3.1. Diversity in Ecophysiological Parameters in Sweet Potato Ecotypes Evaluated under Low-Altitude Conditions
Sweet potato ecotypes collected from several altitude conditions showed a wide response in the tolerance to low altitude conditions. Genotype and altitude factors analyzed independently showed significant differences in some physiological parameters, the three intervals of altitude showed significant differences in Pn, E, VPD, WUE and CEI, while the genotype factor showed statistical differences for all ecophysiological traits (
Table 3).
Through the categorization by altitudinal origin, the three groups presented a differential behavior. Genotypes from high altitude (1401–2555 masl) exhibited the lowest Pn, WUE and CEI, but the highest E and VPD compared with others altitude ranges. These values for physiological parameters, were possibly related to a minor tolerance to low altitude conditions (
Table 4). Genotypes from the intermediate altitude range (924–1298 masl), exhibited intermediate values for Pn, WUE, CEI, E and VPD. Finally, the genotypes from the low altitude range (18–599 masl), showed the highest values for Pn, WUE and CEI, but conversely, they had the lowest values for E and VPD compared with the other altitude intervals. These results indicated that genotypes from this group presented a better physiological response under the low altitude conditions than the genotypes from the greater altitudes, possibly because they were from warm climate regions and had evolved mechanisms to tolerate low altitude conditions (104 masl). The broad adaptation of this species to different edaphoclimatic conditions [
13], and the results obtained in this study, suggested that the species has wide variability to tolerate low altitude conditions and the physiological adaptation of low altitude genotypes indicated higher trait value in comparison with genotypes from other altitudinal ranges.
Regarding the ecophysiological diversity of the characterized genotypes, net photosynthetic rate (Pn) had a range between 21 and 33 µmol CO
2 m
−2 s
−1. The genotypes 0715-076.CES (599 masl), 0516-357.QND (1298 masl) and Agrosavia Ambarina (18 masl) presented the highest values, with 33.04, 32.47 and 31.09 µmol CO
2 m
−2 s
−1, respectively (
Table 4), while 0816-615.STR (1426 masl) and 0516-503.CLD (1749 masl), genotypes from high altitude, presented the lowest averages (21 µmol CO
2 m
−2 s
−1). Most of the values among genotypes were within the range reported for C3 plants [
28,
29]; however, the wide range in this study associated with differences between sweet potato ecotypes used in the study. Previous studies observed Pn values below 20 µmol CO
2 m
−2 s
−1, in sweet potato genotypes subjected to water stress [
9,
11,
30]. The values found in our study showed that the evaluation conditions guaranteed anoptimal physiological state of the evaluated ecotypes. High values of Pn in genotypes from low and intermediate altitudes may indicate a high potential of tolerance to low altitude conditions and possibly to low water availability, since these conditions cause stomatal closure, reduction of Pn and affect the translocation of assimilates to roots. The growth habit of the plant in certain genotypes alloweds it to maintain humidity in the soil, a morphological adaptation that, in turn, would allow it to improve the water status of the plant, modifying the opening and closing of stomata, transpiration and assimilation [
31,
32].
Stomatal conductance (Gs) is a parameter that relates the number and degree of stomatal opening [
33]. A range of variation for Gs was between 0.88–1.95 mol H
2O m
−2 s
−1. The genotypes 0316-256.STR (924 masl), 0816-620.STR (521 masl) and 0816-587A.BOY (2060 masl) presented an average higher than 1.90 mol H
2O m
−2 s
−1, while the genotypes 0816-615.STR (1426 masl) and 0516-503.CLD (1749 masl) showed an average lower than 0.92 mol H
2O m
−2 s
−1. The values found were similar to those exhibited in genotypes subjected to a level of humidity close to the field capacity, since less soil humidity induces the closure of stomata and the values found under these conditions are below 0.8 mol H
2O m
−2 s
−1 [
34,
35]. However, certain sweet potato genotypes are capable of controlling the osmotic potential in the leaves through different mechanisms that regulate stomatal opening through an increase in proline levels, thus, avoiding a decrease in Pn and Gs [
9,
36] and unnecessary water loss through transpiration. Although a broad response was found in the stomatal conductance of the evaluated ecotypes, their adequate tolerance was evidenced regardless of their altitudinal origin.
The concentration of intercellular CO
2 fixed by plants (ICO
2) influences photosynthesis. The genotypes evaluated presented values between 271.01–312.32 μmol CO
2 mol
−1, some being congruent with previous studies [
37], which described values between 231 and 296 μmol CO
2 mol
−1. The genotypes that exhibited the highest ICO
2 values may indicate a greater capacity for CO
2 entry, due to a greater stomatal conductance. However, this result did not generate any increase in Pn, probably because C3 species can have a saturation effect on Rubisco, decreasing its activation state and equilibrium [
10,
36,
37].
Transpiration (E) corresponds to the flow of water, in the form of vapor, generated between the plant and the atmosphere [
38,
39]. There were wide differences in E among the genotypes, those from a high altitude 0816-581.BOY (1689 masl), 0316-263.STR (1650 masl) and 0516-491.CLD (1899 masl), presented the highest values (14.9, 14.8 and 14.1 mmol H
2O m
−2 s
−1, respectively); the lowest values were around 8.74–10.52 mmol H
2O m
−2 s
−1, in genotypes from intermediate and low altitude Agrosavia Ambarina (18 masl), 0113-660.VAL (1000masl) and 0113-668.VAL (1000 masl). Previous studies reported values between 4 and 17 mmol H
2O m
−2 s
−1 in two varieties of sweet potato subjected to different regimes of soil moisture, the highest values with a higher soil moisture content [
34]. Different humidity regimes may influence this trait [
40]. The wide diversity found among evaluated genotypes was consistent with the differing responses that can be found between different genotypes [
1]. However, the differential response of genotypes from high altitude showing higher values compared with the genotypes from other altitudinal intervals suggested their lower adaptation to low altitude conditions, since the adapted genotypes exhibited lower values.
Vapor pressure deficit (VPD) is high when plants close stomata to avoid excessive water loss [
41], sometimes negatively affecting photosynthesis and yield, due to the increase in stomatal resistance. 0816-615.STR (1426 masl) and 0516-503.CLD (1749 masl), genotypes from a high altitude, presented the highest VPD values (1.96 and 1.92 kPa, respectively), and were the genotypes with the lowest Pn. Moreover, the genotype that presented the highest Pn (0715-076.CES, 599 masl) had an intermediate VPD value (1.35 kPa), indicating adequate adaptation of genotypes from low altitude to low altitude evaluation conditions and an ability to keep the plant water content in balance, with an intermediate conductance and transpiration.
Variable responses for leaf internal temperature (LT) was found among the genotypes, with values between 30.54–35.27 °C. Genotype 0715-076.CES (599 masl) presented the highest LT (35.27 °C), and simultaneously exhibited the highest Pn (33.04 µmol CO
2 m
−2 s
−1). Although it has been reported that temperatures around 35 °C influence the reduction of the enzymatic capacity of Rubisco [
42], the results in this investigation indicated that there was no negative effect on Pn with the increase in LT among the genotypes. This process is favorable because the plant will not invert the water reserves, under conditions of water deficit, to reduce the leaf temperature [
43]. This response differs from that reported in another C3 species (cassava
Manihot esculenta), in which an increase in leaf temperature reduced photosynthesis [
43].
Water use efficiency (WUE) represents a characteristic of great interest in plants, allowing the use of this resource in a regulated way under conditions of soil water deficit [
44]. The genotypes with the highest WUE exceeded the value of 1.5 µmol CO
2/mmol H
2O, which corresponded to the highest value found in sweet potato genotypes in previous reports [
45], indicating more efficient water use in genotypes from intermediate and low altitudes. The genotypes with the highest WUE were Agrosavia Ambarina (18 masl), 0113-668.VAL (1000 masl) and 0113-660.VAL (1000 masl). WUE was a variable that was related to E, and among the genotypes with the best response for WUE were the same genotypes with low E values, which showed the ability of these genotypes to fix the necessary CO
2, using the available water efficiently and decreasing transpiration process.
The carboxylation efficiency index (CEI) was estimated using the relationship between Pn and ICO
2. The genotypes from intermediate and low altitudes 0715-076.CES (599 masl) and 0516-357.QND (1298 masl) were the most efficient according to the CEI, with values of 0.1183 and 0.1175, respectively. The least efficient genotype was a genotype from high altitude, 0516-503.CLD (1749 masl) with 0.0737. Efficiency in the carboxylation process may be associated with the ability of Rubisco to increase its affinity for CO
2 fixation with respect to intracellular O
2 and, thus, reduce photorespiration rates and increase Pn [
25,
46]. Consistently, the genotypes that presented the highest Pn reached the highest values for CEI also. This ecophysiological characterization revealed a wide variability between the sweet potato genotypes from different altitude (18 to 2555 masl). In general, the genotypes from low and intermediate altitudes showed a better physiological response, which was confirmed by their high Pn and CEI and low VPD, while only some genotypes from higher altitudes showed such responses.
3.2. Relationship among Ecophysiological Parameters in Sweet Potato
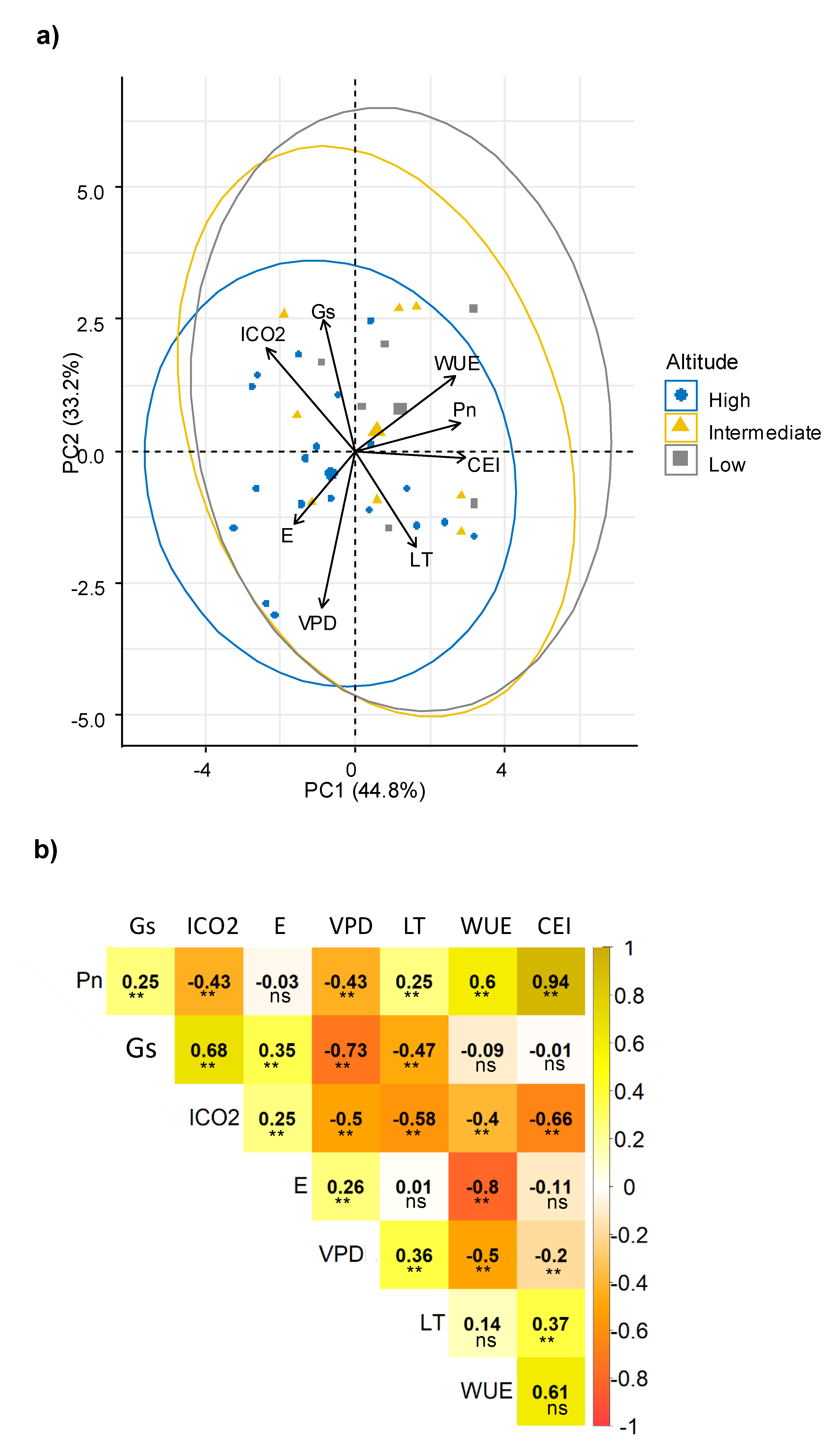
Multivariate analysis determined relationships among physiological traits that although highly variable among the genotypes, indicated two principal components (PC) explained 78% of all variation. Consistently, a relationship among WUE, Pn and CEI was observed (
Figure 1a); all of these parameters were related to the efficiency of the photosynthetic process, and including a third PC (PC3 explained 15.17%), the component matrix indicated that CEI, Pn and WUE were the most important traits for PC1 (
Supplementary materials Table S1). Another potential relationship was observed between Gs and ICO
2, and between E and VPD.
The interrelations among physiological traits were confirmed by correlation analysis (
Figure 1b). Consistently, Pn had a direct correlation with WUE. WUE is considered a key mechanism that allows a reduction in water loss and increase in CO
2 assimilation. Therefore, it is a desirable trait that indicates good adaptation and greater plasticity of the genotypes [
47,
48]. The positive significant correlation between Gs with ICO
2 confirmed the association previously described. A high content of ICO
2 is a result of an increase in Gs, and, consistently with its negative correlation, can simultaneously reduce VPD [
41], Pn, WUE and CEI [
47,
49]. A greater Gs allows an increase in the flux of CO
2 into the plant and can affect transpiration rates and subsequently, the Pn process, as previously reported [
10,
47,
49,
50,
51]. However, a significant reduction of ICO
2 can also reduce Pn [
47].
Regarding the relationship between E and VPD, the positive correlation indicated that there was an increase of E, until a maximum limit of VPD (values below 2 kpa); then, E was reduced. In fact, when the VPD increases, stomata are forced to close to protect the plant from dehydration [
41]. In contrast, an inverse relation between VPD and Gs was observed; an expected response due to VPD increase, forcing the stomata to close to protect the plant from dehydration [
41]. Additionally, an inverse relationship between WUE and both VPD and E was found, indicating that sweet potato plants were more efficient in the use of this resource when the Gs, E and VPD decreased due to greater stomatal resistance and lower water loss [
41]. In addition, this correlation was also reported previously in related species of
Ipomoea, in which a low water loss was found, caused by low Gs, along with the maintenance of good photosynthetic behavior, which represents an adequate WUE [
47].
The positive correlation between Gs and E can be explained by a greater opening of the stomatacausing E to continue along with Pn [
52,
53]. In respect to LT, this parameter decreased with the increase of Gs because the transpiration process is favored in the plant [
44,
54,
55], and water release promote a cooling effect in the leaf. An inverse correlation was found for ICO
2 and LT, indicating that as the LT increased, the ICO
2 was reduced; this may be due to the stomatal closure effect generated by heating the plant canopy, to reduce the E [
56], which would have caused a reduction in CO
2 availability. Similarly, the negative correlation between Pn and VPD indicated a reduction of Pn when the VPD values were high. This behavior is conditioned by the plant response, throughstomatal closure mechanism, which reduces dehydration [
9,
41,
51,
57]. In this way, it is evident that when the Gs was reduced, the diffusion of CO
2 was affected, also implying an effect on Pn [
25].
These associations between ecophysiological parameters demonstrated tolerance strategies in a wide variability of the sweet potato genotypes. The response of Pn, WUE and CEI in the genotypes was largely determined by the values of Gs, ICO2 and E. These parameters defined the response of the genotypes to the evaluation conditions.
3.3. Interaction between Ecophysiological and Morpho-Agronomic Parameters Involved in Sweet Potato Tolerance to Low-Altitude Conditions
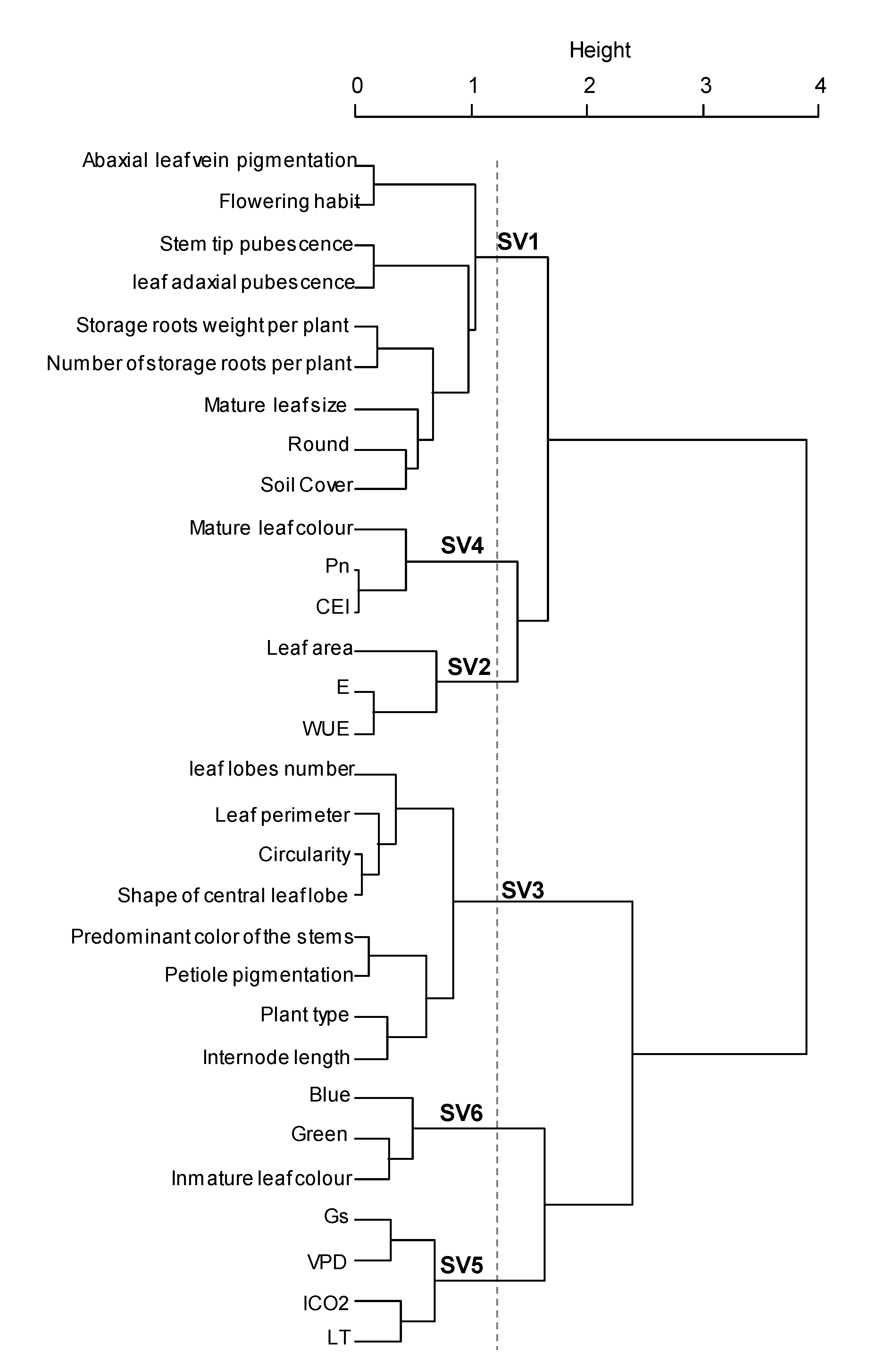
Results from morpho-agronomic and ecophysiological characterization was analyzed simultaneously to determine the interaction of these traits on the tolerance of sweet potato to low altitude conditions. Although, a wide diversity was found among genotypes, several traits were strongly related to each other, providing the same information and producing data redundancy. Therefore, to avoid this constraint, a hierarchical analysis of quantitative and qualitative variables was used for variable selection, determination of any association between them and creation of synthetic variables to improve the integral analysis of the genotypes. Thirty variables were selected, which showed higher stability of the partitions and represented several variables that were not further included, such as: secondary color of the stems, shape of the central leaf lobe, leaf solidity, red coloration, internode diameter, stem latex, petiole pubescence, mature leaf width, petiole diameter and general outline of the leaf. Six general groups were produced from the analyzed variables and were defined as synthetic variables SV1, SV2, SV3, SV4, SV5 and SV6 (
Figure 2). SV1 mainly involved parameters related to the plant type, pubescence, tuberous root formation, ground cover, leaf size and leaf roundness; SV2 grouped characteristics of WUE, E and leaf area; SV3 grouped traits related to leaf shape and pigmentation; SV4 represented Pn, CEI and mature leaf color; SV5 only represented physiological attributes such as Gs, VPD, ICO
2 and LT; finally, SV6 grouped leaf color attributes (
Figure 2,
Supplementary Materials Table S2). This data analysis was performed in line with previous studies which have shown that phenotypic descriptions may contain redundant variables and that variable aggregation can be used to select an optimal set of explanatory variables for an appropriate statistical model by removing multi-collinearity [
58], an aspect generated by the similarity among some groups of variables that bring the same information [
24].
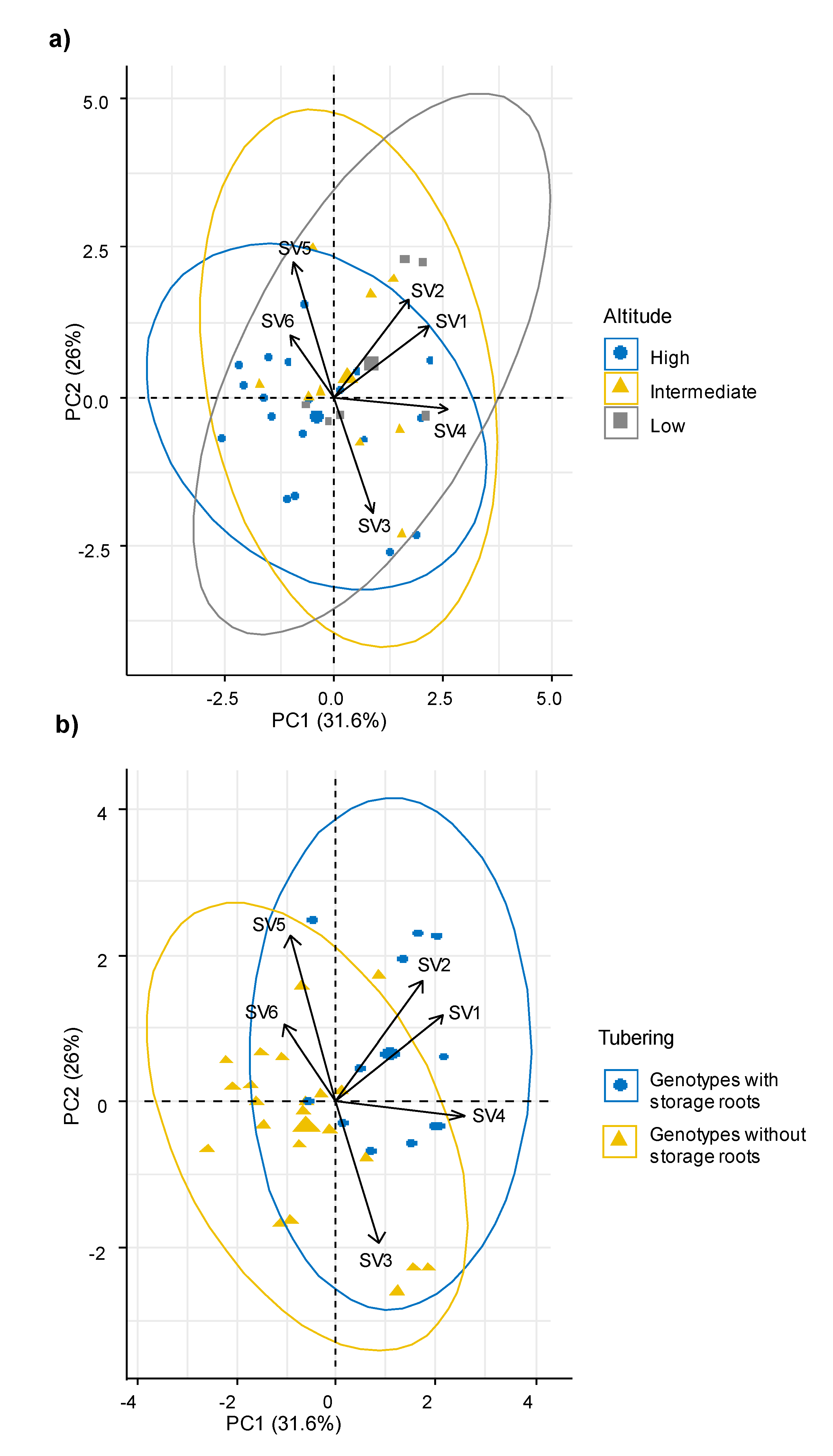
Using these synthetic variables, the behavior of the genotypes from different altitude intervals and with differential tuberization capacity was analyzed. The multivariate analysis showed that the ecophysiological response, the morphological characteristics and the ability to produce storage roots were independent of the altitudinal origin of the sweet potato genotypes (
Figure 3a). Meanwhile, genotypes with capacity to produce storage roots were associated with the synthetic variables SV2, SV4 and SV1, which grouped mainly gas exchange parameters such as Pn, WUE, E and CEI; plant type, soil cover, leaf area, leaf roundness, flowering habit and productive parameters (
Figure 3b).
The results of the correlation analysis confirmed the relationship among different phenotypic characteristics (morphology and color), physiological parameters and with storage root production previously determined by cluster analysis (
Supplementary materials Table S3). From a cluster that grouped the synthetic variables SV1, SV4 and SV2, ground cover, mature leaf size, and flowering habit, a positive correlation with Pn, CEI and WUE, was observed. However, abaxial leaf vein pigments, mature leaf color and leaf adaxial pubescence presented negative correlation with these physiological parameters. A cluster that grouped the synthetic variables SV3, SV6 and SV5, showed a positive correlation with leaf perimeter and Pn, as with circularity with Gs and ICO
2, and internode length was negatively correlated with Pn, ICO
2, E, VPD and WUE.
These results may be associated with the direct effect of leaf size, leaf maturation and amount of biomass, which correspond to parameters associated with the main photosynthetic structures, and is related to Pn, WUE and water loss by transpiration [
59,
60]. However, when leaves reach their maximum maturity and senescence begins, the photosynthetic capacity is reduced [
61]. These morphological traits also were related to LT, CEI and WUE; however, most of the traits that were positively related to Pn were negatively associated to VPD. Flowering habit depends on cultivar and growing conditions, but there was no evidence of any relationship with photosynthetic process. Negative correlation of ecophysiological parameters (Pn, ICO
2, E, VPD and WUE) with variables associated with leaf shape, presence of pigments in plant organs and plant architecture were found. Some plants have structural and pigmentation modifications in the leaves as protective barriers to environmental factors, which reduce their absorption of radiation, avoiding saturation of plant, but exposing the photosynthetic structures to an imbalance [
62]. Furthermore, this type of barrier can reduce the absorption of photosynthetic wavelengths and affect Pn by photoinhibition [
62]. On the other hand, the presence of pubescence may contribute to reduced transpiration [
63], which related to stomatal closure and may imply a negative effect on Pn [
64]. Adaptation to the conditions of genotype origin is given by the physiological characteristics that allow them to adapt to the environment in which they develop and stimulate the acquisition of water, light and other components for growth [
65].
The characterized population presented a wide phenotypic and ecophysiological variability between and within groups categorized by different altitudinal intervals (high, intermediate and low altitude), and tuberization capacity (genotypes with storage roots and genotypes without storage roots). Variance analysis integrating altitude and tuberization factors and their interaction showed that for altitude intervals, only Pn and WUE presented statistically significant differences. Regarding tuberization the parameters Pn, WUE, CEI, root number, root weight, leaf circularity, leaf roundness and leaf solidity were statistically different, while the interaction was not statistically significant for any variable (
Table 5).
The genotypes presented a wide variability in morphometric and agronomic parameters within each altitudinal interval; however, between groups no significant differences were detected (
Table 6). Most of the genotypes that tuberized were collected in places of low altitude, and only some genotypes of high altitude showed the ability to adapt to the evaluation conditions. The evaluated genotypes were grouped with respect to their capacity to produce storage roots, in order to determine the characteristics that allowed to guarantee better tolerance to low altitude conditions, and to be comparable to the Aurora and Ambarina varieties. Genotypes 0715-076.CES, 0316-251.STR, 0316-256.STR, 0316-283A.STR, 0316-299.STR, 0316-300.STR, 0516-352-HUI, 0816-600.BOY, 0816-618.STR and 0113-668.VAL produced storage roots at 120 DAP. The genotype 0316-299.STR presented the highest yield per plant, with 1.326 gr/plant, being comparable with the Ambarina variety (
Table 6).
The group that had the ability to produce storage roots presented higher Pn, WUE and CEI, compared to the group that did not have the ability to produce storage roots (
Table 7). This observation was consistent with reports in which the positive relationship between photosynthesis and root production is indicated [
4,
9]; this condition is also attributed to other C3 species such as cassava [
28], guarantying a major amount of photo-assimilates that can be translocated to reserve organs, in this case the roots [
30]. This group had a higher WUE and CEI compared to the group without storage roots, indicating that these genotypes made efficient use of the water resource [
66], an aspect also corroborated by the low E found in this group. This group also presented the lowest VPD, which could be related to its adequate Pn behavior, since it has been reported that high VPD values indicate negative effects on Pn, due to high stomatal closure [
41].
Regarding the morphometric characteristics of the leaves, the group that tuberized presented a larger leaf area, circularity, leaf roundness and leaf solidity, and lower perimeter, indicating the presence of lobed or more complex leaves [
27], compared to the group without storage roots. These results indicated that in the group with capacity to produce storage roots, most of the genotypes had leaves with a major leaf area available for the capture of sunlight, an aspect that could favor the assimilation of CO
2 by having a larger reception area for photosynthetically active radiation (PAR). Furthermore, the best photosynthetic efficiency favors the production of carbohydrates and, consequently, the production of storage roots [
4,
8,
9].
In the qualitative parameters, dependence analysis showed association of tuberization capacity with soil cover efficiency, mature leaf size and latex production in the stems (
Table 8). The genotypes that produced storage roots had a higher soil cover and mature leaf size, and lower latex production, compared to the group without storage roots. These results were consistently confirmed by quantitative variables in which it was shown that the genotypes with capacity to store starches in the roots presented a major leaf area and complete leaf lamina (less lobed). Although, the physiological and agronomic response of the genotypes was not totally related to their altitudinal origin, most of the genotypes that tuberized originated from altitudes between 18 and 1160 m above sea level, with the exception of four genotypes, whose origin was between 1481 and 1901 masl (
Figure 4). In general, tolerance to low altitude conditions, considered as the ability to survive and produce storage roots, involved a high Pn in certain genotypes that together with the controls, Aurora and Ambarina, had the ability produce storage roots. These results showed the phenotypic plasticity of the genotypes, since even some genotypes from high altitudes were able to produce storage roots in conditions contrasting to those of their origin, and others that, despite having been evaluated in comparable conditions, showed a low productive performance. The genotypes from high altitude that showed low Pn values had small leaves with major complexity (more lobed) and did not have the ability to produce storage roots.
These results suggested a wide variability in phenotypic and ecophysiological response in the sweet potato genotypes. Leaf size (large) and complexity (less lobed), together with soil cover efficiency, can influence the microclimate conditions and ensure major photosynthetic efficiency that favors the production and accumulation of carbohydrates [
4,
8,
9]. In the evaluated sweet potato genotypes, these parameters played a key role in guaranteeing their tolerance to low altitude conditions. Additionally, factorial analysis showed that some ecophysiological parameters, morphological traits and the tuberization capacity were related to the altitudinal origin of the genotypes. Consistently, discrimination of genotypes according to tuberization capacity was found through parameters grouped in the synthetic variables SV2 (WUE, E and leaf area), SV4 (Pn, CEI and Mature leaf color) and SV1 (plant type, pubescence, tuberous roots formation, ground cover, leaf size and leaf roundness, among others), suggesting the key role of these traits for tolerance to low altitude condition.



 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}