Effect of Partial Excision of Early Taproots on Growth and Components of Hydroponic Carrots


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
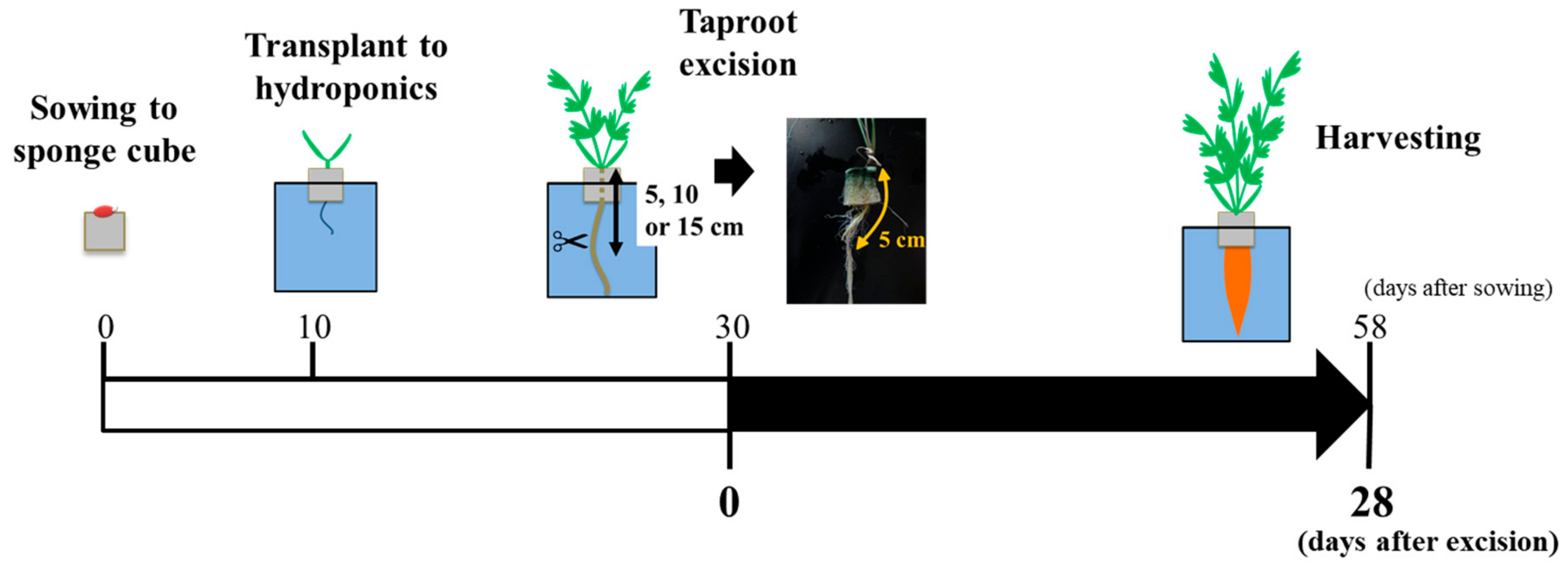
2.1. Plant Growth and Experimental Conditions
2.2. Measurement of Total Phenol Concentration
2.3. Measurement of Soluble Sugar Concentration
2.4. Data Analysis
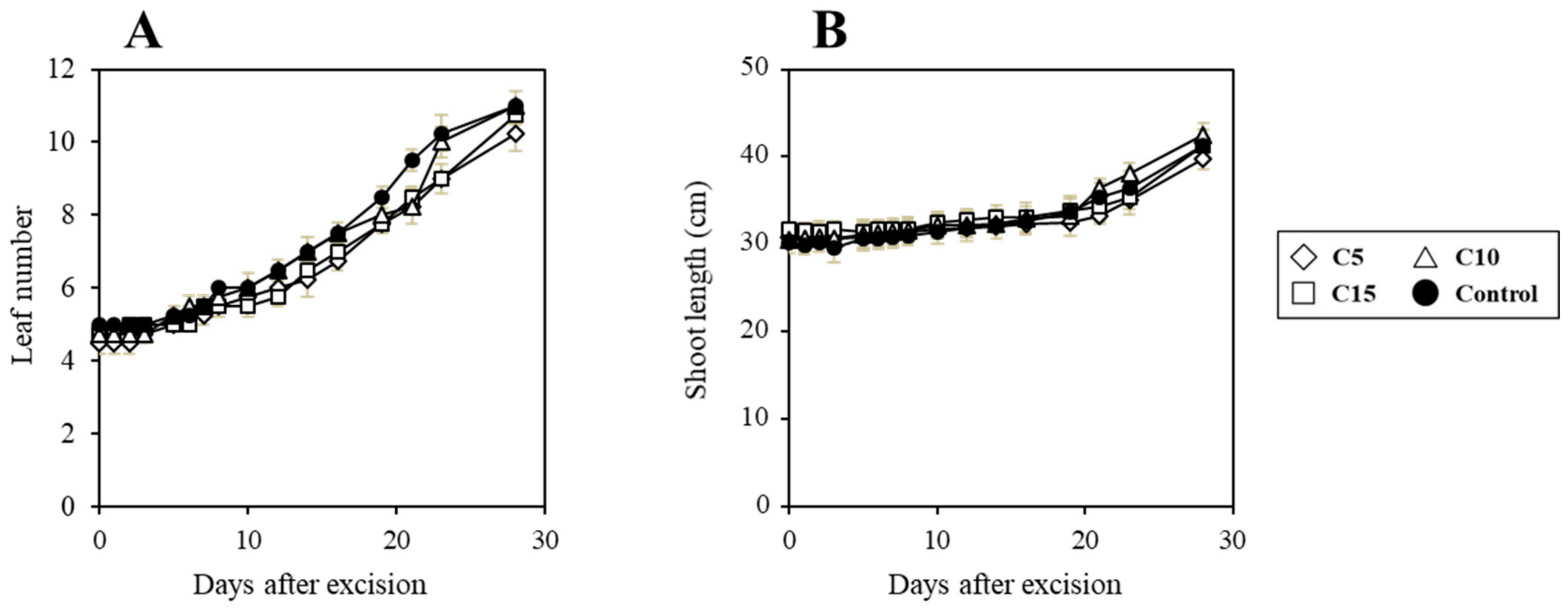
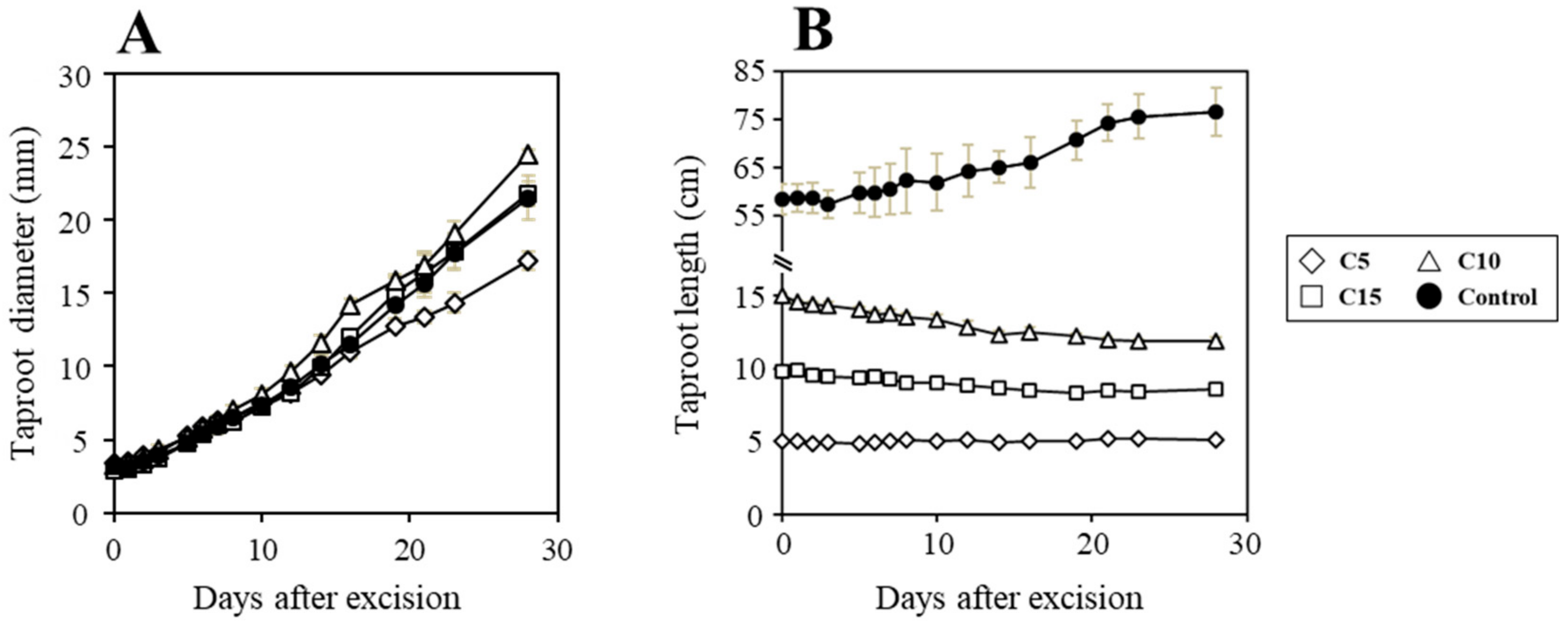
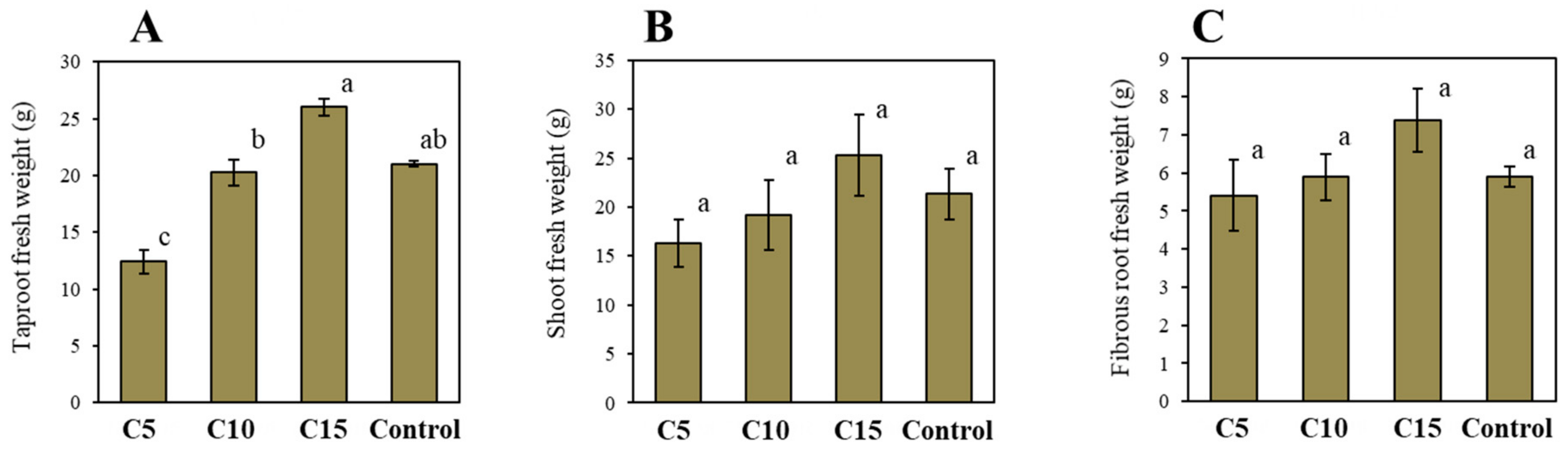
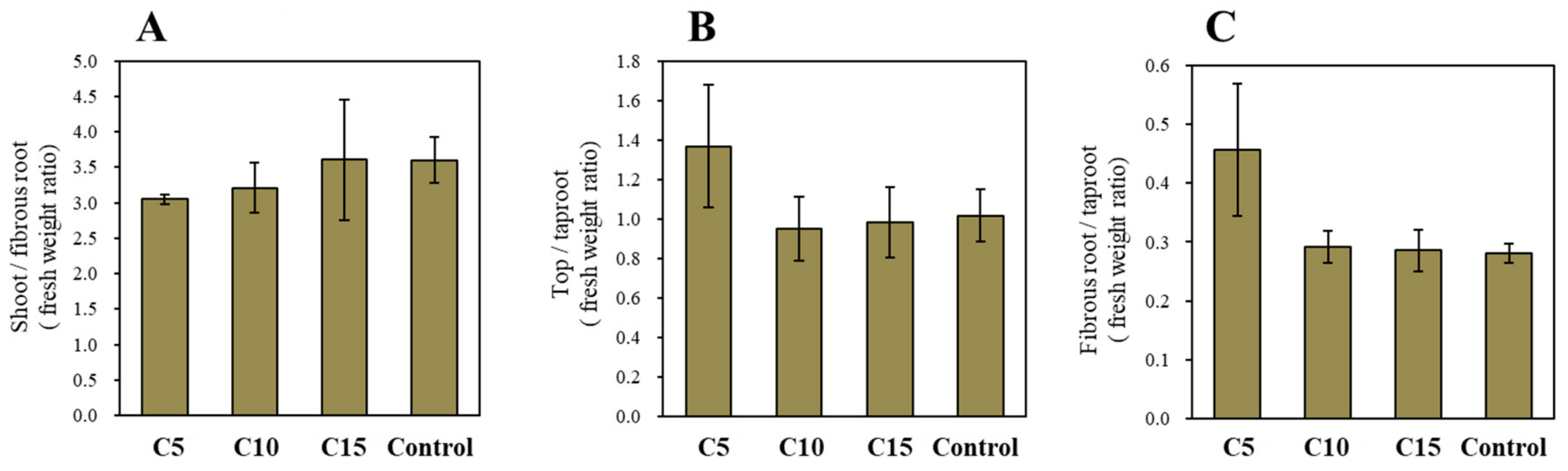
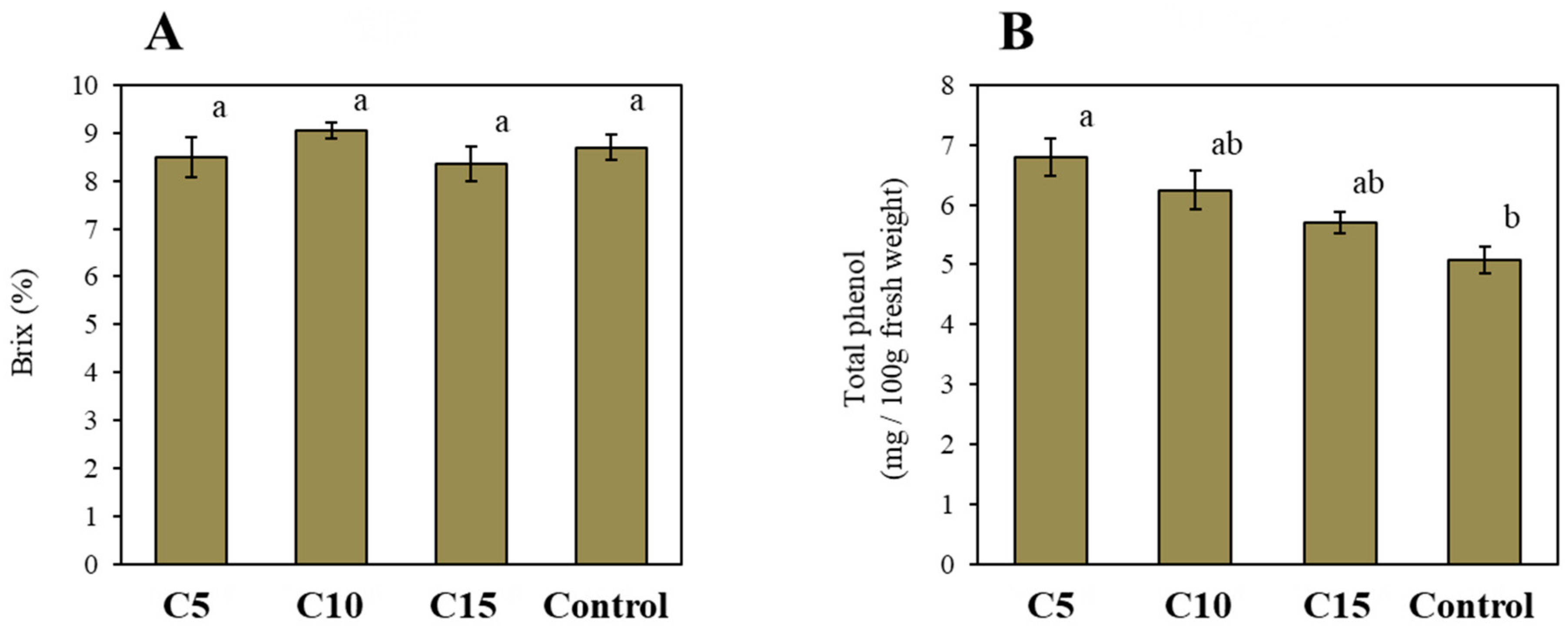
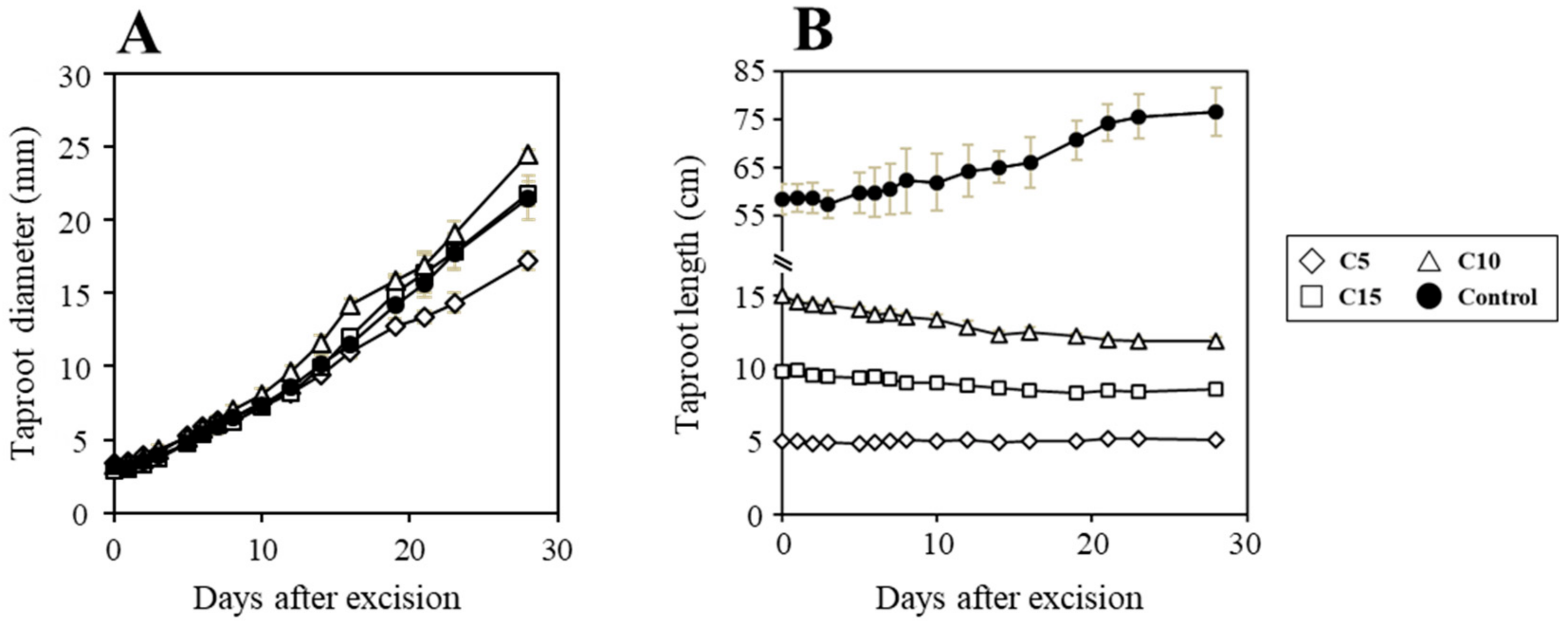
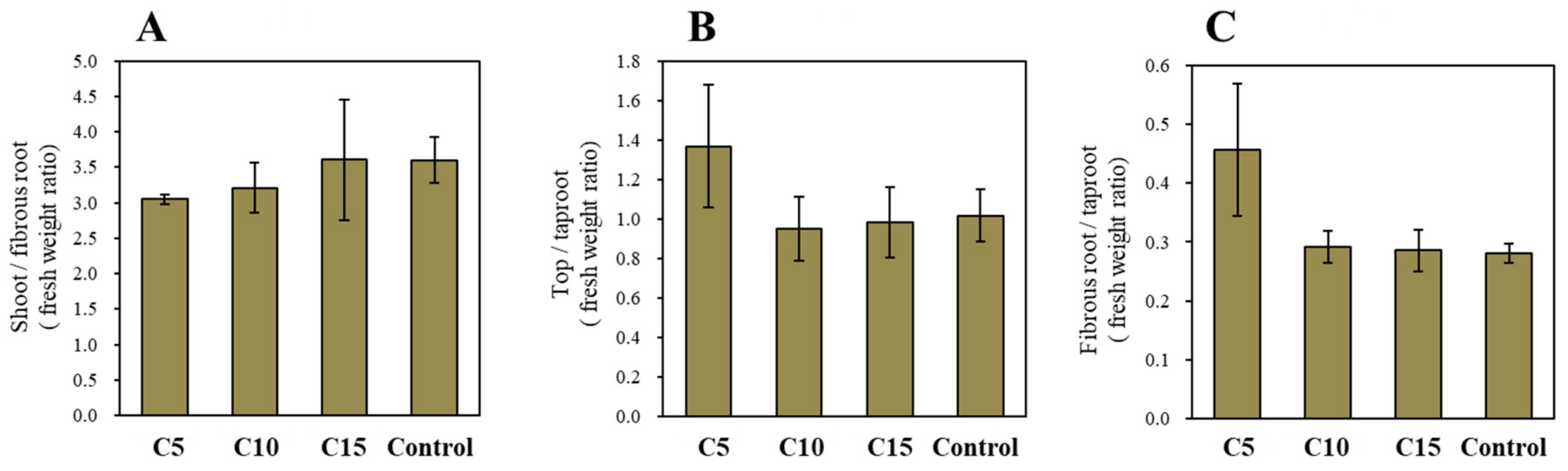
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kozai, T. Plant Factory with Artificial Light; Ohmsha Ltd.: Tokyo, Japan, 2012. [Google Scholar]
- Eguchi, T.; Ito, Y.; Yoshida, S. Periodical wetting increases α-tocopherol content in the tuberous roots of sweetpotato (Ipomoea batatas (L.) Lam.). Environ. Control Biol. 2012, 50, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Kitaya, Y.; Hirai, H.; Endo, R.; Shibuya, T. Effects of water contents and CO2 concentrations in soil on growth of sweet potato. Field Crop. Res. 2013, 152, 36–43. [Google Scholar]
- Kitaya, Y.; Hirai, H.; Wei, X.; Islam, A.F.M.S.; Yamamoto, M. Growth of sweetpotato cultured in the newly designed hydroponic system for space farming. Adv. Space Res. 2008, 41, 730–735. [Google Scholar] [CrossRef]
- Sakamoto, M.; Suzuki, T. Effect of pot volume on the growth of sweetpotato cultivated in the new hydroponic system. Sustain. Agric. Res. 2018, 7, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Chang, D.C.; Park, C.S.; Kim, S.Y.; Lee, Y.B. Growth and tuberization of hydroponically grown potatoes. Potato Res. 2012, 55, 69–81. [Google Scholar] [CrossRef]
- Koca Bozalan, N.; Karadeniz, F. Carotenoid profile, total phenolic content, and antioxidant activity of carrots. Int. J. Food Prop. 2011, 14, 1060–1068. [Google Scholar] [CrossRef]
- Terabayashi, S.; Yomo, T.; Namiki, T. Root development of root crops grown in deep flow and Ebb & flood culture. Environ. Control Biol. 1997, 35, 99–105. [Google Scholar]
- Terabayashi, S.; Harada, N.; Fujime, Y. Effects of aeration and root immersion level on the development of carrot [Daucus carota] root in hydroponics. Hortic. Res. 2008, 7, 439–444. [Google Scholar] [CrossRef]
- Islam, A.F.M.S.; Hirai, H.; Kitaya, Y. Hydroponic cultivation of carrots using modified rockwool blocks. J. Appl. Hortic. 2008, 10, 132–136. [Google Scholar]
- Gichuhi, P.N.; Mortley, D.; Bromfield, E.; Bovell-Benjamin, A.C. Nutritional, physical, and sensory evaluation of hydroponic carrots (Daucus carota L.) from different nutrient delivery systems. J. Food Sci. 2009, 74, S403–S412. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; Kobayashi, Y.; Mondal, M.F.; Ban, T.; Matsubara, H.; Adachi, F.; Asao, T. Growing carrots hydroponically using perlite substrates. Sci. Hortic. 2013, 159, 113–121. [Google Scholar] [CrossRef]
- Sakamoto, M.; Suzuki, T. Elevated root-zone temperature modulates growth and quality of hydroponically grown carrots. Agric. Sci. 2015, 6, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.; Cha, M.; Ku, Y.; Kim, H.; Bae, J. Effect of different culture nutrient solution EC on carrot top growth and nutritional contents in a closed-type plant factory system. Hortic. Sci. Technol. 2018, 36, 37–45. [Google Scholar]
- Que, F.; Wang, G.L.; Feng, K.; Xu, Z.S.; Wang, F.; Xiong, A.S. Hypoxia enhances lignification and affects the anatomical structure in hydroponic cultivation of carrot taproot. Plant Cell Rep. 2018, 37, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Suzuki, T. Synergistic effects of a night temperature shift and methyl jasmonate on the production of anthocyanin in red leaf lettuce. Am. J. Plant Sci. 2017, 8, 1534–1549. [Google Scholar] [CrossRef] [Green Version]
- Zamski, E.; Ucko, O.; Koller, D. The mechanism of root contraction in Gymnarrhena micranatha, a desert plant. New Phytol. 1983, 95, 29–35. [Google Scholar] [CrossRef]
- Smith-Huerta, N.L.; Jernstedt, J.A. Root contraction in hyacinth IV. Orientation of cellulose microfibrils in radial longitudinal and transverse cell walls. Protoplasma 1990, 154, 161–171. [Google Scholar] [CrossRef]
- Magendans, J.F.C. Elongation and contraction of the plant axis and development of spongy tissues in the radish tuber (Raphanus sativus L. cv. Saxa Nova). In Wageningen Agricultural University Papers; Pudoc Scientific Publishers: Wageningen, The Netherlands, 1991; Volume 91. [Google Scholar]
- Pütz, N.; Froebe, H.A. A re-evaluation of the mechanism of root contraction in monocotyledons using the example of Arisarum vulgare Targ.-Tozz.(Araceae). Flora 1995, 190, 285–297. [Google Scholar]
- Cresswell, A.; Sackville Hamilton, N.R.; Thomas, H.; Charnock, R.B.; Cookson, A.R.; Thomas, B.J. Evidence for root contraction in white clover (Trifolium repens L.). Ann. Bot. 1999, 84, 359–369. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Lee, B.D.; Lee, J.M. Effect of soil condition on the root contraction of ginseng. Acta Hortic. 2005, 676, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Jaffe, M.J.; Leopold, A.C. Light activation of contractile roots of Easter Lily. J. Am. Soc. Hortic. Sci. 2007, 132, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, N.; Gierlinger, N.; Pütz, N.; Fratzl, P.; Neinhuis, C.; Burgert, I. G-fibres in storage roots of Trifolium pratense (Fabaceae): Tensile stress generators for contraction. Plant J. 2010, 61, 854–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hole, C.C.; Thomas, T.H.; McKee, J.M.T. Sink development and dry matter distribution in storage root crops. Plant Growth Regul. 1984, 2, 347–358. [Google Scholar] [CrossRef]
- Marcelis, L.F.M. Sink strength as a determinant of dry matter partitioning in the whole plant. J. Exp. Bot. 1996, 47, 1281–1291. [Google Scholar] [CrossRef] [PubMed]
- Marcelis, L.F.M. Fruit growth and biomass allocation to the fruits in cucumber. 1. Effect of fruit load and temperature. Sci. Hortic. 1993, 54, 107–121. [Google Scholar] [CrossRef]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef]
- Jang, G.; Lee, J.H.; Rastogi, K.; Park, S.; Oh, S.H.; Lee, J.Y. Cytokinin-dependent secondary growth determines root biomass in radish (Raphanus sativus L.). J. Exp. Bot. 2015, 66, 4607–4619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M. Recent progress in molecular studies on storage root formation in Sweetpotato (Ipomoea batatas). Jpn. Agric. Res. Q. JARQ 2016, 50, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Jabir, O.; Mohammed, B.; Benard Kinuthia, K.; Almahadi Faroug, M.; Nureldin Awad, F.; Muleke, E.M.; Ahmadzai, Z.; Liu, L. Effects of gibberellin and gibberellin biosynthesis inhibitor (paclobutrazol) applications on radish (Raphanus sativus) taproot expansion and the presence of authentic hormones. Int. J. Agric. Biol. 2017, 19, 779–786. [Google Scholar] [CrossRef]
- Wang, C.C.; Wang, X.Y.; Wang, K.X.; Hu, J.J.; Tang, M.X.; He, W.; Vander Zaag, P. Manipulating aeroponically grown potatoes with gibberellins and calcium nitrate. Am. J. Potato Res. 2018, 95, 351–361. [Google Scholar] [CrossRef]
- Wang, G.L.; Que, F.; Xu, Z.S.; Wang, F.; Xiong, A.S. Exogenous gibberellin altered morphology, anatomic and transcriptional regulatory networks of hormones in carrot root and shoot. BMC Plant Biol. 2015, 15, 290. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.L.; Que, F.; Xu, Z.S.; Wang, F.; Xiong, A.S. Exogenous gibberellin enhances secondary xylem development and lignification in carrot taproot. Protoplasma 2017, 254, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Xu, L.; Wang, Y.; Tang, M.; Zhu, X.; Zhang, W.; Sun, X.; Nie, S.; Muleke, E.; Liu, L. Identification of critical genes associated with lignin biosynthesis in radish (Raphanus sativus L.) by de novo transcriptome sequencing. Mol. Genet. Genom. 2017, 292, 1151–1163. [Google Scholar] [CrossRef] [PubMed]
- Firon, N.; LaBonte, D.; Villordon, A.; Kfir, Y.; Solis, J.; Lapis, E.; Perlman, T.S.; Hetzroni, A.; Althan, L.; Nadir, L.A.; et al. Transcriptional profiling of sweetpotato (Ipomoea batatas) roots indicates down-regulation of lignin biosynthesis and up-regulation of starch biosynthesis at an early stage of storage root formation. BMC Genom. 2013, 14, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozai, T. Resource use efficiency of closed plant production system with artificial light: Concept, estimation and application to plant factory. Proc. Jpn. Acad. Ser. B 2013, 89, 447–461. [Google Scholar] [CrossRef] [Green Version]
- Barba-Espín, G.; Glied, S.; Crocoll, C.; Dzhanfezova, T.; Joernsgaard, B.; Okkels, F.; Lütken, H.; Müller, R. Foliar-applied ethephon enhances the content of anthocyanin of black carrot roots (Daucus carota ssp. sativus var. atrorubens Alef.). BMC Plant Biol. 2017, 17, 70. [Google Scholar]
- Müller, R.; Acosta-Motos, J.R.; Großkinsky, D.K.; Hernández, J.A.; Lütken, H.; Barba-Espin, G. UV-B Exposure of black carrot (Daucus carota ssp. sativus var. atrorubens) plants promotes growth, accumulation of anthocyanin, and phenolic compounds. Agronomy 2019, 9, 323. [Google Scholar]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakamoto, M.; Wada, M.; Suzuki, T. Effect of Partial Excision of Early Taproots on Growth and Components of Hydroponic Carrots. Horticulturae 2020, 6, 5. https://doi.org/10.3390/horticulturae6010005
Sakamoto M, Wada M, Suzuki T. Effect of Partial Excision of Early Taproots on Growth and Components of Hydroponic Carrots. Horticulturae. 2020; 6(1):5. https://doi.org/10.3390/horticulturae6010005
Chicago/Turabian StyleSakamoto, Masaru, Masanori Wada, and Takahiro Suzuki. 2020. "Effect of Partial Excision of Early Taproots on Growth and Components of Hydroponic Carrots" Horticulturae 6, no. 1: 5. https://doi.org/10.3390/horticulturae6010005
APA StyleSakamoto, M., Wada, M., & Suzuki, T. (2020). Effect of Partial Excision of Early Taproots on Growth and Components of Hydroponic Carrots. Horticulturae, 6(1), 5. https://doi.org/10.3390/horticulturae6010005