], decrease in embryo sac size [45], decrease in the amount of pollen [45,46], irregularities in anther development [47], decrease in pollen tube growth [44,46], abnormal embryo or endosperm development [48] and lower fruit or seed formation [49].
The aim of this study is (i) to characterize the morphological and cytological features of haploid and double haploid plants obtained by ovary culture method in different watermelon genotypes, (ii) to evaluate the relationship between these features and (iii) to determine whether the relationship between stomatal characteristics and ploidy levels in haploid and double haploid plants can be used to screen different ploidy levels based on stomatal characteristics.
2. Materials and Methods
This study was carried out in the plant tissue culture laboratories of Agricultural Sciences and Technology Faculty of Sivas Science and Technology University (Sivas, Türkiye), Department of Horticulture of Cukurova University (Adana, Türkiye) and Alata Horticultural Research Institute’s greenhouses (Mersin, Türkiye).
2.1. Materials
As plant material, haploid and double haploid plants obtained by ovary culture method from 12 watermelon genotypes [Kar 37 (1), Kar 48 (2), Kar 68 (3), Kar 180 (4), Kar 17 (5), Kar 22 (6), Kar 116 (7), Kar 192 (8), Kar 340 (9), Kar 341 (10), Kar 88 (14) and Kar 165 (15)] and 3 commercial varieties [Sturbust F1 (11), Ersin F1 (12) and Zumrut F1 (13)] were used. These genotypes were selected from the Cucurbit Genetic Resource Collection of the Cukurova University Department of Horticulture based on their morphological diversity. They were collected from different regions and provinces of Türkiye. Haploid and double haploid lines obtained by ovary culture method were planted in the greenhouses belonging to the Alata Horticultural Research Institute, and during the growing period, normal horticultural cultivation practices were implemented. The materials used in this context were selected randomly from 30 of the plants that completed their development. Genotypes Kar 37 (1), Kar 180 (4), Kar 17 (5), Kar 22 (6), Kar 192 (8), Sturbust F1 (11), Ersin F1 (12), Zumrut F1 (13) and Kar 165 (15) were used in this study. Cytological analyses were performed on two different plants of genotype number 12, but morphological observations could not be made as the plant lost its viability later on (Figure 1).

Figure 1.
Selection of the plants used in this study and their growth under greenhouse conditions (A,B).
2.2. Methods
Stomatal examinations, flow cytometry analysis and pollen analysis [50] were performed to determine the ploidy levels of the plants obtained in vitro.
2.2.1. Determination of Ploidy Level
Flow Cytometry Method
Ploidy levels of watermelon genotypes used in this study were measured by flow cytometry (Figure 2). These measurements were carried out by a private company (Ida Yaşam Teknolojileri, Istanbul, Türkiye).
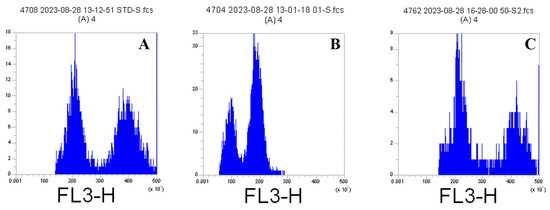
Figure 2.
Control (diploid) (A), haploid (Kar 17) (B) and double haploid (ST) (C).
Stomatal Examinations
A newly developed method by cytology laboratory of the Cukurova University Department of Horticulture based on leaf whitening after fixation and staining with safranin O during stomatal count was used. In the method used, the seventh and eighth leaves were taken from the shoot tip of all plants, and depending on the size of the leaves, whole leaves or 1 cm2 discs were taken and kept in FPA 70 fixative (1:1:18 propionic acid/formaldehyde/alcohol). During the preparation of the samples, the leaves were first washed under running water to remove the fixative from the samples. The samples were transferred to sodium hypochlorite for approximately 5 h to remove chlorophyll from the leaves and make the leaves transparent, and then they were rinsed in pure water to remove the sodium hypochlorite. Then, samples were kept in 1:1 safranin/glycerol liquid for 5 s and washed with pure water once again to remove excess dye. Imaging was performed with a DP72 camera mounted on an Olympus BX51 light (Olympus, Tokyo, Japan) microscope, and stomatal width and stomatal length measurements were carried out (Figure 3).
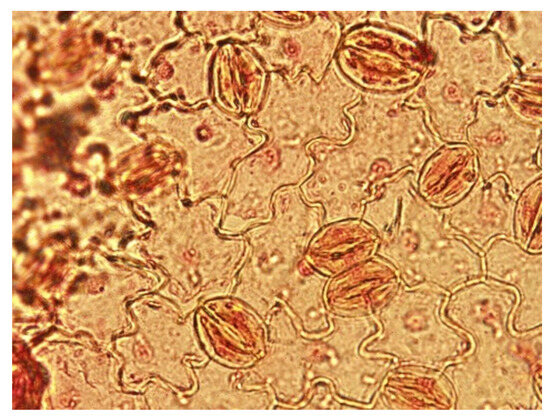
Figure 3.
Stomatal image of double haploid plant.
Stomatal density (number/mm2): In order to determine the stomatal density, a certain area was determined on the micrographs obtained, and the stomata remaining in this area were counted. The data obtained were used to determine the number of stomata per 1 mm2. Stomatal counts were made in at least 10 different areas in 3 different regions of each leaf.
Stomatal length (μm): in order to determine the stomatal length, the length of a total of 30 stomata in each leaf was measured, and the average of the obtained values was taken to determine the stomatal length of each plant.
Stomatal width (μm): in order to determine the stomatal width, the width of a total of 30 stomata on each leaf was measured, and the average of the obtained values was taken to determine the stomatal length of each plant.
Stomatal size (μm): stomatal size is obtained by multiplying the stomatal length and stomatal width [51].
Stomatal index: stomatal index was obtained by dividing the stomatal length by the stomatal width [51].
Leaf area (cm2): in the first month after planting, the 8th leaf of four plants from each genotype, starting from the growth tip [52], was scanned with a Cl-202 area meter and determined as cm2 (Figure 4).

Figure 4.
Determination of leaf areas of haploid (A) and double haploid (B) plants (cm2).
2.2.2. Determining Morphological Differences between Haploid and Double Haploid Plants
In plants determined to be haploid and double haploid, flower diameter (mm), stamen length (mm) and pedicel length (mm) were measured with a caliper 36 days after planting, when the male flowers were at anthesis stage (Figure 5) [53].

Figure 5.
Haploid (A,C) and double haploid (B,D) male flowers.
In plants determined to be haploid and double haploid, 36 days after planting, when the petal of female flower was fully opened, ovary diameter (mm), ovary height (mm), pistil height (mm) and pedicel length (mm) were measured using a caliper at anthesis stage (Figure 6) [53].

Figure 6.
Double haploid (A) and haploid (B) female flowers.
Pollen Analysis
Pollen analysis was carried out to determine the differences between the pollen of haploid and double haploid plants and to determine whether the selfing success in double haploid plants was due to pollen. In this context, in vitro pollen viability and germination rates of flowers taken from randomly selected plants from each genotype determined as haploid and diploid according to the results of flow cytometry and stomatal observations were examined. Studies on pollen include the in vitro viability and germination levels of pollen and were conducted in two different periods. The first period (Figure 7A) covers the period between December and January, when the air temperatures are on average 11 °C during the day and 10 °C at night. The second period (Figure 7B) covers the period between March and April, when the air temperatures are on average 17 °C during the day and 15 °C at night (Figure 7).

Figure 7.
Malformed male flowers observed between November and January (A,B); regular male flowers observed between March and April (C).
Determination of In Vitro Pollen Viability Rate (%)
Pollen viability rates were determined using 1% TTC test (2,3,5 triphenyl tetrazolium chloride). TTC was prepared and evaluated according to Norton [54]. To obtain fresh pollen, at least 15 flowers were isolated 1 day before anthesis from each treatment, collected at anthesis and transferred to the laboratory immediately. The pollen grains were moved to the TTC drops with a needle. For each genotype, three slide replications were prepared, and at least 100 pollen grains were counted from each replication. Pollen counting was carried out by Olympus BX51 light microscope (Olympus, Japan). While counting, red pollens were considered as “viable”, light pink as “semi viable” and uncolored pollens as “non-viable”. The pollen viability rate was calculated according to Norton [54] as the ratio of “sum of total viable and half of semi viable pollens” to “total pollen number”. Pollen viability level was determined for controlling the pollen quality.
Determination of In Vitro Pollen Germination Rate (%)
In vitro pollen germination rate was tested with the “agar in petri” method with the media consisting of 1% agar +250 ppm boric acid +10% sucrose at 25 °C [55,56]. To obtain fresh pollen, at least 15 flowers were isolated 1 day before anthesis from each treatment, collected at anthesis and transferred to the laboratory immediately. The pollen grains were moved to the Petri dishes with a needle and left there for 3 h. The pollen germination was recorded in three Petri replications by counting at least 100 pollen grains in each Petri dish. Pollen counting was performed by Olympus BX51 light microscope (Olympus, Japan). Pollen was recorded as germinated when the length of the pollen tube exceeded its own diameter.
2.2.3. Inbreeding Success (%)
In order to determine the success of inbreeding, inbreeding was performed on each plant, and for this aim, male and female flowers on the same plant were closed with clips one day before the anthesis stage. One day later, pollination was carried out by rubbing the pollen on the stigma (Figure 8). Female flowers that did not set fruit at the period from the pollination stage to fruit set were counted, and this process continued until fruit set was obtained. Inbreeding success (%) was determined by ratio of total number of pollinated female flowers to the number of fruit set.

Figure 8.
Inbreeding processes in double haploid plants ((A) female flower closed one day before anthesis; female flower pollinated on the day of (B,C) anthesis).
2.2.4. Statistical Analysis
This study was conducted according to the randomized plot experimental design with three replications and five samples in each replication. The data were analyzed in the JMP version 13.0.1 (SAS Institute Inc., Cary, NC, USA) statistical program and the LSD test was performed on the data when the difference was statistically significant. ANOVA was carried out to determine the differences of cultivars, harvest dates and their interaction regarding all examined variables. The least significant difference (LSD) test was used to determine the difference among the groups. In addition, the principle component analysis (PCA) was performed by using XL Stat trial software 2023.5.1 (Addinsoft, New York, NY, USA).
3. Results
3.1. Determination of Ploidy Levels of Plants
3.1.1. Flow Cytometry
The results of the flow cytometry measurement are shown in Table 1 and Figure 9. The leaf sample used as a control was taken from a commercial variety known to be diploid. According to the measurement results, 15 out of 34 plants were determined to be haploid, while 19 plants were determined to be double haploid. The following are the number of plants determined as double haploid: 3/5 plants in genotype number 1, 2/5 plants in genotype number 4, 3/5 plants in genotype number 5, 1/4 plants in genotype number 6, 1/4 plants in genotype number 8, 2/3 plants in genotype number 11, 3/3 plants in genotype number 13 and 4/5 plants in genotype number 15.

Table 1.
Stomatal results in haploid and double haploid plants of watermelon genotypes.
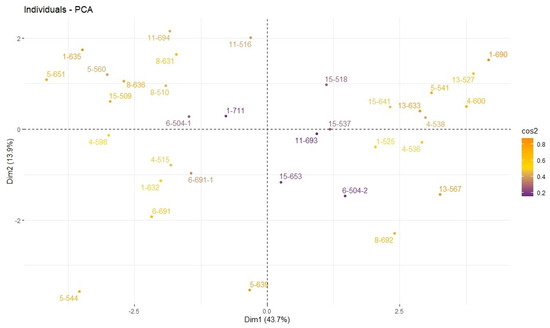
Figure 9.
Closeness distance relationships (PCA) of plants in terms of examined traits.
3.1.2. Stomatal Examinations
Some microscopic observations were carried out on plants where the ploidy level was determined by flow cytometry. The results obtained regarding the stomata of haploid and double haploid plants are presented in Table 1. Within the scope of this study, the stomatal length (μm), stomatal width (μm), stomatal size (μm), stomatal index (μm), stomatal density (μm) and leaf area (cm2) of 34 plants were examined. According to the study results, the differences between all examined parameters were found to be statistically significant.
When the stomatal lengths of plants were compared, the highest values were obtained from the genotypes/plants numbered 4-538 (14.57 μm), 13-567 (13.98 μm) and 6-504-2 (12.82 μm), while the lowest values were found in the genotypes/plants numbered 1-635 (9.32 μm) and 5-560 (9.07 μm). In the stomatal width, it was observed that the stomatal width of diploid plants was larger than haploid plants. The highest stomatal width values were determined in the genotypes/plants numbered 15-641 (9.94 μm), 8-692 (9.89 μm), 4-600 (9.80 μm), 13-527 (9.77 μm), 4-536 (9.38 μm), 5-541 (9.37 μm), 13-567 (9.28 μm), 13-633 (9.27 μm) and 15-537 (9.16 μm). According to the flow cytometry results, these plants were determined to be double haploid. The lowest stomatal widths were obtained from haploid plants and were determined in the genotypes/plants numbered 1-635 (7.29 μm) and 5-544 (7.27 μm).
When the stomatal sizes of the plants were compared, the highest values were obtained from double haploid plants such as the genotypes/plants numbered 8-692 (199.32 μm), 15-641 (129.90 μm) and 13-567 (129.77 μm) (Table 1). The lowest values were determined in haploid plants such as the genotypes/plants numbered 5-544 (71.27 μm) and 1-635 (67-87 μm). When the stomatal indexes of plants were examined, the highest results were found in the genotypes/plants numbered 6-504-2 (1.79 μm) and 4-538 (1.68 μm). The lowest values were determined in the genotypes/plants numbered 15-518 (1.24 μm) and 5-560 (1.14 μm). It has been observed that the stomatal indexes of haploid plants are higher than those of double haploid plants.
In terms of stomatal density, it was determined that the densities of the genotypes/plants numbered 8-692 (40.00 mm2), 15-653 (36.33 mm2) and 5-639 (32.67 mm2) were higher (Table 1). It was found that the genotypes/plants numbered 5-544 (5.00 mm2) and 5.651 (4.67 mm2) had the least stomatal density.
When plants were compared in terms of leaf area, the largest leaves were obtained from the genotypes/plants numbered 5-544 (134.70 cm2) and 5-639 (132.74 cm2) (Table 1). The smallest leaves were determined in the genotypes/plants numbered 6-691-1 (33.22 cm2) and 11-516 (26.51 cm2). It was concluded that the ploidy levels of the plants could not be determined based on the leaf area results.
3.2. Determining Morphological Differences between Haploid and Double Haploid Plants
The data obtained as a result of flow cytometry analysis and stomatal examinations supported each other. In this context, the cytological [pollen germination rate (%) and pollen viability rate (%)] and morphological characteristics [male flower diameter (mm), stamen length (mm), male flowers pedicel length (mm), ovary length (mm), ovary diameter (mm), pistil height (mm) and female flower pedicel length (mm)] of flowers taken from randomly selected haploid and double haploid plants were determined.
3.2.1. Flower Characteristics of Haploid and Double Haploid Plants
The measurements carried out on the flowers of the plants randomly selected among the genotypes determined to be haploid and double haploid are shown in Table 2. In terms of male flower diameter, the differences between genotypes were found to be statistically significant. The genotype/plant having the highest flower diameter was 5-560 (39.71 mm), and it was followed by the genotypes/plants numbered 13-633 (37.77 mm) and 13-527 (37.27 mm). The genotype/plant having the lowest male flower diameter was 11-694 (11.33 mm). It has been determined that the effect of genotypes on the stamen length of plants is important. Accordingly, the longest stamen length was found in the genotype/plant numbered 13-527 (10.89 mm). The shortest stamen lengths were obtained in the genotypes/plants numbered 5-651 (5.19 mm), 6-691 (4.08 mm) and 5-544 (3.80 mm).
Table 2.
Female and male flower measurements in haploid and double haploid watermelon plants.
It was determined that the effect of genotypes on male flower stipe length was statistically significant. The longest male flower stipe length was obtained from the genotype/plant numbered 13-527 (29.36 mm), and it was followed by genotypes/plants numbered 15-641 (27.04 mm) and 15-518 (26.42 mm). The ploidy levels of these plants were double haploid. The shortest male flower stipe lengths were detected in the genotypes/plants numbered 6-504-2 (11.21 mm), 5-544 (11.04 mm) and 5-639 (9.90 mm). These plants have a haploid structure and are statistically in the same group.
When the ovary height of the genotypes was examined, the differences between the averages were found to be statistically significant (Table 2). The highest ovary heights were determined in the genotypes/plants numbered 4-536 (17.97 mm), 5-541 (17.51 mm), 4-538 (17.35 mm) and 11-694 (16.37 mm). The lowest ovary heights were obtained from the genotypes/plants numbered 5-560 (5.96 mm), 5-651 (5.55 mm) and 5-544 (4.74 mm).
In terms of ovary diameter, the highest values were obtained from the genotypes/plants numbered 4-600 (12.18 mm) and 1-690 (11.77 mm), and the lowest values were found in the genotypes/plants numbered 8-510 (3.35 mm) and 5-544 (3.22 mm) (Table 2). It was determined that the ovary diameters of haploid flowers were smaller than double haploid ones.
It has been determined that the effect of genotypes on the pistil height of flowers is important. Accordingly, the highest pistil height was determined in the genotype/plant numbered 5-541 with 7.98 mm, and it was followed by the genotypes/plants numbered 4-600 (7.82 mm) and 4-538 (7.58 mm).
The differences between the female flower stipe lengths of the plants used in this study were found to be statistically significant (Table 2). In this context, the longest female flower stipe length was observed in the genotype/plant numbered 13-567 (39.57 mm). The shortest female flower stipe lengths were determined in the genotypes/plants numbered 6-691 (9.12 mm) and 4-598 (7.24 mm). It was concluded that the male and female flower structures of haploid plants are smaller than those of double haploid plants.
3.2.2. Pollen Analysis
Studies on pollen include the in vitro viability and germination levels of pollen and were conducted in two different periods. The first period (A) covers the period between December and January, when the air temperatures are on average 11 °C during the day and 10 °C at night. The second period (B) covers the period between March and April, when the air temperatures are on average 17 °C during the day and 15 °C at night. Pollen germination and viability rates of plants are shown in Table 3. Genotypes and period–genotype interaction had a statistically significant effect on pollen germination rate. It was determined that the pollen germination rate was the highest in the second period (39.17%), followed by the first period (10.03%). When the effect of genotypes on pollen germination was examined, the highest germination rate was observed in the genotypes/plants numbered 5-639 (52.86%), 15-653 (52.54%) and 13-633 (50.88%), and these plants were statistically in the same group. The lowest germination rates were determined in the genotypes/plants numbered 6-504-1 (6.52%) and 5-560 (3.84%). When the interaction of periods and genotypes was examined, it was found that the highest germination rate was obtained from the second period analysis of the genotype/plant numbered 13-527 (87.72%). It was determined that the lowest interaction was in the first period analysis of the genotype/plant numbered 15-641 (0.33%). It was observed that the pollen of haploid plants did not germinate.
Table 3.
Pollen viability and germination rates (%) in haploid (H) and double haploid (DH) plants of watermelon genotypes.
It was determined that the effects of periods, genotypes and genotype–period interaction on pollen viability rate were important. It was noted that while the pollen viability rate was 17.27% in the first period, it increased and reached 57.93% in the second period. When the effect of genotypes on pollen viability rate was examined, the highest values were obtained from the genotypes/plants numbered 15.518 (81.86%), 11-693 (81.30), 15-641 (80.97%) and 13-567 (79.34%), and these plants were statistically in the same group. The pollen viability rate of the genotype/plant numbered 15-509 (5.00%) was found to be the lowest. In the period–genotype interaction, the highest values were obtained from the flowers taken in the second period of the genotypes/plants numbered 13-567 (96.49%), 15-537 (96.02%) and 13-527 (95.78%).
When pollen viability and germination rates in double haploid plants were examined, it was observed that pollen has higher viability and germination levels in the second period, when the air temperature is on average 17 °C. This is because air temperatures have significant effects on the vigor and germination rates of pollen. The movement of pollen collected during the period when the air temperature is low was noted, and it was observed that the pollen sticks to each other. Due to low air temperatures, it was determined that there were deformations in the anthers (Figure 5) and low pollen viability and germination rates. Temperature directly affects the development of pollen in the anther, the process of reaching the stigma and the germination time [57].
Although there were haploid plants in which pollen was not found at all, the presence of pollen at low rates was detected in plants of some genotypes. When these pollens were tested for viability and germination, it was found that the rates were very low.
3.3. Inbreeding Success
Inbreeding was carried out in both periods in randomly selected genotypes with plants determined to be double haploid (Table 4). In the first period, success was not achieved in the inbreeding process. This can be explained by the fact that low air temperatures negatively affect pollen viability and germination rates, and the pollen cannot germinate on the stigma. It is clearly seen that the increasing air temperatures in the second period have a positive effect on selfing rates.
Table 4.
Inbreeding success in plants (%).
To analyze the structure of the relationship among a group of 34 watermelon genotypes/plants obtained by ovary culture, “Principal Component Analysis (PCA)” was performed based on mean values. Using Principal Component Analysis (PCA), it was determined that there are two main components (Dim1 and Dim2) that represent the variation in watermelon genotypes (Figure 10). The arrows in the graph represent the projections of the variables on the principal components and describe how each variable contributes to the PCA analysis. Dim1 (X-Axis) represents the largest variation in the data and explains 43.7% of the total variation, and Dim2 (Y-Axis) represents the second largest variation and explains 13.9% of the total variation. Male flower stipe length, female flower stipe length, stamen size, pistil height, ovary height, male flower diameter, ovary diameter, stomatal width, stomatal size and stomatal length variables have high effects on Dim1. On Dim2, it has been seen that stomatal density, stomatal index and leaf area variables have more significant effects.
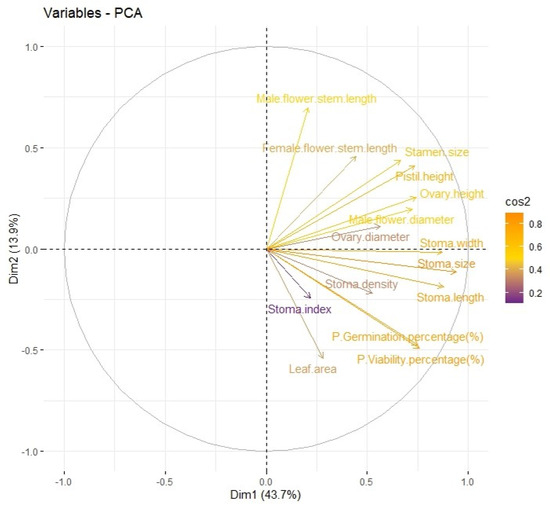
Figure 10.
Distribution of variables on two main components (Dim1 and Dim2) obtained using Principal Component Analysis (PCA).
Figure 9 shows variations in genotypes/plants in the dataset and the distribution of these variations on the main components. The color scale represents the Cos2 values of individuals and contributes to understanding the similarities and differences between individuals. Accordingly, it was understood that the genotypes/plants numbered 1-635, 1-690 and 13-633 had high Cos2 values, and it was noted that these genotypes represented the main variation well. It was determined that the genotypes/plants numbered 1-711, 6-504 and 11-693 had low Cos2 values and contributed less to the main variation. Some genotypes appear to be clearly different from the remaining genotypes in terms of the above-mentioned parameters, indicating that these parameters are the main source of variations.
Figure 11 shows the loadings and contributions of each variable in the PCA components (Dim1–Dim15). Especially in Dim1 and Dim2, certain variables have high loadings, and these variables represent the main sources of variation in the dataset. The Dim1 component was strongly represented by stomatal features such as stomatal length, stomatal width and stomatal size. Additionally, variables such as pollen germination and pollen viability percentages were also important in this component. The stomatal index has a significant effect on the Dim2 component and plays an important role in this component. In other components (Dim3–Dim15), certain variables have lower loadings and generally lower Cos2 values. These components were less representative of variation.
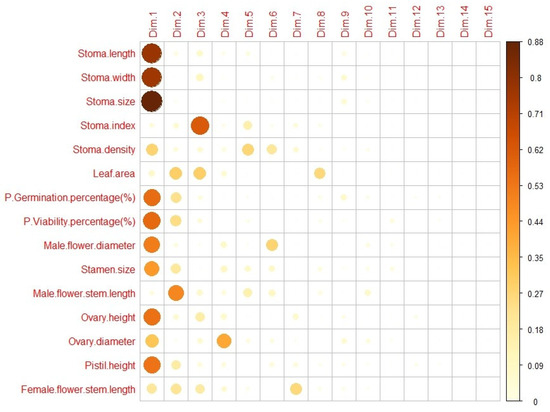
Figure 11.
A combination of “heatmap” and “bubble plot” showing Principal Component Analysis (PCA) results.
Figure 12 shows the relationships between plant morphological and stomatal characteristics in detail. Significant positive correlations were observed between stomatal size and flower structures. While there is a strong positive correlation between stomatal length and stomatal characteristics, there is a weak (−0.18) negative correlation between leaf area. While there is a weak positive correlation between stomatal density and flower structures, there is a strong negative correlation between stomatal length and stomatal size. There are strong positive correlations between pistil height and variables such as stamen size (0.58), ovary height (0.85), ovary diameter (0.55), stomatal length (0.55), stomatal size (0.59) and stomatal width (0.55), which indicate that these features show a tendency to increase together.
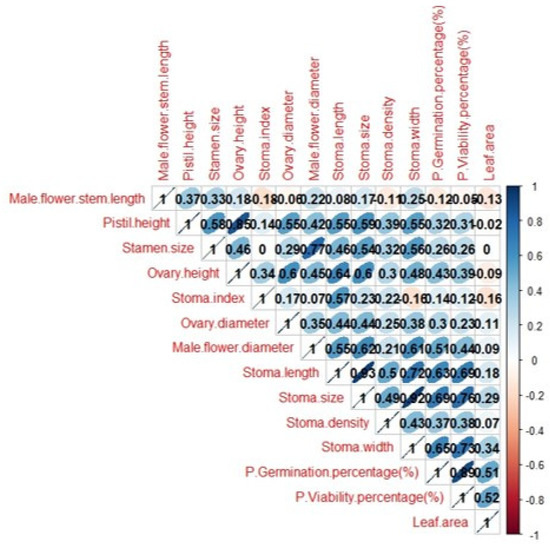
Figure 12.
Correlation matrix.
4. Discussion
Since haploid plants contain only one set of alleles at each locus, they allow doubling of the number of chromosomes to obtain 100% homozygous pure lines in one generation. In order for haploid plants to be used in breeding programs, their chromosome numbers must be doubled to obtain a double haploid [13,58,59,60]. In this context, studies on determining the ploidy level of the obtained plants economically, quickly and without requiring too much infrastructure are important.
During this study, all stomatal characteristics in haploid and double haploid plants were found to be statistically significant. When the study results were evaluated, it was observed that there were increases in stomatal width, stomatal size and stomatal density when moving from haploid to double haploid. A very strong negative relationship was found between stomatal density and stomatal size. As a result of this study, it was once again revealed that stomatal characteristics are an important factor in ploidy analysis. It is thought that stomatal features can be used as morphological markers to screen for different ploidy levels. Determination of stomatal characteristics is both easy and does not require expensive equipment. In addition, the new method used for stomatal examinations in this study is more practical than the conventional leaf membrane peeling methods for plant species with hairy and indented leaf characteristics such as watermelon and is ideal for ensuring that a large number of samples are fixed and stored and then examined under the same conditions. Since leaves are used directly in the new method, it allows stomatal examinations to be easily carried out in plants with dense veins such as beans, where it is difficult to remove membranes [61]. This method also allows for the counting of hairs and chloroplasts on leaves. In addition, the method used allows measurement with artificial intelligence and is open to new generation investigations.
In one study, more significant differences were found between haploid and spontaneous double haploid plants in terms of the number of stomata and stomatal length than in stomatal width, and it was reported that these were more reliable features than stomatal length in confirming the ploidy level [60]. As the ploidy level increases in plants, the length and size of stomatal guard cells also increase, resulting in decreases in stomatal density [51,62]. According to the results of the observations carried out on the stomata of watermelon, it has been reported that the higher the ploidy level of the chromosome, the larger the stomatal length [63]. Stomatal size (length and width) is an indicator of ploidy levels as a measure of stomatal size and has been used to determine ploidy levels in different plant species [64]. In a study conducted by Sari et al. [27], it was determined that the stomatal length of double haploid watermelon plants was 24.0 mm, while haploids were approximately 17.5 mm. It was observed that the average stomatal diameter was 18.2 mm in double haploid plants and 11.3 mm in haploid plants. The average leaf area of haploid plants was found to be 9.0 cm2, while in double haploids it was 44.3 cm2. Khandaker et al. [65] reported that the leaf area of a watermelon is 8.70 cm2 under normal conditions. Jibril et al. [66] found that the stomatal length in a standard watermelon is 3.27 μm, stomatal width is 2.16 μm, the number of stomata is 22.00 and the stomatal index is 30.32. Demirel and Onus [28] reported that the chloroplasts of haploid plants are between 4 and 6 and double haploid plants are between 8 and 12 in cucumber. In this study, it was examined whether there are sufficient criteria in determining the ploidy level of stomatal features. It has been shown that ploidy levels can be determined by stomatal measurements in watermelon, the same as in cucumber [28] and melon [67].
In order to determine the most suitable pollination period and pollination hours for plants, pollen is collected at different maturation stages and physiological periods of plants and flowers and subjected to viability and germination tests. In this way, attempts have been made to overcome the fertilization problems that may occur by determining whether the cause is due to different physiological conditions or pollen [68]. Pollen germination rate is directly related to environmental temperature. As a result of different studies, it has been reported that the pollen germination media being around 25–30 °C positively affects germination [69,70]. Factors affecting pollination success in watermelon include the viability of pollen and equal distribution of pollen on the stigma [71]. Watermelon yield is negatively affected due to inadequate pollination and lack of pollen transfer [72]. Sravani et al. [73] reported that the percentage of fruit set in watermelon was affected by different interspecific pollens. They stated that the fruit set rate in plants pollinated with bottle gourd pollen was 49.26%. Adigüzel et al. [56] found that the germination rates of tetraploid and diploid watermelon genotypes varied between 95.53% (double haploid) and 87.60% (tetraploid), and pollen viability rates varied between 68.75% and 79.50%. Sensoy et al. [68] conducted pollen studies on melon, watermelon, squash and cucumber and reported that the pollen germination rate in watermelon was 90.8% and the viability rate was 78.5%. When the pollen studies carried out in this study were examined, it was determined that pollen viability and germination capacities were low at low temperatures and germination and viability increased with increasing air temperatures. It has been observed that the success of selfing increases as a result of the increase in pollen viability and germination rates. Pollen viability of haploid plants also increased with air temperatures; however, successful selfings were not achieved because the germination power was not sufficient.
In the measurements carried out to determine the flower characteristics of haploid and double haploid plants, it was determined that the flowers of haploid plants were smaller. There is a positive relationship between flower sizes. While there is a moderate positive correlation (0.55) between pistil height and stamen size, there is a strong positive correlation (0.59) with ovary height. In one study, it was reported that the diameter of the male flower was 28.67 mm, the diameter and length of the ovary were between 10.01 and 11.12 mm and the length of the ovary was between 13.65 and 15.18 mm in double haploid plants. In the same study, it was stated that tetraploid plants are larger than diploid plants in terms of leaf and flower structures and diploid–tetraploid plants are morphologically different [74].
5. Conclusions
The information obtained from this study clearly demonstrates that ploidy levels can be determined based on cytological data such as the pollen germination rate (%) and pollen viability rate (%) of flowers and morphological characteristics such as male flower diameter (mm), stamen length (mm), male flower pedicel length (mm), ovary height (mm), ovary diameter (mm), pistil height (mm) and female flower pedicel length (mm). These findings may help develop more effective strategies for identifying haploid and double haploid plants. Preliminary selection of haploid and double haploid watermelon plants can be carried out by evaluating leaf and flower characteristics. Plants determined to be haploid or double haploid by pre-selection should be re-selected by checking stomatal size (width), stomatal index and stomatal density. In double haploid plants, after male and female flowers are formed, selfing can be carried out to ensure fruit set, depending on pollen viability and germination ability. Since fruits cannot be obtained from haploid plants, if chromosome doubling is not performed, the plants are lost. However, when considered in terms of time, it should not be forgotten that waiting for growing and anthesis periods for the plants is quite time-consuming. Precise determination of the ploidy levels of plants can be carried out by chromosome counting and flow cytometric analysis. However, in cases where there is insufficient laboratory infrastructure, selections can be carried out by morphological and cytological evaluations, and fruit sets can be more easy, applicable and economical. It is thought that this study will provide a different perspective for characterizing haploid and double haploid genotypes easily in future plant breeding studies.
Author Contributions
E.K.: experiments, data analysis and writing—original draft; H.T.: supervision, conceptualization and writing—review and editing; Ş.K.: experiments and review and editing; İ.S.: material supply—review and editing; N.S.: material supply—review and editing; T.K.: data analysis—review and editing; G.B.: experiments, supervision, conceptualization and writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
The authors would like to thank Sivas Science and Technology University (Sivas, Türkiye) for their financial support of this project under the project code 2024-ÖAP-TBTF-0001.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Acknowledgments
The authors would like to thank Sivas Science and Technology University (Sivas, Türkiye) for financial support, Cukurova University (Adana, Türkiye) and Alata Horticultural Research Institute (Mersin, Türkiye) that supported this study with laboratory and greenhouse facilities and Berken ÇİMEN (Cukurova University, Adana, Türkiye) for the “Principal Component Analysis (PCA)”.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Dou, J.; Kang, Q.; Li, T.; Umer, M.J.; Alharthi, B.; Liu, D.; Yang, S.; Niu, H.; Ma, C.; Zhu, H.; et al. Construction and application of a new watermelon germplasm with the phenotype of dwarf and branchless. Funct. Integr. Genom. 2023, 23, 310. [Google Scholar] [CrossRef] [PubMed]
- Jordana, C.N.; Stapleton, S.C.; Colee, J.C.; Lee, S.; Gao, Z.; Ray, Z.T.; Anrecio, L.R.; Freed, D.J.; Zhao, X. How Does Watermelon Grafting Impact Fruit Yield and Quality? A Systematic Review. HortScience 2023, 58, 836–845. [Google Scholar] [CrossRef]
- Tian, S.; Zhang, J.; Zhao, H.; Zong, M.; Li, M.; Gong, G.; Wang, J.; Zhang, J.; Ren, Y.; Zhang, H.; et al. Production of double haploid watermelon via maternal haploid induction. Plant Biotechnol. J. 2023, 21, 1308–1310. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Food and Agriculture Organization of the United Nation. Available online: http://www.fao.org (accessed on 31 March 2024).
- TÜİK. Türkiye İstatistik Kurumu (TÜİK). Available online: https://www.tuik.gov.tr/ (accessed on 31 March 2024).
- Sarı, N.; Solmaz, İ. Doubled Haploid Production in Watermelon. Methods Mol. Biol. 2021, 2289, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.C.; Coleman, J.D.; Kearns, A. Plant Cell Culture; Bios Scientific Publications: London, UK, 2003; p. 192. [Google Scholar]
- Baktemur, G.; Keleş, D.; Kara, E.; Taşkın, H. Effects of genotype and nutrient medium on obtaining haploid plants through ovary culture in cucumber. Mol. Biol. Rep. 2022, 49, 5451–5458. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Yiqin, G.; Bo, D.; Tonghui, R. Induction of watermelon anther callus. Hubei Agric. Sci. 2005, 5, 93–95. [Google Scholar]
- Zhu, Y.; Sun, Z.; Sun, D.; Deng, Y.; Wang, Z.; Liu, J. Advances of watermelon anther culture technology. China Cucurbits Veg. 2010, 23, 28–31. [Google Scholar]
- Taskin, H.; Büyükalaca, S.; Keleş, D.; Ekbiç, E. Induction of microspore-derived embryos by anther culture in selected pepper genotypes. Afr. J. Biotechnol. 2011, 10, 17116–17121. [Google Scholar]
- Zhu, Y.; Sun, D.; Deng, Y.; An, G.; Li, W.; Liu, J. Effects of preliminary treatment on watermelon anther culture callus induction. China Cucurbits Veg. 2012, 25, 17–19. [Google Scholar]
- Keleş, D.; Pınar, H.; Ata, A.; Taşkın, H.; Yıldız, S.; Büyükalaca, S. Effect of pepper types on obtaining spontaneous doubled haploid plants via anther culture. HortScience 2015, 50, 1671–1676. [Google Scholar] [CrossRef]
- Ata, A.; Keleş, D.; Taşkın, H.; Büyükalaca, S. Effects of season, genotype, and nutrient medium on pepper anther culture and microspore development. Turk. J. Agric. For. 2019, 43, 123–137. [Google Scholar] [CrossRef]
- Zou, T.; Su, H.N.; Wu, Q.; Sun, X.W. Haploid induction via unfertilized ovary culture in watermelon. Plant Cell Tissue Organ Cult. 2018, 135, 179–187. [Google Scholar] [CrossRef]
- Zhu, Y.C.; Sun, D.X.; Deng, Y.; An, G.L.; Li, W.H.; Si, W.J.; Liu, J.P.; Sun, X.W. Comparative transcriptome analysis of the effect of different heat shock periods on the unfertilized ovule in watermelon (Citrullus lanatus L.). J. Integr. Agr. 2020, 19, 528–540. [Google Scholar] [CrossRef]
- Baktemur, G.; Taşkın, H.; Büyükalaca, S. Comparison of different methods for separation of haploid embryo induced through irradiated pollen and their economic analysis in melon (Cucumis melo var. inodorus). Sci. World J. 2013, 2013, 529502. [Google Scholar] [CrossRef] [PubMed]
- Taşkın, H.; Yücel, K.N.; Baktemur, G.; Çömlekçioğlu, S.; Büyükalaca, S. Effects of different genotypes and gamma ray doses on haploidization with irradiated pollen technique in watermelon (Citrullus lanatus L.). Can. J. Plant Sci. 2013, 93, 1165–1168. [Google Scholar] [CrossRef]
- Baktemur, G.; Yücel, N.K.; Taşkın, H.; Çömlekçioğlu, S.; Büyükalaca, S. Effects of different genotypes and gamma ray doses on haploidization using irradiated pollen technique in squash. Turk. J. Biol. 2014, 38, 318–327. [Google Scholar] [CrossRef]
- Yıldız, Ç.; Koruk, M.; Doğan, A.; Ellialtıoğlu, Ş.Ş. Parthenogenetic Embryogenesis Frequencies of Different Cucumber Genotypes by Irradiated Pollen Pollination and Haploid Embryo Development. Turk. J. Nat. Sci. 2020, 9, 69–73. [Google Scholar] [CrossRef]
- Chen, X.; Li, Y.; Ai, G.; Chen, J.; Guo, D.; Zhu, Z.; Zhu, X.; Tian, S.; Wang, J.; Liu, M.; et al. Creation of a watermelon haploid inducer line via ClDMP3-mediated single fertilization of the central cell. Hortic. Res. 2023, 10, 6. [Google Scholar] [CrossRef]
- Heiser, C.B.J.R. Peppers. In Evoluation of Crop Plants; Simmands, N.W., Ed.; Longman Science & Technology Report: London, UK, 1976; pp. 265–268. [Google Scholar]
- Andrews, J. Peppers. The Domasticated Capsicum; University of Texas Press: Austin, TX, USA, 1985. [Google Scholar]
- Dong, Y.Q.; Zhao, W.X.; Li, X.H.; Liu, X.C.; Gao, N.N.; Huang, J.H.; Wang, W.Y.; Xu, X.L.; Tang, Z.H. Androgenesis, Gynogenesis, and Parthenogenesis Haploids in Cucurbit Species. Plant Cell Rep. 2016, 35, 1991–2019. [Google Scholar] [CrossRef]
- Keller, J. Culture of Unpollinated Ovules, Ovaries and Flower Buds in Some Species of The Genus Allium and Haploid Induction Via Gynogenesis in Onion (Allium cepa L.). Euphytica 1990, 47, 241–247. [Google Scholar] [CrossRef]
- Yaralı, F.; Yanmaz, R. Utilization of Haploidy Techniques in Breeding of Allium Species. Türk Bilimsel Derlemeler Derg. 2013, 6, 44–50. Available online: https://dergipark.org.tr/en/download/article-file/417890 (accessed on 31 March 2024). (In Turkish).
- Sari, N.; Abak, K.; Pitrat, M. Comparison of ploidy level screening methods in watermelon: Citrullus lanatus (Thunb.) Matsum. and Nakai. Sci. Hortic. 1999, 82, 265–277. [Google Scholar] [CrossRef]
- Demirel, E.; Onus, A.N. Obtaining Haploid Embryo and Plant by Gynogenesis in Some Cucumber (Cucumis sativus L.) Cultivars and Types. Uluslararası Tarım Yaban Hayatı Bilim. Derg. 2021, 7, 360–367. (In Turkish) [Google Scholar] [CrossRef]
- Kurtar, E.S.; Uzun, S.; Esendal, E. Haploid Plant Propagation by Anther Culture of Squash (Cucurbita pepo L.). Ondokuz Mayıs Üniversitesi Ziraat Fakültesi Derg. 1998, 14, 33–45. (In Turkish) [Google Scholar]
- Kurtar, E.S.; Balkaya, A.; Ozbakir, M.; Ofluoglu, T. Induction of haploid embryo and plant regeneration via irradiated pollen technique in pumpkin (Cucurbita moschata Duchesne ex. Poir). Afr. J. Biotechnol. 2009, 8, 5944–5951. [Google Scholar]
- Kurtar, E.S.; Balkaya, A.; Göçmen, M.; Karaağaç, O. Dihaploidization in Squash Genotypes (Cucurbita spp) as a Rootstock Candidates for Cucumber (Cucumis sativus L.) via Irradiated Pollen Technique. Selcuk J. Agric. Food Sci. 2017, 31, 34–41. (In Turkish) [Google Scholar] [CrossRef]
- Supena, E.D.J.; Muswita, W.; Suharsono, S.; Custers, J.B.M. Evaluation of crucial factors for implementing shed-microspore culture of Indonesian hot pepper (Capsicum annuum L.). Sci. Hortic. 2006, 107, 226–232. [Google Scholar] [CrossRef]
- İlhan, M.; Kurtar, E.S. Doublehaploidization Efficiency of Selected Pepper Genotypes Via In Vitro Anther Culture. Selcuk. J. Agric. Food Sci. 2022, 36, 253–259. [Google Scholar] [CrossRef]
- Zahroh, Z.A. Dihaploidization in Some Indonesian Pepper Genotypes via Anther Culture. Master’s Thesis, Sakarya University, Sakarya, Türkiye, 2022. (In Turkish). [Google Scholar]
- Güner, N.; Wehner, T.C. The genes of watermelon. HortScience 2004, 39, 1175–1182. [Google Scholar] [CrossRef]
- Wehner, T.C. Watermelon. In Vegetables I; Handbook of Plant Breeding; Prohens, J., Nuez, F., Eds.; Springer: New York, NY, USA, 2008; Volume 1. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Mayer, D.F. Crop Pollination by Bees; CABI Publishing: Wallingford, UK, 2005. [Google Scholar]
- Wijesinghe, S.A.E.C.; Evans, L.J.; Kirkland, L.; Rader, R. A global review of watermelon pollination biology and ecology: The increasing importance of seedless cultivars. Sci. Hortic. 2020, 271, 109493. [Google Scholar] [CrossRef]
- Abacı, Z.T.; Asma, B.M. Pollen Vitality, Germination Conditions and Pollen Tube Lenght Investigation of Hybrid Apricot Genotypes. Anadolu J. Agric. Sci. 2014, 29, 12–19. (In Turkish) [Google Scholar] [CrossRef]
- Kılıç, T.; Doğan, E.; Dursun, H.B.; Çamurcu, S.; Ünsal, H.T.; Kazaz, S. Effects of Pollen Holding Duration in Some Rose Species and Varieties on Pollen Viability and Germination. J. Agric. Fac. Bursa Uludag Univ. 2020, 34, 173–184. (In Turkish) [Google Scholar]
- Çetin, M.; Soylu, A. Standart Ayva Çeşitlerinin Döllenme Biyolojisi Üzerinde Araştırmalar. Bahçe 2006, 35, 83–96. (In Turkish) [Google Scholar]
- Albertos, P.; Wagner, K.; Poppenberger, B. Cold stress signalling in female reproductive tissues. Plant Cell Environ. 2019, 42, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Hedhly, A.; Hormaza, J.I.; Herrero, M. The effect of temperature on pollen germination, pollen tube growth, and stigmatic receptivity in peach. Plant Biol. 2005, 7, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Karabıyık, Ş. Effects of temperature on pollen viability and in vivo pollen tube growth in Citrus sinensis. J. Appl. Bot. Food Qual. 2022, 95, 100–104. [Google Scholar] [CrossRef]
- Bykova, O.; Chuine, I.; Morin, X.; Higgins, S.I. Temperature dependence of the reproduction niche and its relevance for plant species distributions. J. Biogeogr. 2012, 39, 2191–2200. [Google Scholar] [CrossRef]
- Snider, J.L.; Oosterhuis, D.M.; Kawakami, E.M. Diurnal pollen tube growth is slowed by high temperature in field-grown Gossypium hirsutum pistils. J. Plant Physiol. 2011, 168, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Guo, S.; Wang, D.; Zhong, Y.; Chen, M.; Chen, C.; Cheng, D.; Liu, Z.; An, T.; Li, J.; et al. ZmCOI2a and ZmCOI2b redundantly regulate anther dehiscence and gametophytic male fertility in maize. Plant J. 2022, 110, 849–862. [Google Scholar] [CrossRef]
- Cuevas, J.; Rallo, L.; Rapoport, H.F. Procedure to study ovule sensescence inolive. Acta Hortic. 1994, 356, 252–255. [Google Scholar] [CrossRef]
- Carpenedo, S.; Raseira, M.C.B.; Byrne, D.H.; Franzon, R.C. The effect of heat stress on the reproductive structures of peach. J. Am. Pomol. Soc. 2017, 71, 114–120. [Google Scholar]
- Karabıyık, Ş.; Eti, S. Determination of Flowering Dates and Pollen Properties of Some Pecan Nut Cultivars in Adana Ecological Conditions. Turk. J. Agric. Food Sci. Technol. 2018, 6, 1795–1801. (In Turkish) [Google Scholar] [CrossRef][Green Version]
- Cimen, B. Induction of Polyploidy in C35 Citrange through In Vitro Colchicine Treatments of Seed-Derived Explants. Int. J. Fruit Sci. 2020, 20, 1929–1941. [Google Scholar] [CrossRef]
- Sarı, N. Karpuzlarda Işınlanmış Polen Uyartımıyla Haploid Bitki Eldesi Üzerine Genotipin ve Mevsimin Etkisi Ile Işınlama Yerine Geçebilecek Uygulamalar Üzerinde Araştırmalar. Ph.D. Thesis, Cukurova University, Adana, Türkiye, 1994. (In Turkish). [Google Scholar]
- Şimşek, İ.; Göçmen, M.; Sarı, N. Diploid ve tetraploid karpuz bitkilerinde morfolojik ve sitolojik farklılıkların belirlenmesi. Derim 2013, 30, 1–14. (In Turkish) [Google Scholar]
- Norton, J.D. Testing of Plum Pollen Viability with Tetrazolium Salts. Proc. J. Am. Soc. Hortic. 1996, 89, 132–134. [Google Scholar]
- Karabıyık, Ş.; Sarıdaş, M.A.; Eti, S.; Paydaş Kargı, S. The Effects of Boron and Calcium Applications on Pollen Characteristics and Distorted Fruit Formation in Sweet ann Strawberry Varieties. Bahçe 2017, 46, 271–279. [Google Scholar]
- Adıgüzel, P.; Solmaz, İ.; Karabıyık, Ş.; Sarı, N. Comparison on Flower, Fruit and Seed Characteristics of Tetraploid and Diploid Watermelons (Citrullus lanatus Thunb. Matsum. and Nakai). Int. J. Agric. Environ. Food Sci. 2022, 6, 704–710. [Google Scholar] [CrossRef]
- Yıldırım, K.C.; Demir, İ. Changing in Pollen Viability with Storage Time, Temperature and Plant Situation. In Proceedings of the Turkey 5th Seed Congress with International Participation and Sectoral Business Forum, Diyarbakir, Türkiye, 19–23 October 2014. [Google Scholar]
- Gyulai, G.; Gémesné, J.A.; Sági, Z.; Venczel, G.; Pintér, P.; Kristóf, Z.; Törjék, O.; Heszky, L.; Bottka, S.; Kiss, J.; et al. Doubled haploid development and PCRanalysis of F1 hybrid derived DH-R2 paprika (Capsicum annuum L.) lines. J. Plant Physiol. 2000, 156, 168–174. [Google Scholar] [CrossRef]
- Ari, E.; Bedir, H.; Yildirim, S.; Yıldırım, T. Androgenic Responses of 64 Ornamental Pepper (Capsicum annuum L.) Genotypes to Shed-Microspore Culture in Autumn Season. Turk. J. Biol. 2016, 40, 706–717. [Google Scholar] [CrossRef]
- Bat, H.; Altındağ, F.N.; Yiğit, M.A.; Ellialtıoğlu, Ş.Ş.; Çölekçioğlu, N. Ploidy estimation in pepper and eggplant via stomata characteristics. Int. J. Agric. For. Life Sci. 2021, 5, 139–146. [Google Scholar]
- Eş, E. Stimulation of Autopolyploid in Phaseolus vulgaris L. Master’s Thesis, Namık Kemal University, Tekirdağ, Türkiye, 2011. (In Turkish). [Google Scholar]
- Usman, M.; Fatima, B.; Gillani, K.A.; Khan, M.S.; Khan, M.M. Exploitation of otential target tissues to develop polyploids in citrus. Pak. J. Bot. 2008, 40, 1755–1766. [Google Scholar]
- Sukamto, L.A.; Ahmad, F.; Hawo, A.H. Pengaruh Oryzalin Terhadap Tingkat Ploidi Tanaman Garut (Maratana arundinacea L.). Bul. Littro 2010, 21, 93–102. [Google Scholar]
- Moghbel, N.; Borujeni, M.K.; Bernard, F. Colchicine effect on the DNA content and stomata size of Glycyrrhiza glabra var. glandulifera and Carthamus tinctorius L. cultured in vitro. J. Genet. Eng. Biotechnol. 2015, 13, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Khandaker, M.M.; Fazil, R.; Alam, M.A.; Saifuddin, M.; Zakaria, A.J. Effects of temperature treatment on seed germination, root development and seedling growth of Citrullus lanatus (watermelon). Bulg. J. Agric. Sci. 2020, 26, 558–566. [Google Scholar]
- Jibril, S.M.; Bello, H.J. Leaf epidermal structures and stomata ontogeny in some members of the family Cucurbitaceae. Int. J. Plant Soil. Sci. 2016, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Çelikli, F.B. Effect of Different Nutrient Media and Irradiation Doses on Haploid Embriyo and Plant Formation in Galia and Kırkagac Melon (Cucumis melo L.) Genotypes. Master’s Thesis, Akdeniz University, Antalya, Türkiye, 2016. (In Turkish). [Google Scholar]
- Şensoy, A.S.; Ercan, N.; Ayar, F.; Temirkaynak, M. The Evaluation of Pollen Viability and the Determine Some Pollen Characteristics in Some Species of Cucurbitaceae Family. Akdeniz Univ. J. Fac. Agric. 2003, 16, 1–6. (In Turkish) [Google Scholar]
- Koyuncu, F. Response of In Vitro Pollen Germination and Pollen Tube Growth of Strawberry Cultivars to Temperature. Eur. J. Hortic. Sci. 2006, 71, 125–128. [Google Scholar]
- Voyiatzsis, D.G.; Paraskevopoulou-Paroussi, G. Factors Affecting the Quality and in Vitro Germination Capacity of Strawberry Pollen. Int. J. Fruit Sci. 2005, 5, 25–35. [Google Scholar] [CrossRef]
- Sánchez, M.; Velásquez, Y.; González, M.; Cuevas, J. Pollination Effectiveness of the Hoverfly Eristalinus aeneus (Scopoli, 1763) in Diploid and Triploid Associated Watermelon Crop. Insects 2022, 13, 1021. [Google Scholar] [CrossRef]
- Sawe, T.; Eldegard, K.; Totland, Ø.; Macrice, S.; Nielsen, A. Enhancing pollination is more effective than increased conventional agriculture inputs for improving watermelon yields. Ecol. Evol. 2020, 10, 5343–5353. [Google Scholar] [CrossRef]
- Sravani, V.; Ashok, P.; Sasikala, K.; Babu, B.R. Effect of Intergeneric Pollen on Induction of Parthenocarpy in Watermelon (Citrullus lanatus Thunb.). Int. J. Curr. Microbiol. App. Sci. 2018, 7, 890–895. [Google Scholar] [CrossRef]
- Şimşek, İ. Obtaining Tetraploid Lines That Is Aimed for Developing Seedless Watermelon (Citrullus lanatus (Thunb.) Matsum and Nakai) Varieties. Master’s Thesis, Cukurova University, Adana, Türkiye, 2011. (In Turkish). [Google Scholar]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).